
Protein Profile and Simulated Digestive Behavior of Breast Milk from Overweight and Normal Weight Mothers

 ,
,  , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Subjects and Samples
2.2. MALDI-TOF Protein Profile
2.3. In Vitro Simulated Gastrointestinal Digestion
2.4. Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis (SDS-PAGE) and Identification of Bands by In-Gel Digestion
2.5. Analysis by Ultra-Performance Liquid Chromatography (UPLC)
2.6. Analysis of Digests by HPLC–Tandem Mass Spectrometry (HPLC–MS/MS)
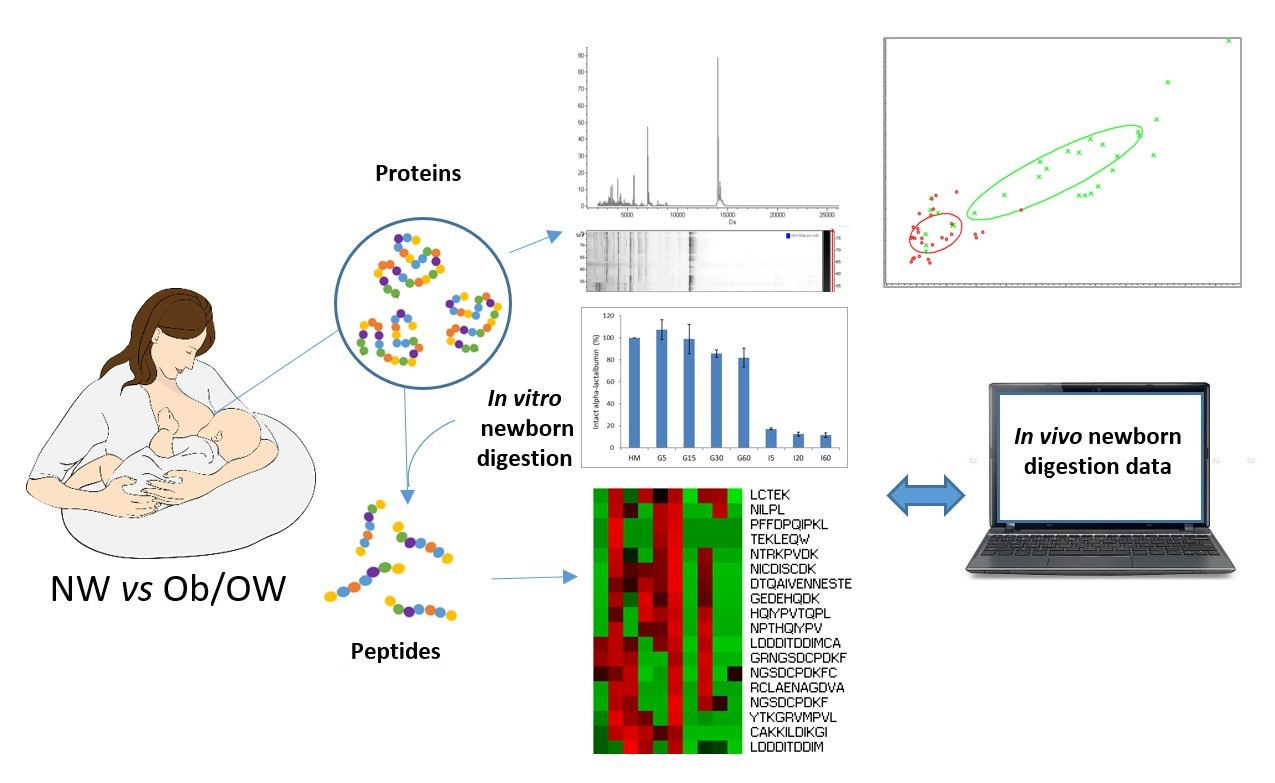
