
Resuscitation of Pulsed Electric Field-Treated Staphylococcus aureus and Pseudomonas putida in a Rich Nutrient Medium



Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions
2.2. PBS Solution
2.3. Solution pH and Conductivity
2.4. Total Specific Energy
2.5. Heat Transfer Model
2.6. Design and Construction of the Electroporator
2.6.1. Construction of the Electroporator Chamber
2.6.2. Applied PEF Characterization
2.6.3. PEF Procedure for Determining the Conditions of Bacterial Eradication
2.6.4. PEF Procedure for Determining Membrane Permeability, Bacterial Size and Viability as a Function of Dilution in PBS and BHI Medium
2.7. Viable Count Assay
2.8. Examination of Bacterial Membrane Permeability and Cell Size by FCM Analysis
2.9. Proteolysis and Mass Spectrometry Analysis
2.9.1. Proteolysis
2.9.2. Mass Spectrometry Analysis
2.10. Statistics
3. Results and Discussion
3.1. Total Specific Energy and the Temperature Profile
3.2. PEF Conditions for P. putida F1 and S. aureus Eradication
3.3. Viability of PEF-Treated Bacteria as a Function of Suspension in BHI Medium and PBS
3.4. Membrane Permeability of PEF-Treated Bacteria Suspended in BHI Medium and PBS
3.5. Cell Size of PEF-Treated Bacterial as a Function of Dilution in BHI Medium and PBS
3.6. MS Analysis of the Proteins from PEF-Treated P. putida F1 Suspension, Compared to Nontreated Bacteria
4. Conclusions
Supplementary Materials
 ) Voltage output (
) Voltage output (  ), current output (
), current output (  ). Temperature in the electroporation chamber. Figure S3: Temperature profile at different times during the PEF treatment along the electrodes (A); Maximum temperature (
). Temperature in the electroporation chamber. Figure S3: Temperature profile at different times during the PEF treatment along the electrodes (A); Maximum temperature (  ) in the electrode sample system, and average temperature ( ) in the electrode sample system (B); Electrode sample system’s temperature map after 70 s operation time (C). Figure S4: Over-expressed proteins in PEF-treated P. putida F1 in BHI, compared to the non-treated sample.
) in the electrode sample system, and average temperature ( ) in the electrode sample system (B); Electrode sample system’s temperature map after 70 s operation time (C). Figure S4: Over-expressed proteins in PEF-treated P. putida F1 in BHI, compared to the non-treated sample.Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neumann, E.; Kakorin, S.; Tœnsing, K. Fundamentals of electroporative delivery of drugs and genes. Bioelectrochem. Bioenerg. 1999, 48, 3–16. [Google Scholar] [CrossRef]
- Kotnik, T.; Frey, W.; Sack, M.; Meglič, S.H.; Peterka, M.; Miklavčič, D. Electroporation-based applications in biotechnology. Trends Biotechnol. 2015, 33, 480–488. [Google Scholar] [CrossRef]
- Schottroff, F.; Krottenthaler, A.; Jaeger, H. Stress induction and response, inactivation, and recovery of vegetative microor-ganisms by pulsed electric fields. In Handbook of Electroporation; Springer International Publishing AG: Cham, Switzerland, 2017; pp. 2539–2557. [Google Scholar]
- Yarmush, M.L.; Golberg, A.; Serša, G.; Kotnik, T.; Miklavčič, D. Electroporation-Based Technologies for Medicine: Principles, Applications, and Challenges. Annu. Rev. Biomed. Eng. 2014, 16, 295–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pucihar, G.; Kotnik, T.; Kandušer, M.; Miklavčič, D. The influence of medium conductivity on electropermeabilization and survival of cells in vitro. Bioelectrochemistry 2001, 54, 107–115. [Google Scholar] [CrossRef]
- Baldwin, W.H.; Gregory, B.W.; Osgood, C.J.; Schoenbach, K.H.; Kolb, J.F. Membrane Permeability and Cell Survival after Nanosecond Pulsed-Electric-Field Exposure—Significance of Exposure-Media Composition. IEEE Trans. Plasma Sci. 2010, 38, 2948–2953. [Google Scholar] [CrossRef]
