
In Vitro Antioxidant and Anti-Colon Cancer Activities of Sesamum indicum L. Leaf Extract and Its Major Component, Pedaliin

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Sesame Leaf Extract
2.3. Antioxidant Phytochemical Contents
2.4. Antioxidant Activity Assay
2.5. Cell Culture and General Condition for Treatment
2.6. Cell Viability Assay
2.7. Flow Cytometry
2.8. Trans-Well Invasion and Migration Assay
2.9. Scratch Would-Healing Migration Assay
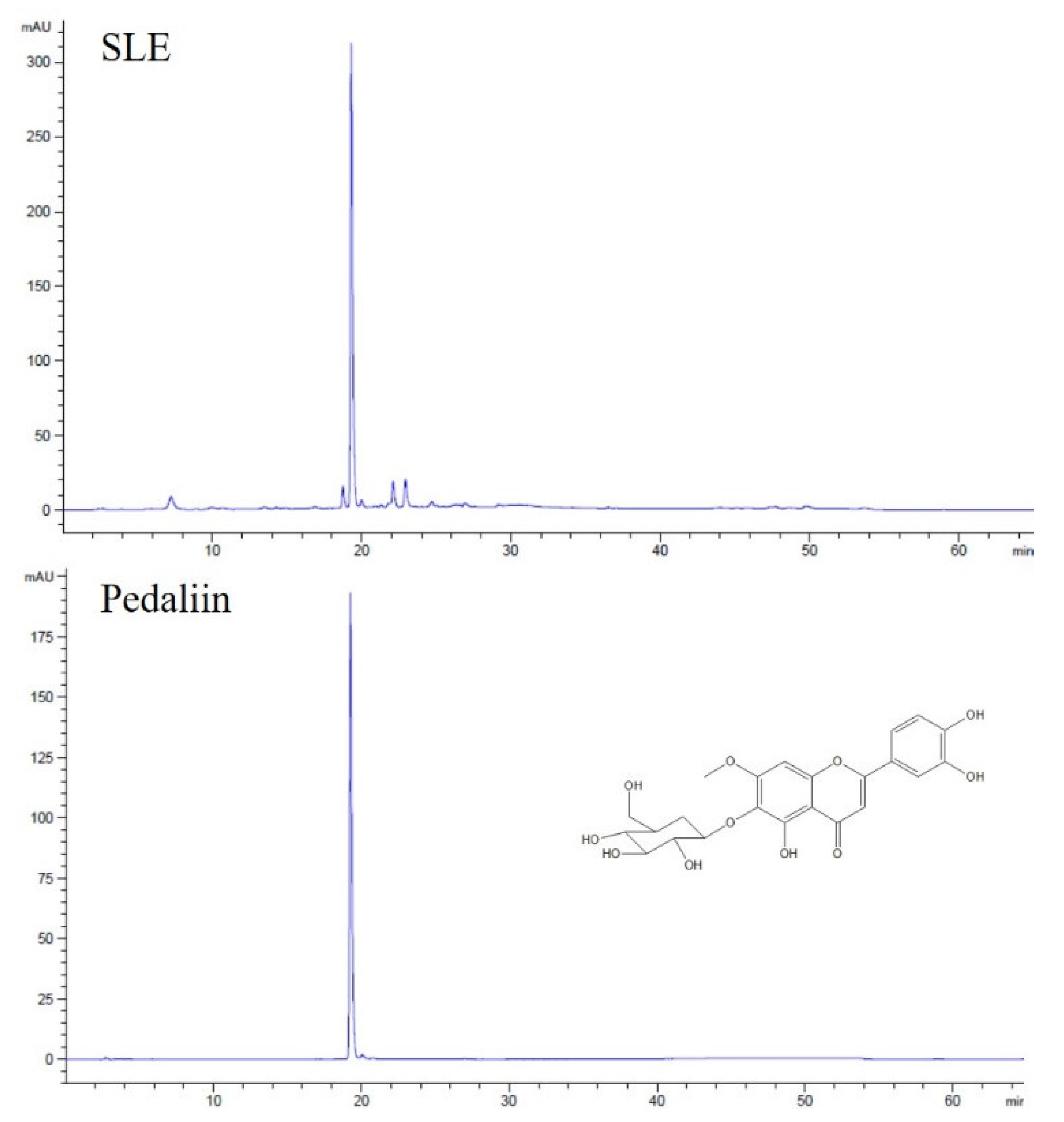
2.10. High-Performance Liquid Chromatography Anlaysis
2.11. Data Analyses
3. Results and Discussion
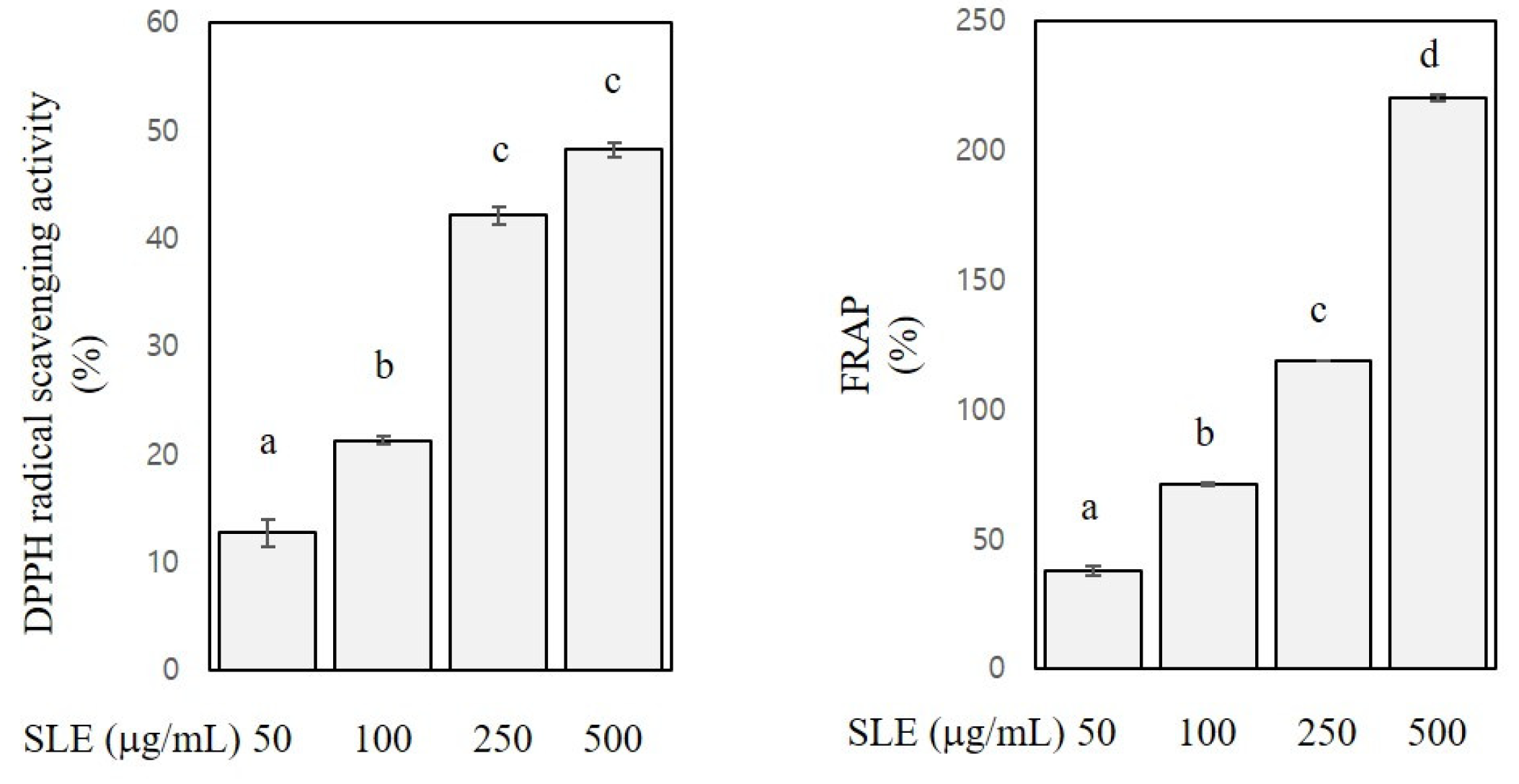
3.1. Antioxidant Contents and Activities of SLE
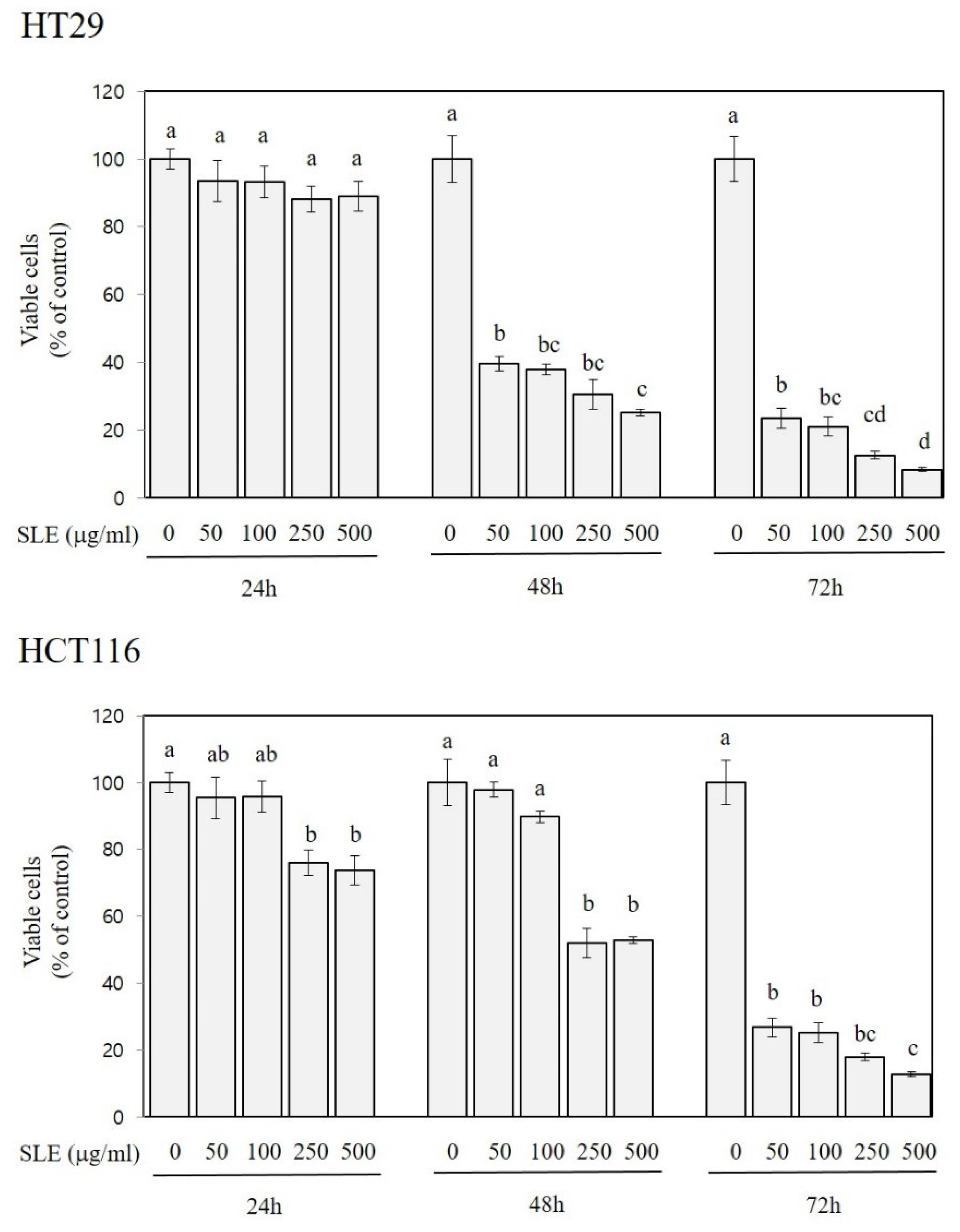
3.2. Growth-Inhibitory Activities of SLE in Human Colon Cancer Cells
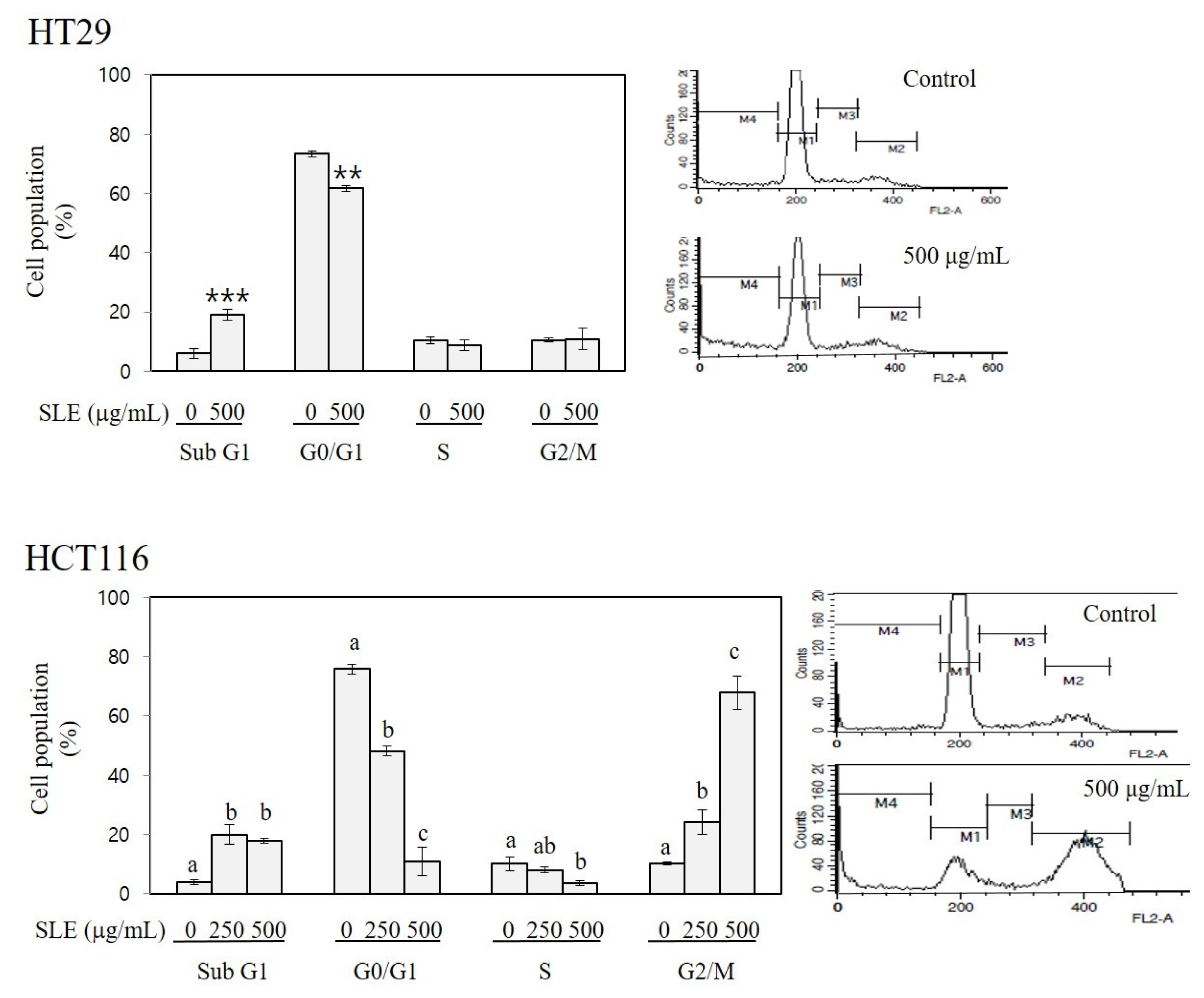
3.3. Apoptosis-Inducing and Cell Cycle-Arresting Activities of SLE in Human Colon Cancer Cells
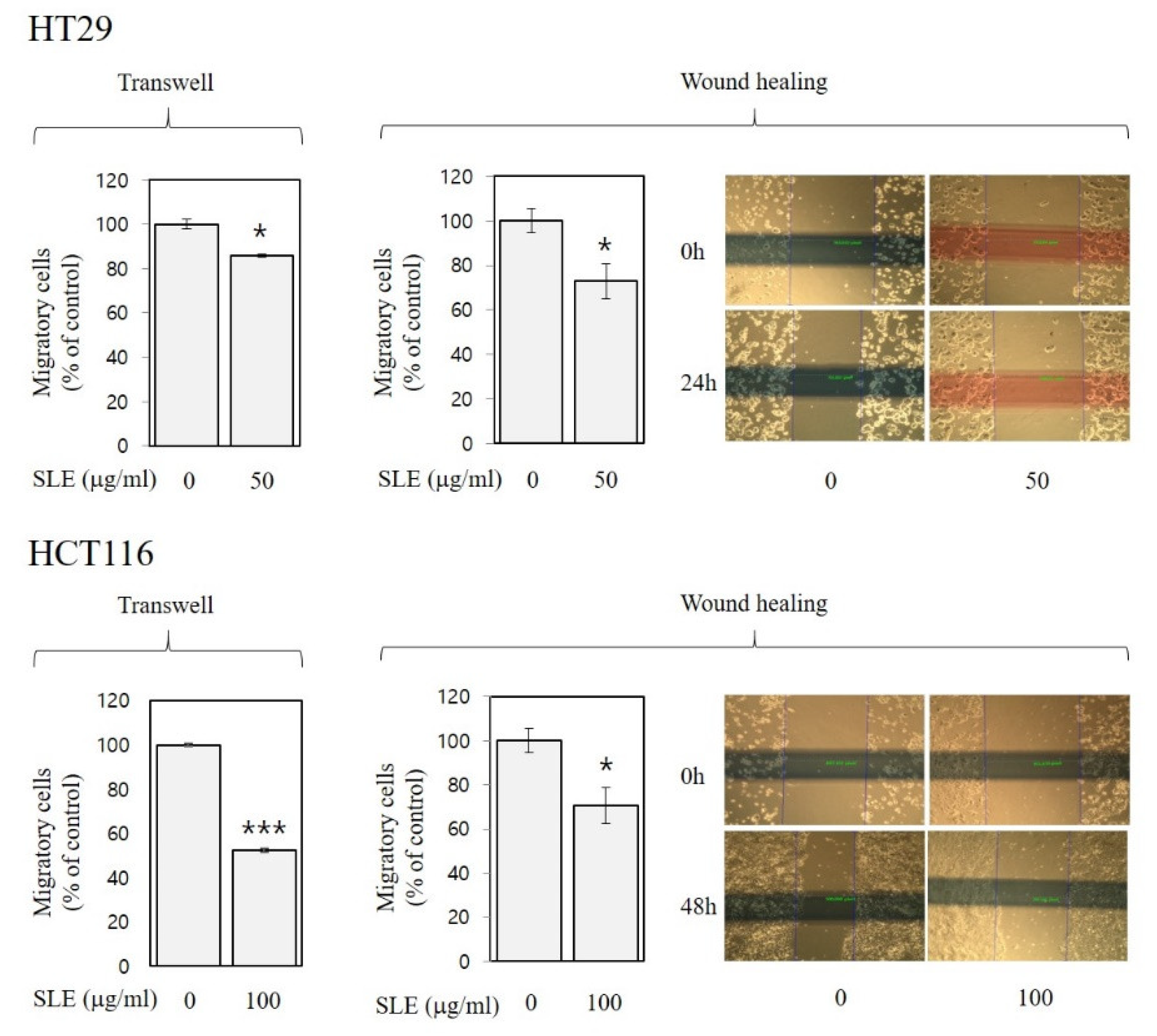
3.4. Inhibitory Activities of SLE against Invasion and Migration in Human Colon Cancer Cells
3.5. Pedaliin as a Major Constituent of SLE
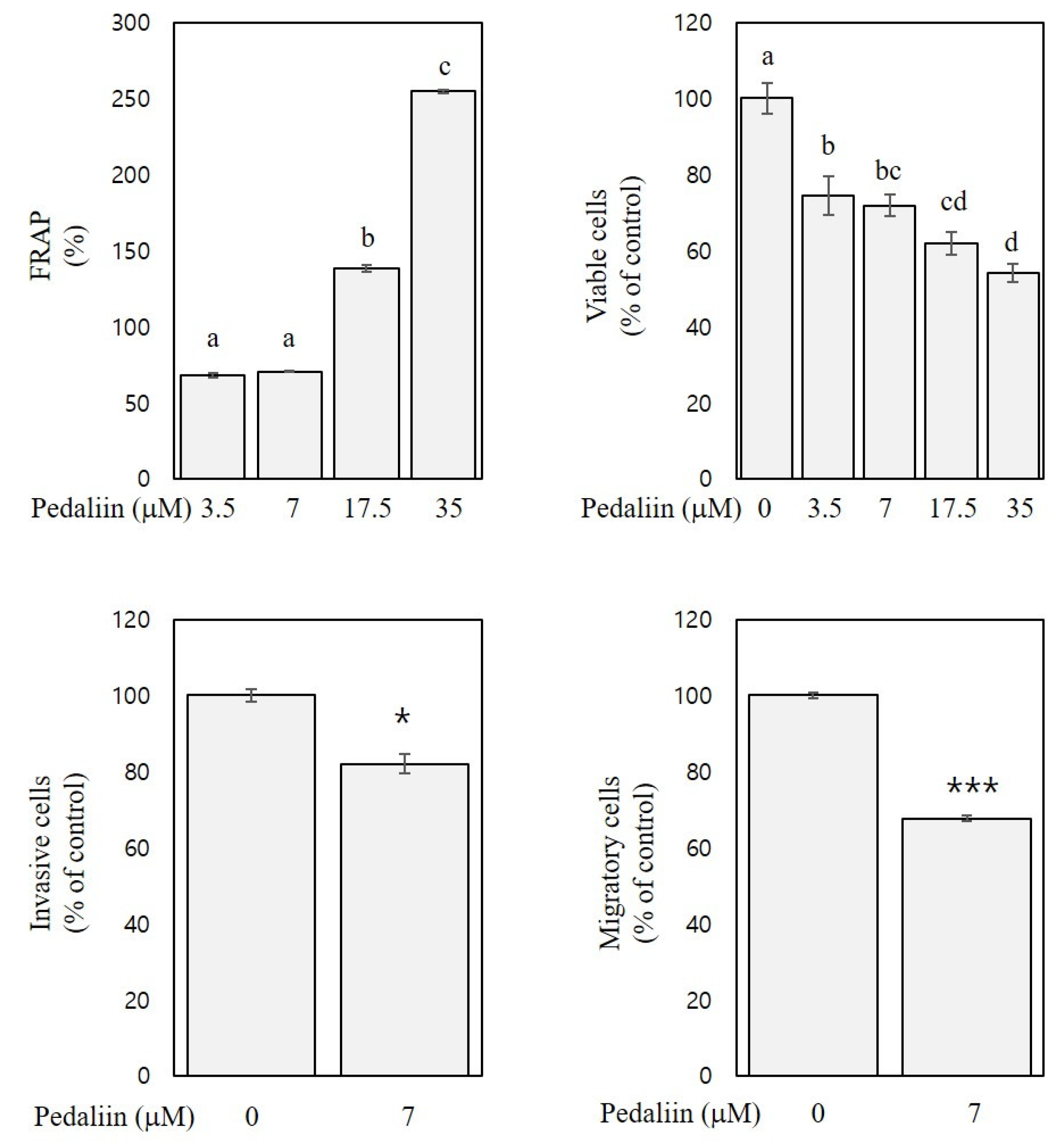