3. Results and Discussion
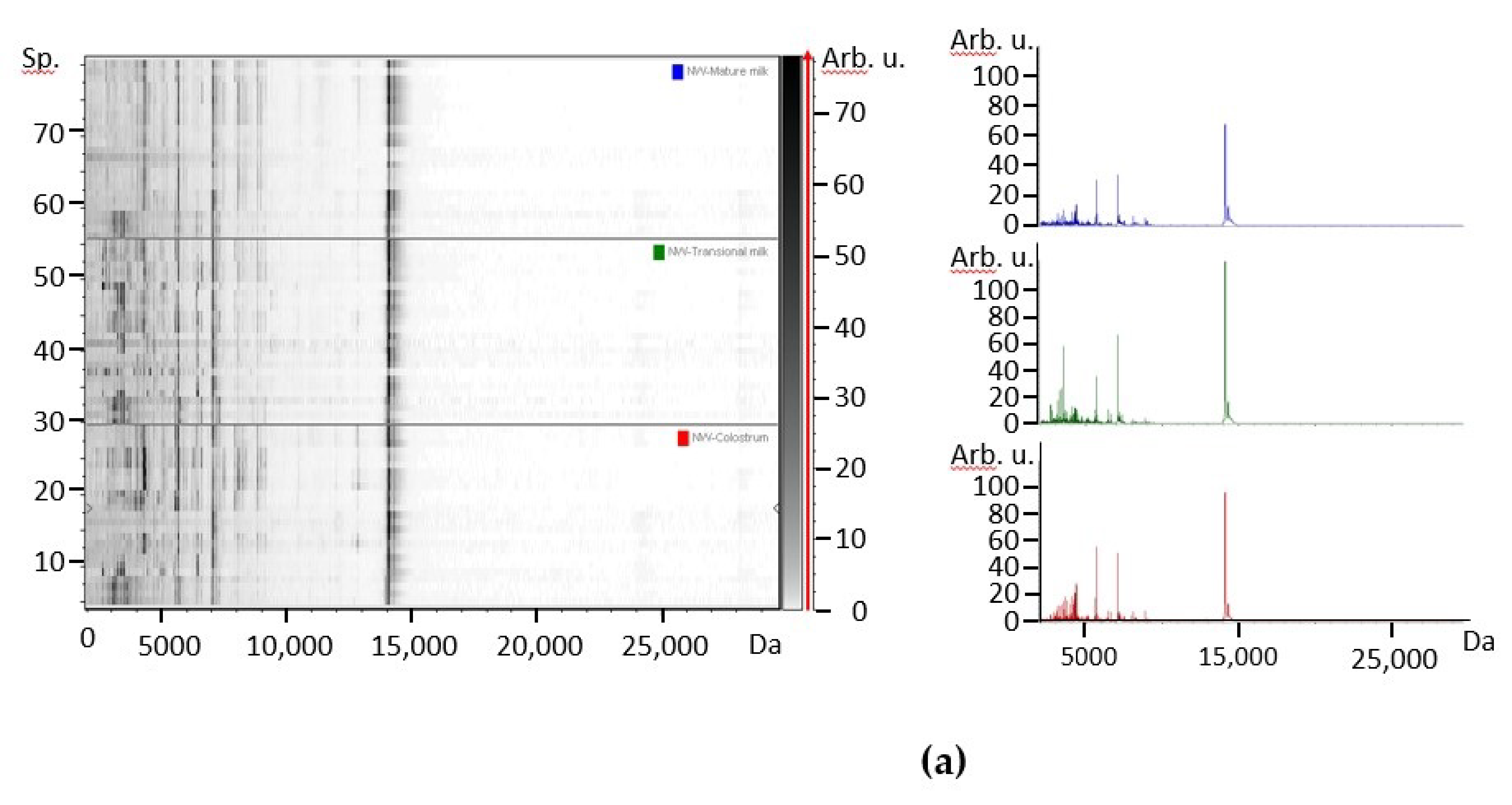
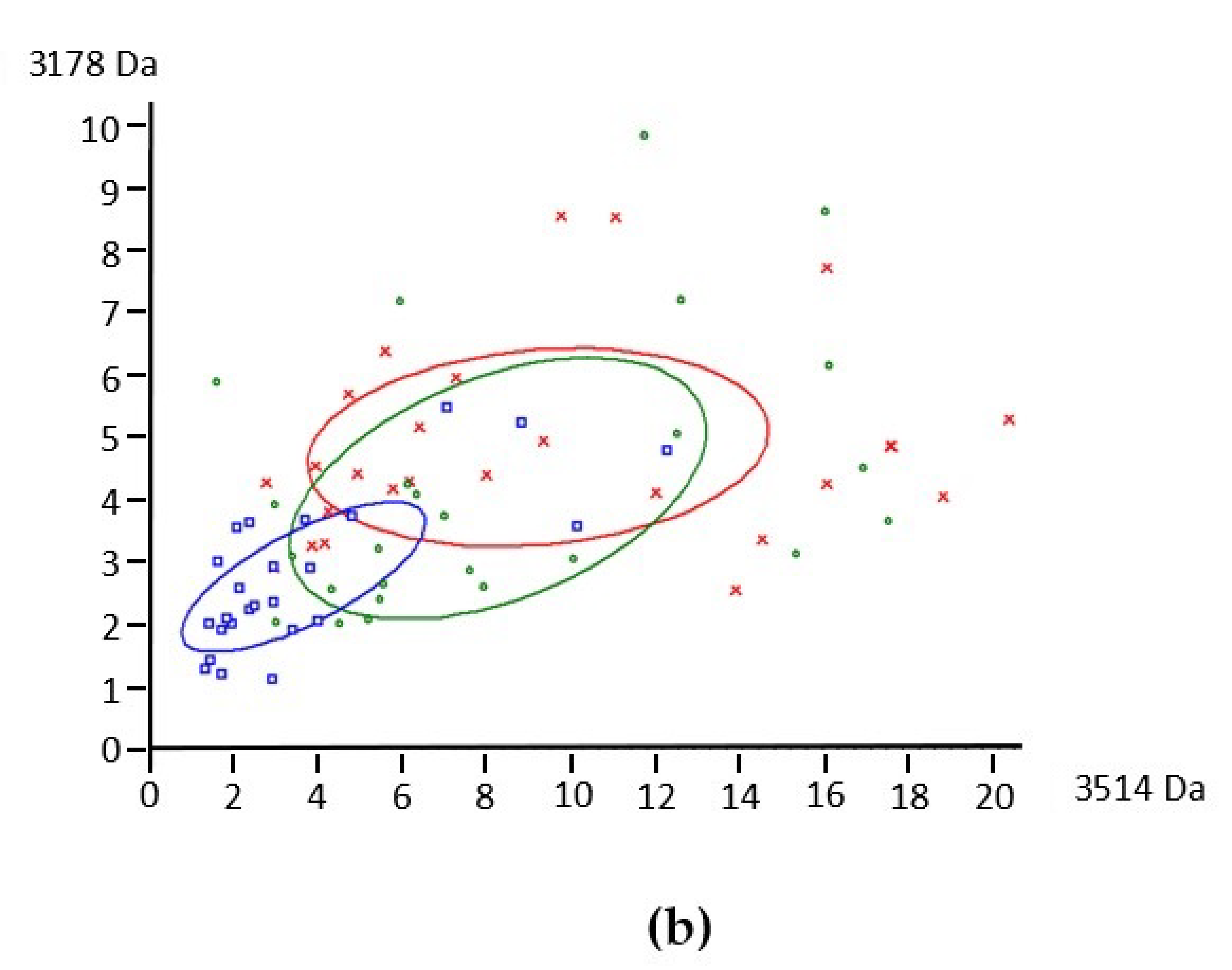
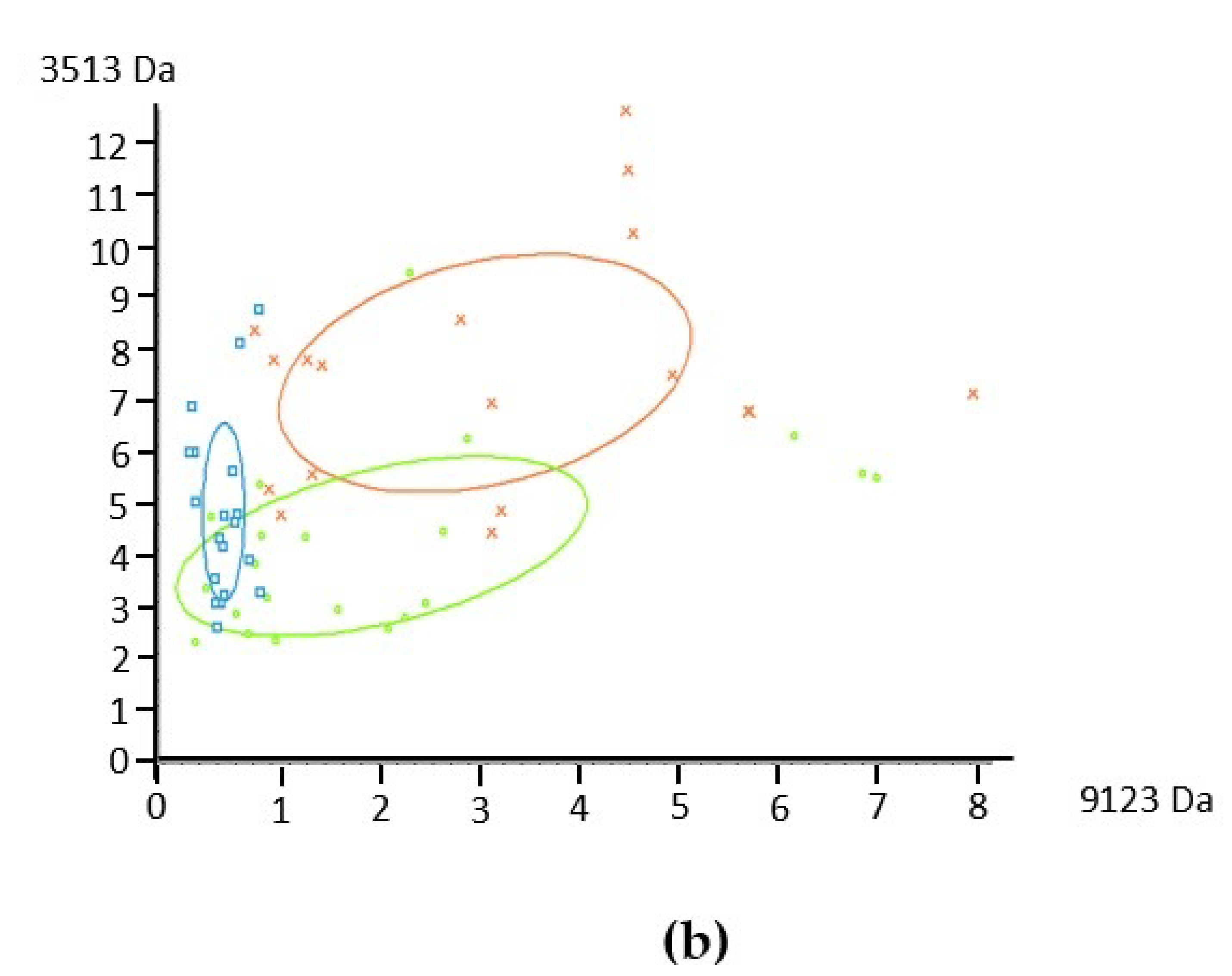
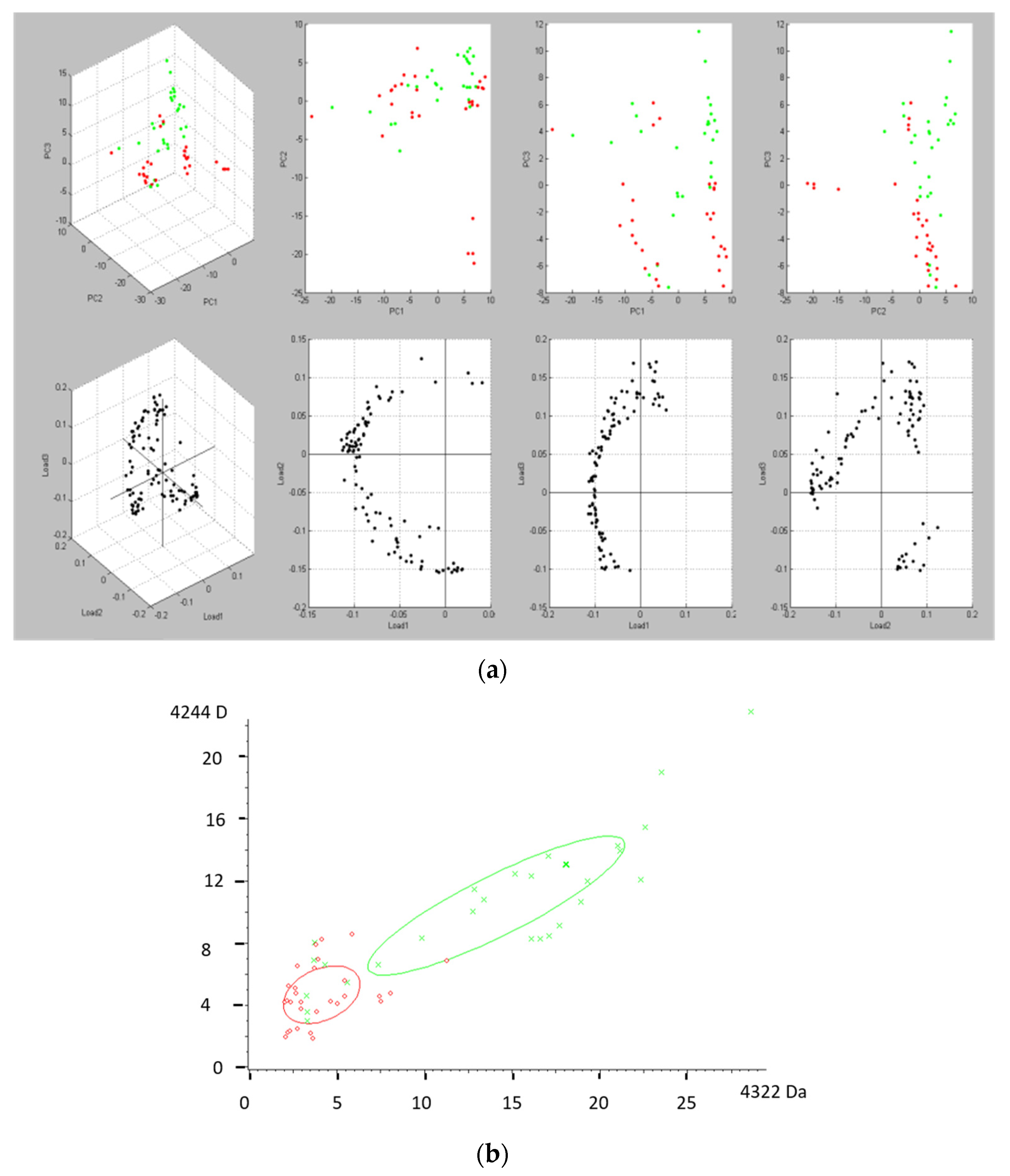
3.1. Protein Profile of Human Milks by MALDI-TOF Mass Spectrometry