- Kramar, P.; Miklavcic, D.; Lebar, A.M. Determination of the lipid bilayer breakdown voltage by means of linear rising signal. Bioelectrochemistry 2007, 70, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Pucihar, G.; Krmelj, J.; Reberšek, M.; Napotnik, T.B.; Miklavčič, D. Equivalent Pulse Parameters for Electroporation. IEEE Trans. Biomed. Eng. 2011, 58, 3279–3288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinosita, K.; Tsong, T.Y. Voltage-induced pore formation and hemolysis of human erythrocytes. Biochim. Biophys. Acta BBA Biomembr. 1977, 471, 227–242. [Google Scholar] [CrossRef]
- Rols, M.; Teissié, J. Electropermeabilization of mammalian cells. Quantitative analysis of the phenomenon. Biophys. J. 1990, 58, 1089–1098. [Google Scholar] [CrossRef] [Green Version]
- Espina, L.; García-Gonzalo, D.; Pagán, R. Detection of Thermal Sublethal Injury in Escherichia coli via the Selective Medium Plating Technique: Mechanisms and Improvements. Front. Microbiol. 2016, 7, 1376. [Google Scholar] [CrossRef]
- Davis, C. Enumeration of probiotic strains: Review of culture-dependent and alternative techniques to quantify viable bacteria. J. Microbiol. Methods 2014, 103, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Schottroff, F.; Fröhling, A.; Zunabovic-Pichler, M.; Krottenthaler, A.; Schlüter, O.; Jäger, H. Sublethal Injury and Viable but Non-culturable (VBNC) State in Microorganisms During Preservation of Food and Biological Materials by Non-thermal Processes. Front. Microbiol. 2018, 9, 2773. [Google Scholar] [CrossRef] [PubMed]
- Colwell, R.R. Viable but not cultivable bacteria. In Uncultivated Microorganisms; Epstein, S.S., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 121–129. [Google Scholar]
- Ayrapetyan, M.; Oliver, J.D. The viable but non-culturable state and its relevance in food safety. Curr. Opin. Food Sci. 2016, 8, 127–133. [Google Scholar] [CrossRef]
- Kell, D.B.; Kaprelyants, A.S.; Weichart, D.H.; Harwood, C.R.; Barer, M.R. Viability and activity in readily culturable bacteria: A review and discussion of the practical issues. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 1998, 73, 169–187. [Google Scholar] [CrossRef] [PubMed]
- Bogosian, G.; Bourneuf, E.V. A matter of bacterial life and death. EMBO Rep. 2001, 2, 770–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Mendis, N.; Trigui, H.; Oliver, J.D.; Faucher, S.P. The importance of the viable but non-culturable state in human bacterial pathogens. Front. Microbiol. 2014, 5, 258. [Google Scholar] [CrossRef] [Green Version]
- Koukkidis, G.; Haigh, R.; Allcock, N.; Jordan, S.; Freestone, P. Salad Leaf Juices Enhance Salmonella Growth, Colonization of Fresh Produce, and Virulence. Appl. Environ. Microbiol. 2016, 83, e02416-16. [Google Scholar] [CrossRef] [Green Version]
- Ferro, S.; Amorico, T.; Deo, P. Role of food sanitising treatments in inducing the ‘viable but nonculturable’ state of microorganisms. Food Control. 2018, 91, 321–329. [Google Scholar] [CrossRef]
- Huo, Z.-Y.; Zhou, J.-F.; Wu, Y.; Wu, Y.-H.; Liu, H.; Liu, N.; Hu, H.-Y.; Xie, X. A Cu3P nanowire enabling high-efficiency, reliable, and energy-efficient low-voltage electroporation-inactivation of pathogens in water. J. Mater. Chem. A 2018, 6, 18813–18820. [Google Scholar] [CrossRef]
- Jaeger, H.; Schulz, M.; Lu, P.; Knorr, D. Adjustment of milling, mash electroporation and pressing for the development of a PEF assisted juice production in industrial scale. Innov. Food Sci. Emerg. Technol. 2012, 14, 46–60. [Google Scholar] [CrossRef]
- Saldaña, G.; Puértolas, E.; Álvarez, I.; Meneses, N.; Knorr, D.; Raso, J. Evaluation of a static treatment chamber to investigate kinetics of microbial inactivation by pulsed electric fields at different temperatures at quasi-isothermal conditions. J. Food Eng. 2010, 100, 349–356. [Google Scholar] [CrossRef]