3.6. FRAP and In Vitro Anti-Colon Cancer Activities of Pedaliin
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, M.-S.; Aquino, L.B.B.; Barbaza, M.Y.U.; Hsieh, C.-L.; De Castro-Cruz, K.A.; Yang, L.-L.; Tsai, P.-W. Anti-Inflammatory and Anticancer Properties of Bioactive Compounds from Sesamum indicum L.—A Review. Molecules 2019, 24, 4426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andargie, M.; Vinas, M.; Rathgeb, A.; Möller, E.; Karlovsky, P. Lignans of Sesame (Sesamum indicum L.): A Comprehensive Review. Molecules 2021, 26, 883. [Google Scholar] [CrossRef] [PubMed]
- Namiki, M. Nutraceutical Functions of Sesame: A Review. Crit. Rev. Food Sci. Nutr. 2007, 47, 651–673. [Google Scholar] [CrossRef] [PubMed]
- Fuji, Y.; Uchida, A.; Fukahori, K.; Chino, M.; Ohtsuki, T.; Matsufuji, H. Chemical characterization and biological activity in young sesame leaves (Sesamum indicum L.) and changes in iridoid and polyphenol content at different growth stages. PLoS ONE 2018, 13, e0194449. [Google Scholar] [CrossRef]
- Kubmarawa, D.; Andenyang, I.; Magomya, A. Amino acid profile of two non-conventional leafy vegetables, Sesamum indicum and Balanites aegyptiaca. Afr. J. Biotechnol. 2008, 7, 3502–3504. [Google Scholar]
