Human Milk Oligosaccharides (HMOs) and Infant Microbiota: A Scoping Review


 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Methodologies
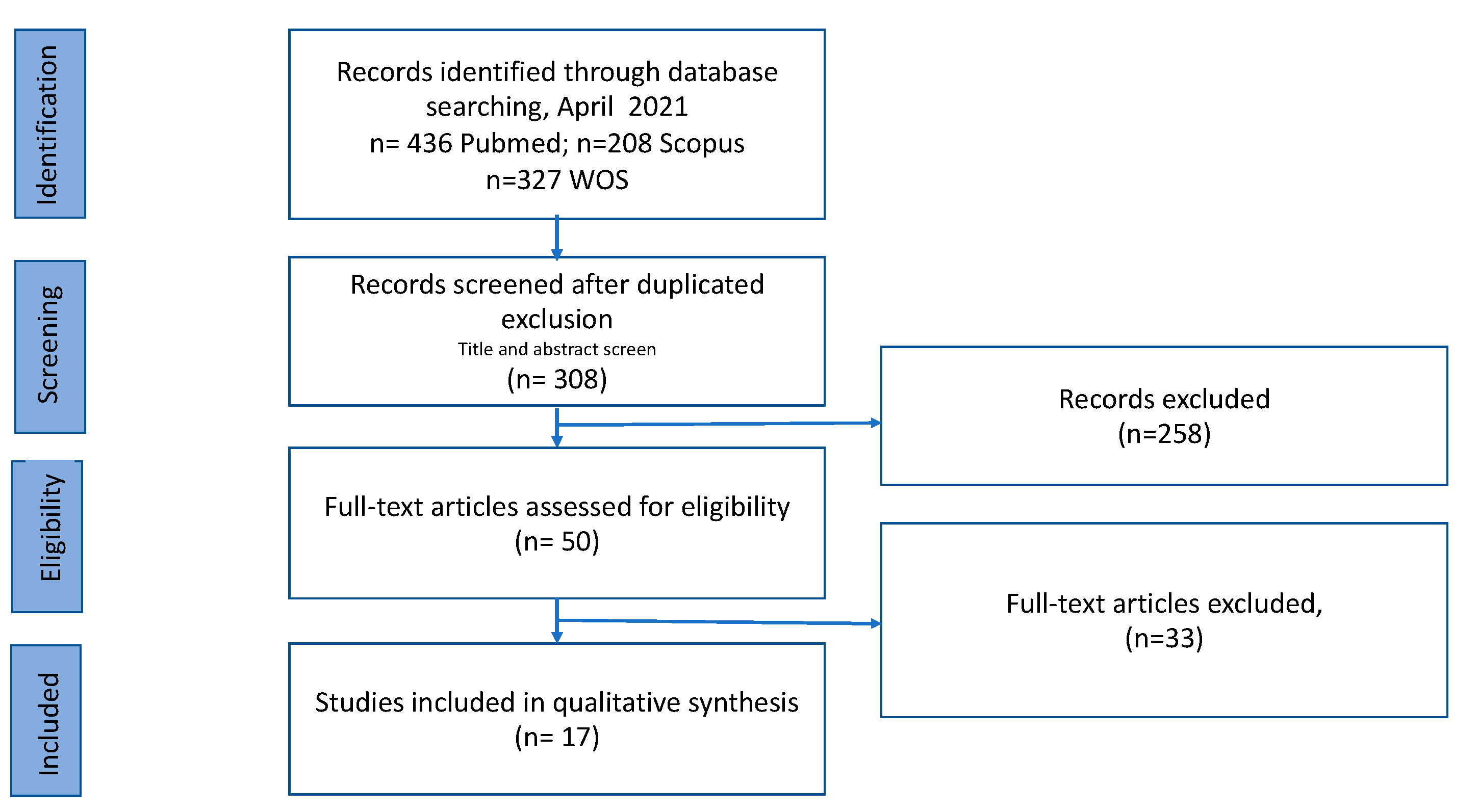
2.1. Search Strategy and Inclusion Criteria
2.2. Article Screening and Data Abstraction
3. Results
3.1. Synthesis
3.2. Analysis of Methodologies
4. Discussion
4.1. What Impact Does MSeS Have on the Infant Gut Microbiota?
4.1.1. Considering the Mode of Birth
4.1.2. Regardless of the Mode of Delivery
4.2. What Impact Does the HMO Profile Have on the Infant Gut Microbiota?
4.3. Can Specific HMOs Be Added to Formula to Modulate the Infant Microbiota?
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Collective, G.B. UNICEF Nurturing the Health and Wealth of Nations: The Investment Case for Breastfeeding; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Chouraqui, J.-P. Does the contribution of human milk oligosaccharides to the beneficial effects of breast milk allow us to hope for an improvement in infant formulas? Crit. Rev. Food Sci. Nutr. 2020, 1–12. [Google Scholar] [CrossRef]
- Bode, L. The functional biology of human milk oligosaccharides. Early Hum. Dev. 2015, 91, 619–622. [Google Scholar] [CrossRef]
- Balogh, R.; Jankovics, P.; Béni, S. Qualitative and quantitative analysis of N-acetyllactosamine and lacto-N-biose, the two major building blocks of human milk oligosaccharides in human milk samples by high-performance liquid chromatography–tandem mass spectrometry using a porous graphitic. J. Chromatogr. A 2015, 1422, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Vandenplas, Y.; Berger, B.; Carnielli, V.P.; Ksiazyk, J.; Lagström, H.; Sanchez Luna, M.; Migacheva, N.; Mosselmans, J.-M.; Picaud, J.-C.; Possner, M. Human milk oligosaccharides: 2′-fucosyllactose (2′-FL) and lacto-N-neotetraose (LNnT) in infant formula. Nutrients 2018, 10, 1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stahl, B.; Thurl, S.; Henker, J.; Siegel, M.; Finke, B.; Sawatzki, G. Detection of four human milk groups with respect to Lewis-bloodgroup-dependent oligosaccharides by serologic and chromatographic analysis. Adv. Exp. Med. Biol. 2001, 299–306. [Google Scholar]
- Azad, M.B.; Robertson, B.; Atakora, F.; Becker, A.B.; Subbarao, P.; Moraes, T.J.; Mandhane, P.J.; Turvey, S.E.; Lefebvre, D.L.; Sears, M.R. Human milk oligosaccharide concentrations are associated with multiple fixed and modifiable maternal characteristics, environmental factors, and feeding practices. J. Nutr. 2018, 148, 1733–1742. [Google Scholar] [CrossRef]
- Garrido, D.; Ruiz-Moyano, S.; Kirmiz, N.; Davis, J.C.; Totten, S.M.; Lemay, D.G.; Ugalde, J.A.; German, J.B.; Lebrilla, C.B.; Mills, D.A. A novel gene cluster allows preferential utilization of fucosylated milk oligosaccharides in Bifidobacterium longum subsp. longum SC596. Sci. Rep. 2016, 6, 35045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickramasinghe, S.; Pacheco, A.R.; Lemay, D.G.; Mills, D.A. Bifidobacteria grown on human milk oligosaccharides downregulate the expression of inflammation-related genes in Caco-2 cells. BMC Microbiol. 2015, 15, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newburg, D.S.; Ruiz-Palacios, G.M.; Morrow, A.L. Human milk glycans protect infants against enteric pathogens. Annu. Rev. Nutr. 2005, 25, 37–58. [Google Scholar] [CrossRef]