- Knorr, D.; Froehling, A.; Jaeger, H.; Reineke, K.; Schlueter, O.; Schoessler, K. Emerging Technologies in Food Processing. Annu. Rev. Food Sci. Technol. 2011, 2, 203–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evrendilek, G.A.; Altuntas, J.; Sangun, M.K.; Zhang, H.Q. Apricot Nectar Processing by Pulsed Electric Fields. Int. J. Food Prop. 2013, 16, 216–227. [Google Scholar] [CrossRef]
- Sobrino-López, A.; Martin-Belloso, O. Review: Potential of High-Intensity Pulsed Electric Field Technology for Milk Processing. Food Eng. Rev. 2009, 2, 17–27. [Google Scholar] [CrossRef]
- Amiali, M.; Ngadi, M.O.; Smith, J.P.; Raghavan, V.G.S. Inactivation of Escherichia coli O157:H7 and Salmonella enteritidis in Liquid Egg White Using Pulsed Electric Field. J. Food Sci. 2006, 71, M88–M94. [Google Scholar] [CrossRef]
- Puértolas, E.; López, N.; Condón, S.; Raso, J.; Álvarez, I. Pulsed electric fields inactivation of wine spoilage yeast and bacteria. Int. J. Food Microbiol. 2009, 130, 49–55. [Google Scholar] [CrossRef]
- Neumann, E.; Schaefer-Ridder, M.; Wang, Y.; Hofschneider, P. Gene transfer into mouse lyoma cells by electroporation in high electric fields. EMBO J. 1982, 1, 841–845. [Google Scholar] [CrossRef] [PubMed]
- Ereineke, K.; Eschottroff, F.; Emeneses, N.; Eknorr, D. Sterilization of liquid foods by pulsed electric fields–an innovative ultra-high temperature process. Front. Microbiol. 2015, 6, 400. [Google Scholar] [CrossRef] [Green Version]
- Alkhafaji, S.R.; Farid, M. An investigation on pulsed electric fields technology using new treatment chamber design. Innov. Food Sci. Emerg. Technol. 2007, 8, 205–212. [Google Scholar] [CrossRef]
- Guionet, A.; David, F.; Zaepffel, C.; Coustets, M.; Helmi, K.; Cheype, C.; Packan, D.; Garnier, J.-P.; Blanckaert, V.; Teissié, J.E. coli electroeradication on a closed loop circuit by using milli-, micro- and nanosecond pulsed electric fields: Comparison between energy costs. Bioelectrochemistry 2015, 103, 65–73. [Google Scholar] [CrossRef]
- Coustets, M.; Ganeva, V.; Galutzov, B.; Teissie, J. Millisecond duration pulses for flow-through electro-induced protein extraction from E. coli and associated eradication. Bioelectrochemistry 2015, 103, 82–91. [Google Scholar] [CrossRef]
- Emanuel, E.; Roman, P.; Cahan, R. Influence of the current density in moderate pulsed electric fields on P. putida F1 eradication. Bioelectrochemistry 2019, 126, 172–179. [Google Scholar] [CrossRef]
- Emanuel, E.; Dubrovin, I.; Hanya, E.; Pinhasi, G.A.; Pogreb, R.; Cahan, R. Eradication of Saccharomyces cerevisiae by Pulsed Electric Field Treatments. Microorganisms 2020, 8, 1684. [Google Scholar] [CrossRef]
- French, D.M.; Uhler, M.D.; Gilgenbach, R.M.; Lau, Y.Y. Conductive versus capacitive coupling for cell electroporation with nanosecond pulses. J. Appl. Phys. 2009, 106, 074701. [Google Scholar] [CrossRef]
- Khan, S.I.; Blumrosen, G.; Vecchio, D.; Golberg, A.; Mccormack, M.C.; Yarmush, M.L.; Hamblin, M.R.; Austen, W.G. Eradication of multidrug-resistant pseudomonas biofilm with pulsed electric fields. Biotechnol. Bioeng. 2016, 113, 643–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahan, R.; Stein, M.; Anker, Y.; Langzam, Y.; Nitzan, Y. Innovative utilization of coal bottom ash for bioremediation of toxic organic pollutants. Int. Biodeterior. Biodegrad. 2013, 85, 421–428. [Google Scholar] [CrossRef]
- Raso, J.; Frey, W.; Ferrari, G.; Pataro, G.; Knorr, D.; Teissie, J.; Miklavčič, D. Recommendations guidelines on the key information to be reported in studies of application of PEF technology in food and biotechnological processes. Innov. Food Sci. Emerg. Technol. 2016, 37, 312–321. [Google Scholar] [CrossRef] [Green Version]
- COMSOL Multiphysics, Documentation for COMSOL Release 5.4 2019. Available online: https://www.comsol.com/ (accessed on 1 March 2021).