3.2. Protein Degradation of Human Milk during Infant In Vitro Gastrointestinal Digestion
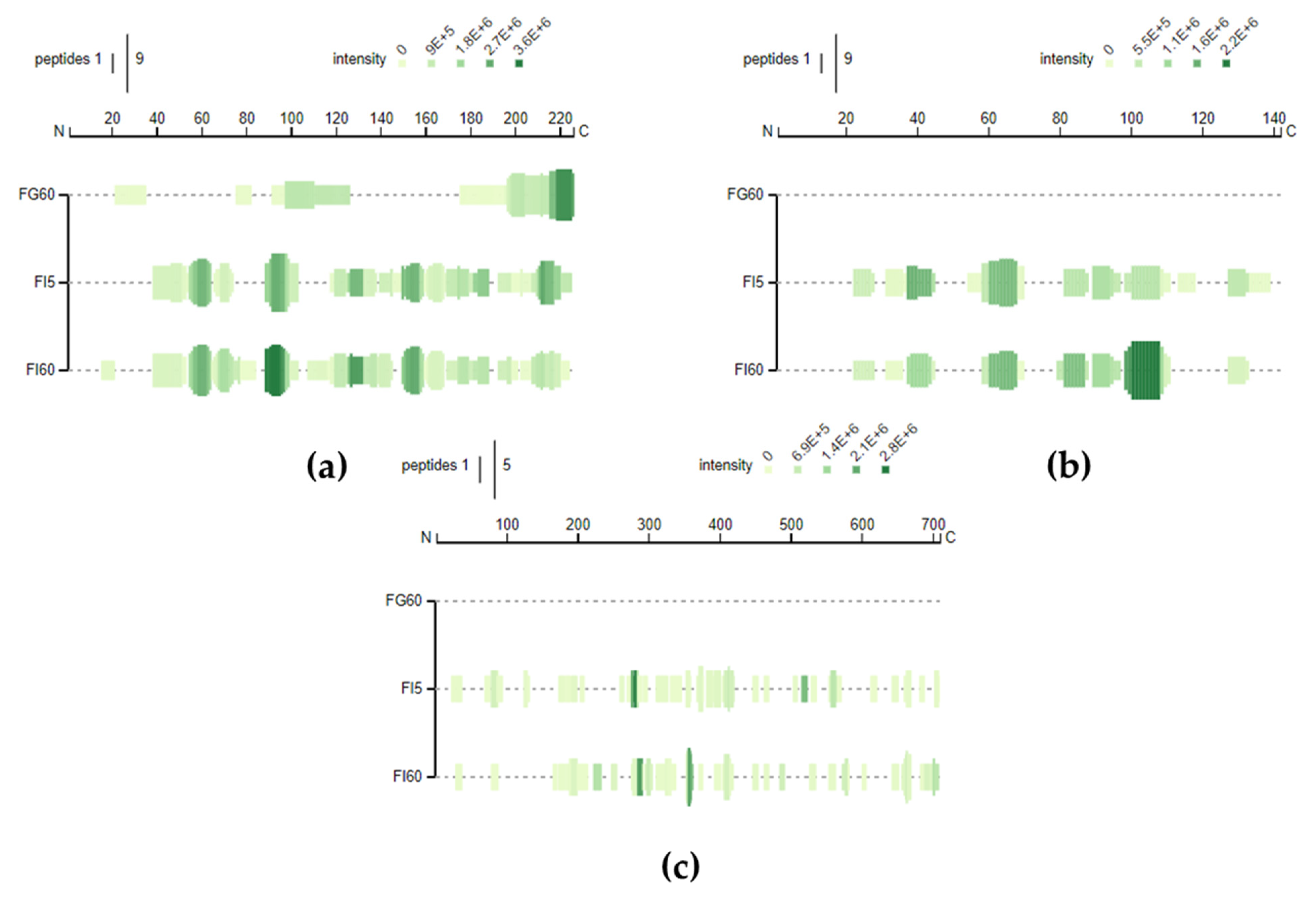
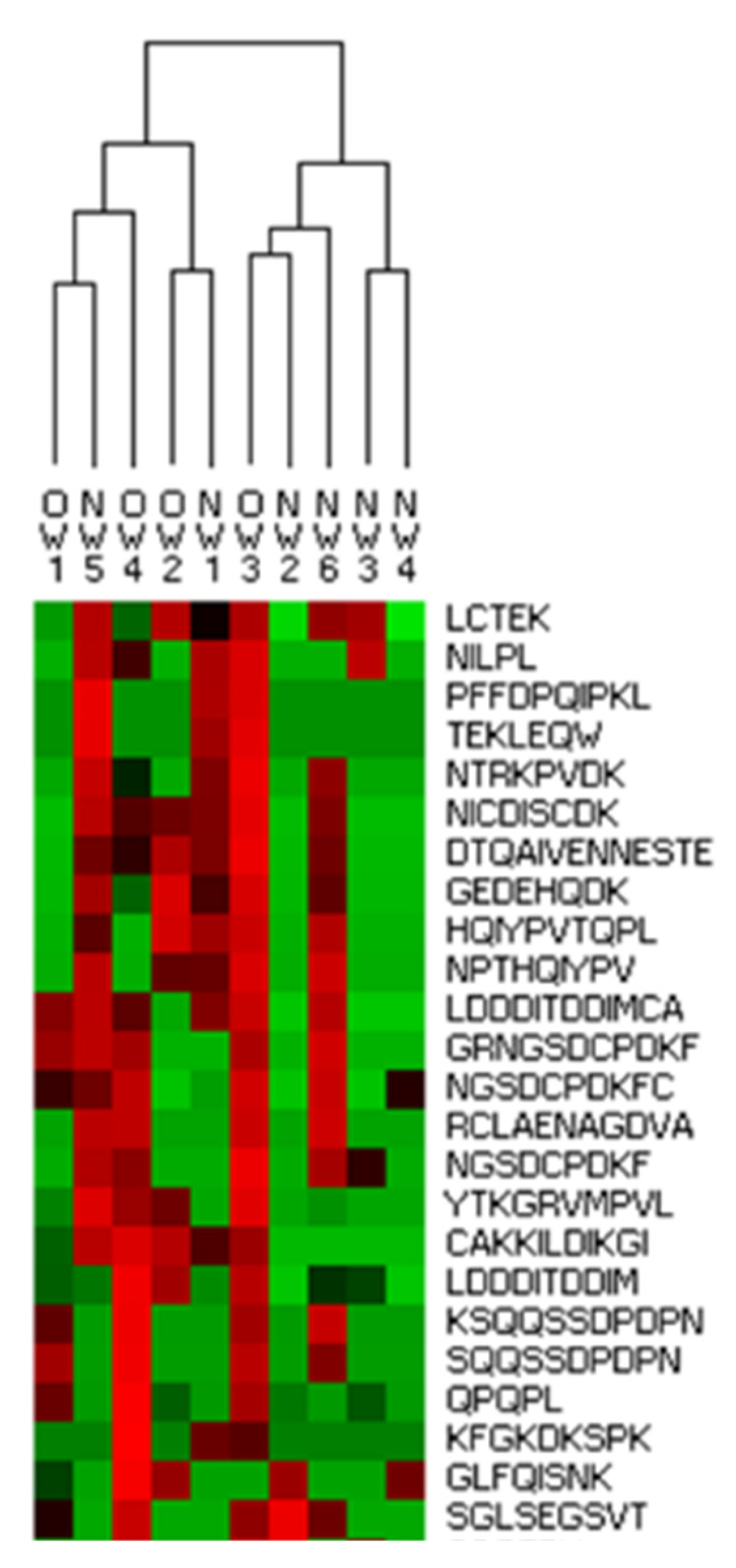
3.3. Peptidomic Characterization of In Vitro Digests
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Butte, N.F.; Lopez-Alarcon, M.G.; Garza, C. Nutrient Adequacy of Exclusive Breastfeeding for the Term Infant During the First Six Months of Life; WHO: Geneva, Switzerland, 2002; Available online: https://apps.who.int/iris/handle/10665/42519 (accessed on 14 February 2018).