- Fuji, Y.; Ohtsuki, T.; Matsufuji, H. Accumulation and Subcellular Localization of Acteoside in Sesame Plants (Sesamum indicum L.). ACS Omega 2018, 3, 17287–17294. [Google Scholar] [CrossRef] [Green Version]
- Ohtsuki, T.; Matsuoka, K.; Fuji, Y.; Nishizaki, Y.; Masumoto, N.; Sugimoto, N.; Sato, K.; Matsufuji, H. Development of an HPLC method with relative molar sensitivity based on 1H-qNMR to determine acteoside and pedaliin in dried sesame leaf powders and processed foods. PLoS ONE 2020, 15, e0243175. [Google Scholar] [CrossRef]
- Dat, N.T.; Dang, N.H.; Thanh le, N. New flavonoid and pentacyclic triterpene from Sesamum indicum leaves. Nat. Prod. Res. 2016, 30, 311–315. [Google Scholar] [CrossRef]
- Morita, N. Studies on the Flavonoid of Sesame Leaves. I. On the Structure of the Glycoside, Pedaliin. Chem. Pharm. Bull. 1960, 8, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Okwuosa, C.N.; Okoi-Ewa, R.; Achukwu, P.U.; Onuba, A.C.; Azubuike, N.C. Gastro-protective effect of crude hexane leaf extract of Sesamum indicum in rabbits. Niger. J. Physiol. Sci. 2011, 26, 49–54. [Google Scholar]