- Grahl, T.; Markl, H. Killing of microorganisms by pulsed electric fields. Appl. Microbiol. Biotechnol. 1996, 45, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Michael, E.; Gomila, M.; Lalucat, J.; Nitzan, Y.; Pechatnikov, I.; Cahan, R. Proteomic Assessment of the Expression of Genes Related to Toluene Catabolism and Porin Synthesis inPseudomonas stutzeriST-9. J. Proteome Res. 2017, 16, 1683–1692. [Google Scholar] [CrossRef]
- Papadopoulou, O.S.; Iliopoulos, V.; Mallouchos, A.; Panagou, E.Z.; Chorianopoulos, N.; Tassou, C.C.; Nychas, G.-J.E. Spoilage Potential of Pseudomonas (P. fragi, P. putida) and LAB (Leuconostoc mesenteroides, Lactobacillus sakei) Strains and Their Volatilome Profile during Storage of Sterile Pork Meat Using GC/MS and Data Analytics. Foods 2020, 9, 633. [Google Scholar] [CrossRef] [PubMed]
- Patarata, L.; Novais, M.; Fraqueza, M.J.; Silva, J.A. Influence of Meat Spoilage Microbiota Initial Load on the Growth and Survival of Three Pathogens on a Naturally Fermented Sausage. Foods 2020, 9, 676. [Google Scholar] [CrossRef]
- Jayaram, S.; Castle, G.; Margaritis, A. The effects of high field DC pulse and liquid medium conductivity on survivability of Lactobacillus brevis. Appl. Microbiol. Biotechnol. 1993, 40, 117–122. [Google Scholar] [CrossRef]
- Siemer, C.; Toepfl, S.; Heinz, V. Inactivation of Bacillus subtilis spores by pulsed electric fields (PEF) in combination with thermal energy—I. Influence of process- and product parameters. Food Control. 2014, 39, 163–171. [Google Scholar] [CrossRef]
- Böckmann, R.A.; Grubmüller, H. Multistep Binding of Divalent Cations to Phospholipid Bilayers: A Molecular Dynamics Study. Angew. Chem. Int. Ed. 2004, 43, 1021–1024. [Google Scholar] [CrossRef] [PubMed]
- Sachs, J.N.; Nanda, H.; Petrache, H.I.; Woolf, T.B. Changes in Phosphatidylcholine Headgroup Tilt and Water Order Induced by Monovalent Salts: Molecular Dynamics Simulations. Biophys. J. 2004, 86, 3772–3782. [Google Scholar] [CrossRef] [Green Version]
- Muraji, M.; Tatebe, W.; Berg, H. The influence of extracellular alkali and alkaline-earth ions on electropermeation of Saccharomyces cerevisiae. Bioelectrochem. Bioenerg. 1998, 46, 293–295. [Google Scholar] [CrossRef]
- Hülsheger, H.; Potel, J.; Niemann, E.G. Electric field effects on bacteria and yeast cells. Radiat. Environ. Biophys. 1983, 22, 149–162. [Google Scholar] [CrossRef] [PubMed]
- García, D.; Gomez, N.; Raso, J.; Pagán, R. Bacterial resistance after pulsed electric fields depending on the treatment medium pH. Innov. Food Sci. Emerg. Technol. 2005, 6, 388–395. [Google Scholar] [CrossRef]
- Cregenzán-Alberti, O.; Halpin, R.; Whyte, P.; Lyng, J.; Noci, F. Study of the suitability of the central composite design to predict the inactivation kinetics by pulsed electric fields (PEF) in Escherichia coli, Staphylococcus aureus and Pseudomonas fluorescens in milk. Food Bioprod. Process. 2015, 95, 313–322. [Google Scholar] [CrossRef]
- Sharma, P.; Bremer, P.; Oey, I.; Everett, D. Bacterial inactivation in whole milk using pulsed electric field processing. Int. Dairy J. 2014, 35, 49–56. [Google Scholar] [CrossRef]