- Lönnerdal, B. Bioactive Proteins in Human Milk: Health, Nutrition, and Implications for Infant Formulas. J. Pediatr. 2016, 173, S4–S9. [Google Scholar] [CrossRef]
- Zepf, F.D.; Rao, P.; Moore, J.; Stewart, R.; Ladino, Y.M.; Hartmann, B.T. Human breast milk and adipokines—A potential role for the soluble leptin receptor (sOb-R) in the regulation of infant energy intake and development. Med. Hypotheses 2016, 86, 53–55. [Google Scholar] [CrossRef] [PubMed]
- Lönnerdal, B. Infant formula and infant nutrition: Bioactive proteins of human milk and implications for composition of infant formulas. Am. J. Clin. Nutr. 2014, 99, 712S–717S. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Weber, D.; Xu, W.; Durbin-Johnson, B.P.; Phinney, B.S.; Lönnerdal, B. Absolute Quantification of Human Milk Caseins and the Whey/Casein Ratio during the First Year of Lactation. J. Proteome Res. 2017, 16, 4113–4121. [Google Scholar] [CrossRef]
- Ren, Q.; Zhou, Y.; Zhang, W.; Tian, Y.; Sun, H.; Zhao, X.; Xu, Y.; Jiang, S. Longitudinal changes in the bioactive proteins in human milk of the Chinese population: A systematic review. Food Sci. Nutr. 2021, 9, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, S.; Boylan, M.; Hart, S.L.; Román-Shriver, C.; Spallholz, J.E.; Pence, B.C.; Sawyer, B.G. Glucose and Insulin Levels are Increased in Obese and Overweight Mothers’ Breast-Milk. Food Nutr. Sci. 2014, 2, 201–206. [Google Scholar] [CrossRef]
- Andreas, N.J.; Hyde, M.J.; Gale, C.; Parkinson, J.R.C.; Jeffries, S.; Holmes, E.; Modi, N. Effect of Maternal Body Mass Index on Hormones in Breast Milk: A Systematic Review. PLoS ONE 2014, 9, e115043. [Google Scholar] [CrossRef]
- Saben, J.L.; Sims, C.R.; Piccolo, B.D.; Andres, A. Maternal adiposity alters the human milk metabolome: Associations between nonglucose monosaccharides and infant adiposity. Am. J. Clin. Nutr. 2020, 112, 1228–1239. [Google Scholar] [CrossRef]
- Young, B.E.; Patinkin, Z.; Palmer, C.; De La Houssaye, B.; Barbour, L.A.; Hernandez, T.; Friedman, J.E.; Krebs, N.F. Human milk insulin is related to maternal plasma insulin and BMI: But other components of human milk do not differ by BMI. Eur. J. Clin. Nutr. 2017, 71, 1094–1100. [Google Scholar] [CrossRef]
- Leghi, G.E.; Netting, M.J.; Middleton, P.F.; Wlodek, M.E.; Geddes, D.T.; Muhlhausler, B.S. The impact of maternal obesity on human milk macronutrient composition: A systematic review and meta-analysis. Nutrients 2020, 12, 934. [Google Scholar] [CrossRef]
- Bourlieu, C.; Deglaire, A.; de Oliveira, S.C.; Ménard, O.; Le Gouar, Y.; Carrière, F.; Dupont, D. Towards infant formula biomimetic of human milk structure and digestive behaviour. OCL 2017, 24, D206. [Google Scholar] [CrossRef]
- Devle, H.; Ulleberg, E.K.; Naess-Andresen, C.F.; Rukke, E.O.; Vegarud, G.; Ekeberg, D. Reciprocal interacting effects of proteins and lipids during exvivo digestion of bovine milk. Int. Dairy J. 2014, 36, 6–13. [Google Scholar] [CrossRef]
- Dallas, D.C.; Guerrero, A.; Khaldi, N.; Borghese, R.; Bhandari, A.; Underwood, M.A.; Lebrilla, C.B.; German, J.B.; Barile, D. A Peptidomic Analysis of Human Milk Digestion in the Infant Stomach Reveals Protein-Specific Degradation Patterns. J. Nutr. 2014, 144, 815–820. [Google Scholar] [CrossRef]
- Nielsen, S.D.; Beverly, R.L.; Underwood, M.A.; Dallas, D.C. Release of functional peptides from mother’s milk and fortifier proteins in the premature infant stomach. PLoS ONE 2018, 13, e0208204. [Google Scholar] [CrossRef]
- Beverly, R.L.; Woonnimani, P.; Scottoline, B.P.; Lueangsakulthai, J.; Dallas, D.C. Peptides from the Intestinal Tract of Breast Milk-Fed Infants Have Antimicrobial and Bifidogenic Activity. Int. J. Mol. Sci. 2021, 22, 2377. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, L.; Di Girolamo, F.; Mennini, M.; Masotti, A.; Salvatori, G.; Rigon, G.; Signore, F.; Pietrantoni, E.; Scapaticci, M.; Lante, I.; et al. A MALDI-TOF MS approach for mammalian, human, and formula milks’ profiling. Nutrients 2018, 10, 1238. [Google Scholar] [CrossRef] [PubMed]