- Ribeiro, S.B.; Pagnocca, F.C.; Victor, S.R.; Bueno, O.C.; Hebling, M.J.; Bacci, M., Jr.; Silva, O.A.; Fernandes, J.B.; Vieira, P.C.; Silva, M.F. Activity of sesame leaf extracts against the symbiotic fungus of Atta sexdens L. An. Soc. Entomol. Bras. 1998, 27, 421–426. [Google Scholar] [CrossRef]
- Kwak, Y.; Ki, S.; Noh, E.K.; Shin, H.N.; Han, Y.-J.; Lee, Y.; Ju, J. Comparison of antioxidant and anti-proliferative activities of perilla (Perilla frutescens Britton) and sesame (Seasamum indicum L.) leaf extracts. Korean J. Food Cookery Sci. 2013, 29, 241–248. [Google Scholar]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Klaunig, J.E.; Wang, Z. Oxidative stress in carcinogenesis. Curr. Opin. Toxicol. 2018, 7, 116–121. [Google Scholar] [CrossRef]
- Fernández-Villa, T.; Álvarez-Álvarez, L.; Rubín-García, M.; Obón-Santacana, M.; Moreno, V. The role of dietary patterns in colorectal cancer: A 2019 update. Expert Rev. Gastroenterol. Hepatol. 2020, 14, 1–10. [Google Scholar] [CrossRef]
- Csepregi, K.; Kocsis, M.; Hideg, E. On the spectrophotometric determination of total phenolic and flavonoid contents. Acta Biol. Hung. 2013, 64, 500–509. [Google Scholar] [CrossRef]