- Craven, H.; Swiergon, P.; Ng, S.; Midgely, J.; Versteeg, C.; Coventry, M.; Wan, J. Evaluation of pulsed electric field and minimal heat treatments for inactivation of pseudomonads and enhancement of milk shelf-life. Innov. Food Sci. Emerg. Technol. 2008, 9, 211–216. [Google Scholar] [CrossRef]
- Zhang, S.; Ye, C.; Lin, H.; Lv, L.; Yu, X. UV Disinfection induces a VBNC state in Escherichia coli and Pseudomonas aerugino-sa. Environ. Sci. Technol. 2015, 49, 1721–1728. [Google Scholar] [CrossRef]
- Lavieri, N.A.; Sebranek, J.G.; Cordray, J.C.; Dickson, J.S.; Jung, S.; Manu, D.K.; Mendonça, A.F.; Brehm-Stecher, B.F.; Stock, J.; Stalder, K.J. Evaluation of the Thin Agar Layer Method for the Recovery of Pressure-Injured and Heat-Injured Listeria monocytogenes. J. Food Prot. 2014, 77, 828–831. [Google Scholar] [CrossRef]
- Divol, B.; Lonvaud-Funel, A. Evidence for viable but nonculturable yeasts in botrytis-affected wine. J. Appl. Microbiol. 2005, 99, 85–93. [Google Scholar] [CrossRef]
- Asakura, H.; Igimi, S.; Kawamoto, K.; Yamamoto, S.; Makino, S.-I. Role of in vivo passage on the environmental adaptation of enterohemorrhagicEscherichia coliO157:H7: Cross-induction of the viable but nonculturable state by osmotic and oxidative stresses. FEMS Microbiol. Lett. 2005, 253, 243–249. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Günther, S.; Hübschmann, T.; Wick, L.Y.; Harms, H.; Müller, S. Limits of propidium iodide as a cell viability indicator for environmental bacteria. Cytom. Part A 2007, 71, 592–598. [Google Scholar] [CrossRef]
- Wang, M.-S.; Wang, L.-H.; Bekhit, A.E.-D.A.; Yang, J.; Hou, Z.-P.; Wang, Y.-Z.; Dai, Q.-Z.; Zeng, X.-A. A review of sublethal effects of pulsed electric field on cells in food processing. J. Food Eng. 2018, 223, 32–41. [Google Scholar] [CrossRef]
- García, D.; Gómez, N.; Mañas, P.; Raso, J.; Pagán, R. Pulsed electric fields cause bacterial envelopes permeabilization depending on the treatment intensity, the treatment medium pH and the microorganism investigated. Int. J. Food Microbiol. 2007, 113, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Somolinos, M.; Mañas, P.; Condón, S.; Pagán, R.; García, D. Recovery of Saccharomyces cerevisiae sublethally injured cells after Pulsed Electric Fields. Int. J. Food Microbiol. 2008, 125, 352–356. [Google Scholar] [CrossRef] [PubMed]
- García, D.; Mañas, P.; Gomez, N.; Raso, J.; Pagán, R. Biosynthetic requirements for the repair of sublethal membrane damage in Escherichia coli cells after pulsed electric fields. J. Appl. Microbiol. 2006, 100, 428–435. [Google Scholar] [CrossRef]
- Arroyo, C.; Somolinos, M.; Cebrián, G.; Condón, S.; Pagán, R. Pulsed electric fields cause sublethal injuries in the outer membrane of Enterobacter sakazakii facilitating the antimicrobial activity of citral. Lett. Appl. Microbiol. 2010, 51, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Mishra, Y.; Chaurasia, N.; Rai, L.C. AhpC (alkyl hydroperoxide reductase) from Anabaena sp. PCC 7120 protects Escherichia coli from multiple abiotic stresses. Biochem. Biophys. Res. Commun. 2009, 381, 606–611. [Google Scholar] [CrossRef]