- Hammer, B.; Strickert, M.; Villmann, T. Supervised neural gas with general similarity measure. Neural Process. Lett. 2005, 21, 21–44. [Google Scholar] [CrossRef]
- Holland, J.H. Adaptation in Natural and Artificial Systems; University of Michigan Press: Ann Arbor, MI, USA, 1975. [Google Scholar]
- Ménard, O.; Bourlieu, C.; De Oliveira, S.C.; Dellarosa, N.; Laghi, L.; Carrière, F.; Capozzi, F.; Dupont, D.; Deglaire, A. A first step towards a consensus static in vitro model for simulating full-term infant digestion. Food Chem. 2018, 240, 338–345. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef] [PubMed]
- Miralles, B.; Sanchón, J.; Sánchez-Rivera, L.; Martínez-Maqueda, D.; Le Gouar, Y.; Dupont, D.; Amigo, L.; Recio, I. Digestion of micellar casein in duodenum cannulated pigs. Correlation between in vitro simulated gastric digestion and in vivo data. Food Chem. 2021, 343, 128424. [Google Scholar] [CrossRef] [PubMed]
- Visser, S.; Slangen, C.J.; Rollema, H.S. Phenotyping of bovine milk proteins by reversed-phase high-performance liquid chromatography. J. Chromatogr. A 1991, 548, 361–370. [Google Scholar] [CrossRef]
- Sánchez-Rivera, L.; Diezhandino, I.; Gómez-Ruiz, J.Á.; Fresno, J.M.; Miralles, B.; Recio, I. Peptidomic study of Spanish blue cheese (Valdeón) and changes after simulated gastrointestinal digestion. Electrophoresis 2014, 35, 1627–1636. [Google Scholar] [CrossRef]
- Oliveros, J.C. Venny 2.1.0. Available online: https://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 25 March 2021).
- Manguy, J.; Jehl, P.; Dillon, E.T.; Davey, N.E.; Shields, D.C.; Holton, T.A. Peptigram: A Web-Based Application for Peptidomics Data Visualization. J. Proteome Res. 2017, 16, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Caraux, G.; Pinloche, S. PermutMatrix: A graphical environment to arrange gene expression profiles in optimal linear order. Bioinformatics 2005, 21, 1280–1281. [Google Scholar] [CrossRef] [PubMed]
- Jose’stam, J.; Sauer, P.J.; Boehm, G. Can we define an infant’s need from the composition of human milk? Am. J. Clin. Nutr. 2013, 98, 521S–528S. [Google Scholar] [CrossRef]
- Young, B.E.; Levek, C.; Reynolds, R.M.; Rudolph, M.C.; MacLean, P.; Hernandez, T.L.; Friedman, J.E.; Krebs, N.F. Bioactive components in human milk are differentially associated with rates of lean and fat mass deposition in infants of mothers with normal vs. elevated BMI. Pediatr. Obes. 2018, 13, 598–606. [Google Scholar] [CrossRef]
- Kratzsch, J.; Bae, Y.J.; Kiess, W. Adipokines in human breast milk. Best Pract. Res. Clin. Endocrinol. Metab. 2018, 32, 27–38. [Google Scholar] [CrossRef]
- Young, B.E.; Johnson, S.L.; Krebs, N.F. Biological determinants linking infant weight gain and child obesity: Current knowledge and future directions. Adv. Nutr. 2012, 3, 675–686. [Google Scholar] [CrossRef]
- Lönnerdal, B. Bioactive Proteins in Human Milk: Mechanisms of Action. J. Pediatr. 2010, 156, S26–S30. [Google Scholar] [CrossRef]
- Baba, T.; Downs, D.; Jackson, K.W.; Tang, J.; Wang, C.S. Structure of Human Milk Bile Salt Activated Lipase. Biochemistry 1991, 30, 500–510. [Google Scholar] [CrossRef]
- de Oliveira, S.C.; Deglaire, A.; Ménard, O.; Bellanger, A.; Rousseau, F.; Henry, G.; Dirson, E.; Carrière, F.; Dupont, D.; Bourlieu, C. Holder pasteurization impacts the proteolysis, lipolysis and disintegration of human milk under in vitro dynamic term newborn digestion. Food Res. Int. 2016, 88, 263–275. [Google Scholar] [CrossRef]
- Davidson, L.A.; Lonnerdal, B. Persistence of human milk proteins in the breast-fed infant. Acta Paediatr. Scand. 1987, 76, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Cundiff, J.K.; Maria, S.D.; McMahon, R.J.; Wickham, M.S.J.; Faulks, R.M.; Van Tol, E.A.F. Differential digestion of human milk proteins in a simulated stomach model. J. Proteome Res. 2014, 13, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Peterson, J.A.; Hamosh, M.; Scallan, C.D.; Ceriani, R.L.; Henderson, T.R.; Mehta, N.R.; Armand, M.; Hamosh, P. Milk fat globule glycoproteins in human milk and in gastric aspirates of mother’s milk-fed preterm infants. Pediatr. Res. 1998, 44, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Beverly, R.L.; Underwood, M.A.; Dallas, D.C. Peptidomics Analysis of Milk Protein-Derived Peptides Released over Time in the Preterm Infant Stomach. J. Proteome Res. 2019, 18, 912–922. [Google Scholar] [CrossRef]