- Margraf, T.; Karnopp, A.R.; Rosso, N.D.; Granato, D. Comparison between Folin-Ciocalteu and Prussian Blue Assays to Estimate The Total Phenolic Content of Juices and Teas Using 96-Well Microplates. J. Food Sci. 2015, 80, C2397–C2403. [Google Scholar] [CrossRef]
- Seo, J.; Lee, J.; Yang, H.Y.; Ju, J. Antirrhinum majus L. flower extract inhibits cell growth and metastatic properties in human colon and lung cancer cell lines. Food Sci. Nutr. 2020, 8, 6259–6268. [Google Scholar] [CrossRef] [PubMed]
- Scolnik, P.A.; Zannoni, D.; Marrs, B.L. Spectral and functional comparisons between the carotenoids of the two antenna complexes of Rhodopseudomonas capsulata. Biochim. Biophys. Acta Bioenerg. 1980, 593, 230–240. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Payet, B.; Sing, A.S.C.; Smadja, J. Assessment of Antioxidant Activity of Cane Brown Sugars by ABTS and DPPH Radical Scavenging Assays: Determination of Their Polyphenolic and Volatile Constituents. J. Agric. Food Chem. 2005, 53, 10074–10079. [Google Scholar] [CrossRef]
- Visavadiya, N.P.; Soni, B.; Dalwadi, N. Free radical scavenging and antiatherogenic activities of Sesamum indicum seed extracts in chemical and biological model systems. Food Chem. Toxicol. 2009, 47, 2507–2515. [Google Scholar] [CrossRef]
- Gul, M.Z.; Bhakshu, L.M.; Ahmad, F.; Kondapi, A.K.; Qureshi, I.A.; Ghazi, I.A. Evaluation of Abelmoschus moschatus extracts for antioxidant, free radical scavenging, antimicrobial and antiproliferative activities using in vitro assays. BMC Complement. Altern. Med. 2011, 11, 64. [Google Scholar] [CrossRef] [Green Version]
- Yahia, Y.; Benabderrahim, M.A.; Tlili, N.; Bagues, M.; Nagaz, K. Bioactive compounds, antioxidant and antimicrobial activities of extracts from different plant parts of two Ziziphus Mill. species. PLoS ONE 2020, 15, e0232599. [Google Scholar] [CrossRef] [PubMed]
- Khanam, U.K.S.; Oba, S.; Yanase, E.; Murakami, Y. Phenolic acids, flavonoids and total antioxidant capacity of selected leafy vegetables. J. Funct. Foods 2012, 4, 979–987. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Gan, R.-Y.; Li, S.; Zhou, Y.; Li, A.-N.; Xu, D.-P.; Li, H.-B. Antioxidant Phytochemicals for the Prevention and Treatment of Chronic Diseases. Molecules 2015, 20, 21138–21156. [Google Scholar] [CrossRef] [PubMed]