- Zuo, F.; Yu, R.; Khaskheli, G.B.; Ma, H.; Chen, L.; Zeng, Z.; Mao, A.; Chen, S. Homologous overexpression of alkyl hydroperoxide reductase subunit C (ahpC) protects Bifidobacterium longum strain NCC2705 from oxidative stress. Res. Microbiol. 2014, 165, 581–589. [Google Scholar] [CrossRef]
- Noinaj, N.; Guillier, M.; Barnard, T.J.; Buchanan, S.K. TonB-Dependent Transporters: Regulation, Structure, and Function. Annu. Rev. Microbiol. 2010, 64, 43–60. [Google Scholar] [CrossRef] [Green Version]
- Zaborin, A.; Gerdes, S.; Holbrook, C.; Liu, D.C.; Zaborina, O.Y.; Alverdy, J.C. Pseudomonas aeruginosa Overrides the Virulence Inducing Effect of Opioids When It Senses an Abundance of Phosphate. PLoS ONE 2012, 7, e34883. [Google Scholar] [CrossRef]
- Monedero, V.; Revilla-Guarinos, A.; Zúñiga, M. Physiological Role of Two-Component Signal Transduction Systems in Food-Associated Lactic Acid Bacteria. Adv. Appl. Microbiol. 2017, 99, 1–51. [Google Scholar] [CrossRef] [PubMed]
- De Vrij, W.; Bulthuis, R.A.; Van Iwaarden, P.R.; Konings, W.N. Mechanism of L-glutamate transport in membrane vesicles from Bacillus stearothermophilus. J. Bacteriol. 1989, 171, 1118–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rallu, F.; Gruss, A.; Ehrlich, S.D.; Maguin, E. Acid- and multistress-resistant mutants of Lactococcus lactis: Identification of intracellular stress signals. Mol. Microbiol. 2002, 35, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Ling, N.; Gao, J.; Zhang, X.; Zhang, M.; Tong, L.; Zeng, H.; Zhang, J.; Wu, Q. Roles of outer membrane protein W (OmpW) on survival, morphology, and biofilm formation under NaCl stresses in Cronobacter sakazakii. J. Dairy Sci. 2018, 101, 3844–3850. [Google Scholar] [CrossRef]
- Morales, E.H.; Calderón, I.L.; Collao, B.; Gil, F.; Porwollik, S.; McClelland, M.; Saavedra, C.P. Hypochlorous acid and hydrogen peroxide-induced negative regulation of Salmonella enterica serovar Typhimurium ompW by the response regulator ArcA. BMC Microbiol. 2012, 12, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeats, C.; Rawlings, N.D.; Bateman, A. The PepSY domain: A regulator of peptidase activity in the microbial environment? Trends Biochem. Sci. 2004, 29, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Orelle, C.; Mathieu, K.; Jault, J.-M. Multidrug ABC transporters in bacteria. Res. Microbiol. 2019, 170, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Monaco, C.; Talà, A.; Spinosa, M.R.; Progida, C.; De Nitto, E.; Gaballo, A.; Bruni, C.B.; Bucci, C.; Alifano, P. Identification of a Meningococcal l-Glutamate ABC Transporter Operon Essential for Growth in Low-Sodium Environments. Infect. Immun. 2006, 74, 1725–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bickle, T.A. Restricting restriction. Mol. Microbiol. 2003, 51, 3–5. [Google Scholar] [CrossRef]