- Wada, Y.; Phinney, B.S.; Weber, D.; Lönnerdal, B. In vivo digestomics of milk proteins in human milk and infant formula using a suckling rat pup model. Peptides 2017, 88, 18–31. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Normal Weight a (N = 9) | Overweight/Obese a (N = 9) | p-Value b |
|---|---|---|---|
| Age, years | 33.0 (5.5) | 32.0 (5.0) | 0.8642 |
| Colostrum collection, days postpartum | 4.0 (1.0) | 5.0 (1.0) | 0.1340 |
| Transitional milk collection, days postpartum | 9.5 (1.0) | 10.3 (1.3) | 0.1970 |
| Mature milk collection, days postpartum | 19.0 (3.0) | 19.0 (0.8) | 0.6445 |
| Gestational age, days | 278.0 (14.0) | 282.0 (10.8) | 0.2162 |
| BMI early pregnancy, kg m−2 | 21.45 (1.36) | 30.53 (7.88) | 0.0012 |
| BMI at delivery, kg m−2 | 25.95 (2.39) | 34.23 (3.64) | 0.0047 |
| Weight gain, kg | 12.0 (4.5) | 11.5 (4.5) | 0.7576 |
| Newborn weight, kg | 3.23 (0.40) | 3.44 (0.38) | 0.2343 |
| Gender distribution, % | |||
| Female | 43 | 50 | |
| Male | 57 | 50 |
| Protein Name | Accession Number | Peptides Identified in Two or More Subjects (N) | Mol. Weight, Parent Protein (kDa) | Sequence Length, Parent Protein | Score (Digest Matches) |
|---|---|---|---|---|---|
| β-casein | P05814 | 123 | 25.38 | 226 | 332.54 |
| αs1-casein | P47710 | 26 | 21.67 | 185 | 46.73 |
| κ-casein | P07498 | 11 | 20.31 | 182 | 36.41 |
| Lactotransferrin | P02788 | 121 | 78.18 | 710 | 103.31 |
| α-lactalbumin | P00709 | 61 | 16.23 | 142 | 277.99 |
| Bile Salt-activated lipase | P19835 | 19 | 79.32 | 753 | 30.53 |
| Immunoglobulin heavy constant alpha 2 | P01877 | 18 | 36.59 | 340 | 27.96 |
| Butyrophilin subfamily 1 member A1 | Q13410 | 23 | 58.96 | 526 | 28.9 |
| Polymericimmunoglobulin receptor | P01833 | 25 | 83.28 | 764 | 37.73 |
| Tenascin | P24821 | 59 | 240.85 | 2201 | 29.5 |
| Mucin-4 | Q99102 | 56 | 231.52 | 2169 | 38.42 |
| Xanthine dehydrogenase/oxidase | P47989 | 44 | 146.42 | 1333 | 23.27 |
| Receptor tyrosine-proteinkinase erbB-4 | Q15303 | 29 | 146.81 | 1308 | 28.3 |
| Osteopontin | P10451 | 19 | 35.42 | 314 | 24.96 |
| Cadherin-1 | P12830 | 12 | 97.46 | 882 | 28.02 |
| Clusterin | P10909 | 11 | 52.50 | 449 | 25.23 |
| Galectin-3-binding protein | Q08380 | 9 | 65.33 | 585 | 24.74 |
| Mucin-1 | P15941 | 8 | 122.10 | 1255 | 28.77 |
| Zinc-alpha-2-glycoprotein | P25311 | 4 | 34.26 | 298 | 20.67 |
| Plateletglycoprotein 4 | P16671 | 3 | 53.05 | 472 | 22.34 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Hernández, S.; Théron, L.; Jiménez-Barrios, P.; Olalla-Herrera, M.; Recio, I.; Miralles, B. Protein Profile and Simulated Digestive Behavior of Breast Milk from Overweight and Normal Weight Mothers. Foods 2021, 10, 887. https://doi.org/10.3390/foods10040887
Sánchez-Hernández S, Théron L, Jiménez-Barrios P, Olalla-Herrera M, Recio I, Miralles B. Protein Profile and Simulated Digestive Behavior of Breast Milk from Overweight and Normal Weight Mothers. Foods. 2021; 10(4):887. https://doi.org/10.3390/foods10040887
Chicago/Turabian StyleSánchez-Hernández, Silvia, Laëtitia Théron, Pablo Jiménez-Barrios, Manuel Olalla-Herrera, Isidra Recio, and Beatriz Miralles. 2021. "Protein Profile and Simulated Digestive Behavior of Breast Milk from Overweight and Normal Weight Mothers" Foods 10, no. 4: 887. https://doi.org/10.3390/foods10040887
APA StyleSánchez-Hernández, S., Théron, L., Jiménez-Barrios, P., Olalla-Herrera, M., Recio, I., & Miralles, B. (2021). Protein Profile and Simulated Digestive Behavior of Breast Milk from Overweight and Normal Weight Mothers. Foods, 10(4), 887. https://doi.org/10.3390/foods10040887