- Evan, G.I.; Vousden, K.H. Proliferation, cell cycle and apoptosis in cancer. Nat. Cell Biol. 2001, 411, 342–348. [Google Scholar] [CrossRef]
- Chen, Y.; Zhu, Z.; Guo, Q.; Zhang, L.; Zhang, X. Variation in concentrations of major bioactive compounds in Prunella vulgaris L. related to plant parts and phenological stages. Biol. Res. 2012, 45, 171–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sellami, I.H.; Maamouri, E.; Chahed, T.; Wannes, W.A.; Kchouk, M.E.; Marzouk, B. Effect of growth stage on the content and composition of the essential oil and phenolic fraction of sweet marjoram (Origanum majorana L.). Ind. Crop. Prod. 2009, 30, 395–402. [Google Scholar] [CrossRef]
- Liu, R.H. Potential Synergy of Phytochemicals in Cancer Prevention: Mechanism of Action. J. Nutr. 2004, 134, 3479S–3485S. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytochemicals | Contents |
|---|---|
| Extraction yield | 11.2 1 |
| Total polyphenols | 36.5 ± 1.1 2,3 |
| Total flavonoids | 129.2 ± 20.9 2,4 |
| Carotenoids | 537.7 ± 67.6 2,5 |
| Concentration Range Tested (μg/mL) | Linear Regression Data | LOD 1 (μg/mL) | LOQ 1 (μg/mL) | |