- Ziegler, C.M.; Bremer, E.; Krämer, R. The BCCT family of carriers: From physiology to crystal structure. Mol. Microbiol. 2010, 78, 13–34. [Google Scholar] [CrossRef]



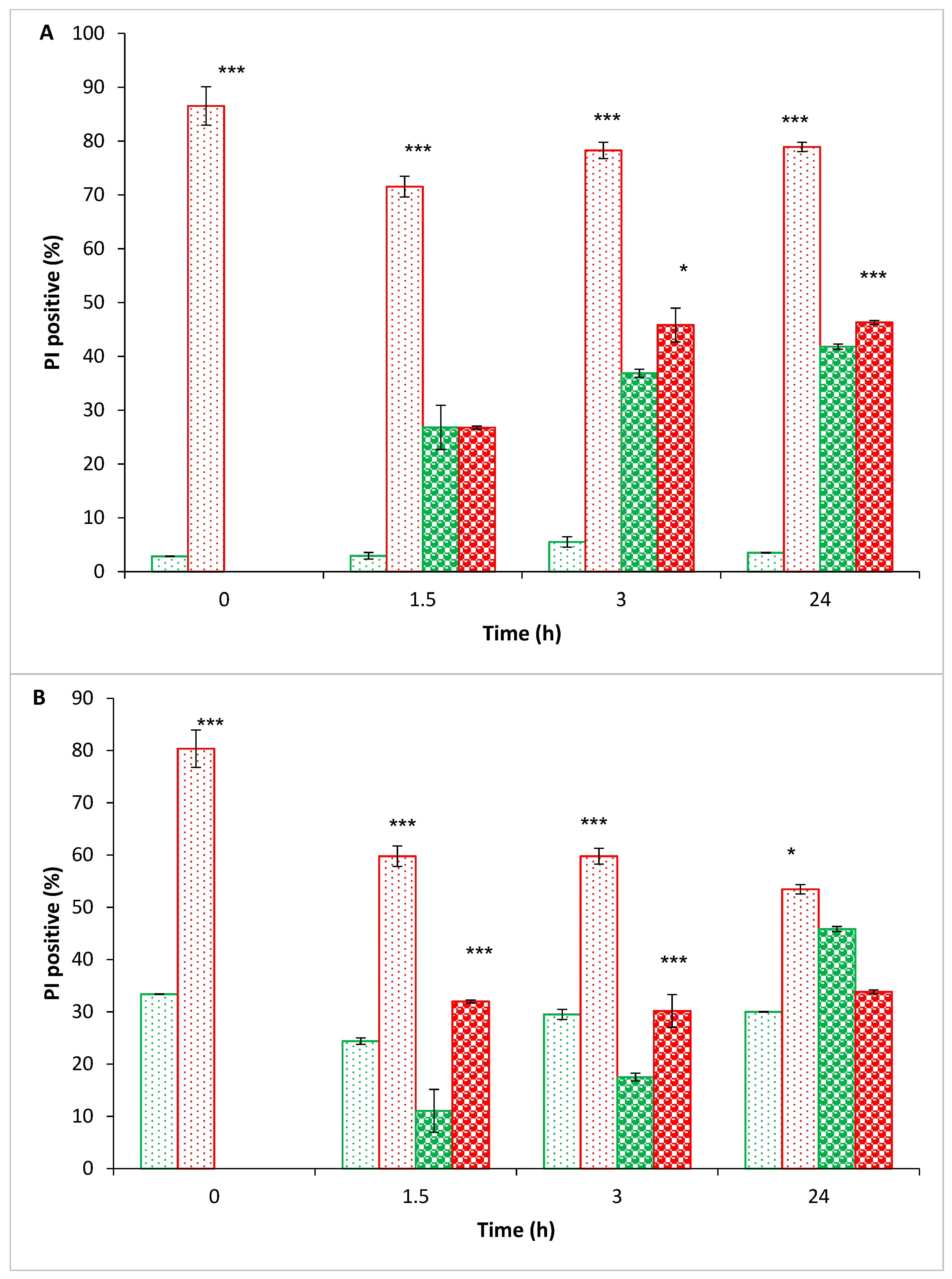
 ); non-treated bacteria in time ‘0’ (
); non-treated bacteria in time ‘0’ (  ); PEF-treated bacteria in brain-heart infusion (BHI) (1.5–24 h) (
); PEF-treated bacteria in brain-heart infusion (BHI) (1.5–24 h) (  ); PEF-treated bacteria in phosphate buffer saline (PBS) (1.5–24 h) (
); PEF-treated bacteria in phosphate buffer saline (PBS) (1.5–24 h) (  ); nontreated bacteria in BHI (1.5–24 h) (
); nontreated bacteria in BHI (1.5–24 h) (  ); non-treated bacteria in PBS (1.5–24 h) (
); non-treated bacteria in PBS (1.5–24 h) (  ). p value (t test): Significance of the CFU count in each examined time (PEF-treated bacteria in BHI or in BPS) related to its control (nontreated bacteria in BHI or PBS, respectively) p < 0.001 ***; significance of the CFU of the treated-bacteria in PBS related to treated-bacteria in BHI, in each examined time p < 0.01 ##; p < 0.001 ###.
); non-treated bacteria in time ‘0’ ( ); PEF-treated bacteria in brain-heart infusion (BHI) (1.5–24 h) ( ); PEF-treated bacteria in phosphate buffer saline (PBS) (1.5–24 h) ( ); nontreated bacteria in BHI (1.5–24 h) ( ); non-treated bacteria in PBS (1.5–24 h) ( ). p value (t test): Significance of the CFU count in each examined time (PEF-treated bacteria in BHI or in BPS) related to its control (nontreated bacteria in BHI or PBS, respectively) p < 0.001 ***; significance of the CFU of the treated-bacteria in PBS related to treated-bacteria in BHI, in each examined time p < 0.01 ##; p < 0.001 ###.
). p value (t test): Significance of the CFU count in each examined time (PEF-treated bacteria in BHI or in BPS) related to its control (nontreated bacteria in BHI or PBS, respectively) p < 0.001 ***; significance of the CFU of the treated-bacteria in PBS related to treated-bacteria in BHI, in each examined time p < 0.01 ##; p < 0.001 ###.
); non-treated bacteria in time ‘0’ ( ); PEF-treated bacteria in brain-heart infusion (BHI) (1.5–24 h) ( ); PEF-treated bacteria in phosphate buffer saline (PBS) (1.5–24 h) ( ); nontreated bacteria in BHI (1.5–24 h) ( ); non-treated bacteria in PBS (1.5–24 h) ( ). p value (t test): Significance of the CFU count in each examined time (PEF-treated bacteria in BHI or in BPS) related to its control (nontreated bacteria in BHI or PBS, respectively) p < 0.001 ***; significance of the CFU of the treated-bacteria in PBS related to treated-bacteria in BHI, in each examined time p < 0.01 ##; p < 0.001 ###.
 ); PEF-treated bacteria which were suspended in PBS (0–24 h) (
); PEF-treated bacteria which were suspended in PBS (0–24 h) (  ); nontreated bacteria which were diluted in BHI (1.5–24 h) (
); nontreated bacteria which were diluted in BHI (1.5–24 h) (  ); nontreated bacteria which were suspended in PBS (0–24 h) (
); nontreated bacteria which were suspended in PBS (0–24 h) (  ). Propidium iodide (PI)-positive control using ethanol was 90 ± 3%. p values (t-test): p < 0.05 *; p < 0.001 ***.
); PEF-treated bacteria which were suspended in PBS (0–24 h) ( ); nontreated bacteria which were diluted in BHI (1.5–24 h) ( ); nontreated bacteria which were suspended in PBS (0–24 h) ( ). Propidium iodide (PI)-positive control using ethanol was 90 ± 3%. p values (t-test): p < 0.05 *; p < 0.001 ***.
). Propidium iodide (PI)-positive control using ethanol was 90 ± 3%. p values (t-test): p < 0.05 *; p < 0.001 ***.
); PEF-treated bacteria which were suspended in PBS (0–24 h) ( ); nontreated bacteria which were diluted in BHI (1.5–24 h) ( ); nontreated bacteria which were suspended in PBS (0–24 h) ( ). Propidium iodide (PI)-positive control using ethanol was 90 ± 3%. p values (t-test): p < 0.05 *; p < 0.001 ***.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein IDs | Protein Name | Gene Name | Mol. Weight [kDa] | log2 * LFQ Intensity Nontreated | log2 * LFQ Intensity PEF-Treated |
|---|---|---|---|---|---|
| A5W5H2 | Alkyl hydroperoxide reductase/Thiol specific antioxidant/Mal allergen | Pput_3256 | 20.507 | 28.96 ± 0.43 | 33.33 ± 0.33 |
| A5VXD9 | TonB-dependent siderophore receptor | Pput_0376 | 88.459 | 19.18 ± 1.94 | 24.63 ± 1.18 |
| A5W124 | TonB-dependent siderophore receptor | Pput_1678 | 90.426 | ND | 24.12 ± 0.28 |
| A5W341 | TonB-dependent siderophore receptor | Pput_2412 | 79.564 | ND | 21.21 ± 0.73 |
| A5VZF9 | Integral membrane sensor signal transduction histidine kinase | Pput_1108 | 70.283 | ND | 21.84 ± 1.71 |
| A5VWS0 | Probable proton/glutamate-aspartate symporter | gltP | 47.566 | 21.96 ± 0.55 | 25.61 ± 0.18 |
| A5VXU7 | OmpW family protein | Pput_0539 | 24.19 | ND | 26.06 ± 1.47 |
| A5W1U8 | ABC-type metal ion transport system periplasmic component/surface adhesin-like protein | Pput_1965 | 30.162 | 18.25 ± 2.17 | 22.68 ± 0.16 |
| A5W1V5 | Propeptide, PepSY and peptidase M4 | Pput_1972 | 44.145 | ND | 24.35 ± 1.01 |
| A5VZG2 | L-glutamate ABC transporter membrane protein/L-aspartate ABC transporter membrane protein | Pput_1111 | 27.45 | 17.90 ± 1.57 | 22.14 ± 1.10 |
| A5W312 | Deoxyribonuclease I | Pput_2383 | 35.477 | ND | 21.05 ± 0.18 |
| A5WA94 | Choline/carnitine/betaine transporter | Pput_4934 | 73.501 | 20.91 ± 0.14 | 24.61 ± 0.52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emanuel, E.; Dubrovin, I.; Pogreb, R.; Pinhasi, G.A.; Cahan, R. Resuscitation of Pulsed Electric Field-Treated Staphylococcus aureus and Pseudomonas putida in a Rich Nutrient Medium. Foods 2021, 10, 660. https://doi.org/10.3390/foods10030660
Emanuel E, Dubrovin I, Pogreb R, Pinhasi GA, Cahan R. Resuscitation of Pulsed Electric Field-Treated Staphylococcus aureus and Pseudomonas putida in a Rich Nutrient Medium. Foods. 2021; 10(3):660. https://doi.org/10.3390/foods10030660
Chicago/Turabian StyleEmanuel, Efrat, Irina Dubrovin, Roman Pogreb, Gad A. Pinhasi, and Rivka Cahan. 2021. "Resuscitation of Pulsed Electric Field-Treated Staphylococcus aureus and Pseudomonas putida in a Rich Nutrient Medium" Foods 10, no. 3: 660. https://doi.org/10.3390/foods10030660
APA StyleEmanuel, E., Dubrovin, I., Pogreb, R., Pinhasi, G. A., & Cahan, R. (2021). Resuscitation of Pulsed Electric Field-Treated Staphylococcus aureus and Pseudomonas putida in a Rich Nutrient Medium. Foods, 10(3), 660. https://doi.org/10.3390/foods10030660