|---|---|---|---|---|
| Calibration Curve 2 | Correlation Coefficient (R2) | |||
| 1–500 | Y = 38.919X + 98.892 | 0.9997 | 0.040 | 0.135 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.; Yang, H.Y.; Lee, H.J.; Ju, J. In Vitro Antioxidant and Anti-Colon Cancer Activities of Sesamum indicum L. Leaf Extract and Its Major Component, Pedaliin. Foods 2021, 10, 1216. https://doi.org/10.3390/foods10061216
Kim S, Yang HY, Lee HJ, Ju J. In Vitro Antioxidant and Anti-Colon Cancer Activities of Sesamum indicum L. Leaf Extract and Its Major Component, Pedaliin. Foods. 2021; 10(6):1216. https://doi.org/10.3390/foods10061216
Chicago/Turabian StyleKim, Seoyun, Hyi Young Yang, Hwa Jin Lee, and Jihyeung Ju. 2021. "In Vitro Antioxidant and Anti-Colon Cancer Activities of Sesamum indicum L. Leaf Extract and Its Major Component, Pedaliin" Foods 10, no. 6: 1216. https://doi.org/10.3390/foods10061216
APA StyleKim, S., Yang, H. Y., Lee, H. J., & Ju, J. (2021). In Vitro Antioxidant and Anti-Colon Cancer Activities of Sesamum indicum L. Leaf Extract and Its Major Component, Pedaliin. Foods, 10(6), 1216. https://doi.org/10.3390/foods10061216