- Xiao, L.; van De Worp, W.R.P.H.; Stassen, R.; van Maastrigt, C.; Kettelarij, N.; Stahl, B.; Blijenberg, B.; Overbeek, S.A.; Folkerts, G.; Garssen, J. Human milk oligosaccharides promote immune tolerance via direct interactions with human dendritic cells. Eur. J. Immunol. 2019, 49, 1001–1014. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.E.; Autran, C.A.; Szyszka, A.; Escajadillo, T.; Huang, M.; Godula, K.; Prudden, A.R.; Boons, G.-J.; Lewis, A.L.; Doran, K.S. Human milk oligosaccharides inhibit growth of group B Streptococcus. J. Biol. Chem. 2017, 292, 11243–11249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tricco, A.C.; Lillie, E.; Zarin, W.; O’Brien, K.K.; Colquhoun, H.; Levac, D.; Moher, D.; Peters, M.D.J.; Horsley, T.; Weeks, L. PRISMA extension for scoping reviews (PRISMA-ScR): Checklist and explanation. Ann. Intern. Med. 2018, 169, 467–473. [Google Scholar] [CrossRef] [Green Version]
- Arksey, H.; O’Malley, L. Scoping studies: Towards a methodological framework. Int. J. Soc. Res. Methodol. 2005, 8, 19–32. [Google Scholar] [CrossRef] [Green Version]
- Tonon, K.M.; Morais, T.B.; Taddei, C.R.; Araújo-Filho, H.B.; Abrão, A.C.F.V.; Miranda, A.; de Morais, M.B. Gut microbiota comparison of vaginally and cesarean born infants exclusively breastfed by mothers secreting α1–2 fucosylated oligosaccharides in breast milk. PLoS ONE 2021, 16, e0246839. [Google Scholar] [CrossRef]
- Masi, A.C.; Embleton, N.D.; Lamb, C.A.; Young, G.; Granger, C.L.; Najera, J.; Smith, D.P.; Hoffman, K.L.; Petrosino, J.F.; Bode, L. Human milk oligosaccharide DSLNT and gut microbiome in preterm infants predicts necrotising enterocolitis. Gut 2020. [Google Scholar] [CrossRef] [PubMed]
- Borewicz, K.; Gu, F.; Saccenti, E.; Hechler, C.; Beijers, R.; de Weerth, C.; van Leeuwen, S.S.; Schols, H.A.; Smidt, H. The association between breastmilk oligosaccharides and faecal microbiota in healthy breastfed infants at two, six, and twelve weeks of age. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Berger, B.; Porta, N.; Foata, F.; Grathwohl, D.; Delley, M.; Moine, D.; Charpagne, A.; Siegwald, L.; Descombes, P.; Alliet, P. Linking human milk oligosaccharides, infant fecal community types, and later risk to require antibiotics. MBio 2020, 11, e03196-19. [Google Scholar] [CrossRef] [Green Version]
- Quin, C.; Vicaretti, S.D.; Mohtarudin, N.A.; Garner, A.M.; Vollman, D.M.; Gibson, D.L.; Zandberg, W.F. Influence of sulfonated and diet-derived human milk oligosaccharides on the infant microbiome and immune markers. J. Biol. Chem. 2020, 295, 4035–4048. [Google Scholar] [CrossRef]
- Borewicz, K.; Gu, F.; Saccenti, E.; Arts, I.C.W.; Penders, J.; Thijs, C.; van Leeuwen, S.S.; Lindner, C.; Nauta, A.; van Leusen, E. Correlating Infant Fecal Microbiota Composition and Human Milk Oligosaccharide Consumption by Microbiota of 1-Month-Old Breastfed Infants. Mol. Nutr. Food Res. 2019, 63, 1801214. [Google Scholar] [CrossRef]
- Paganini, D.; Uyoga, M.A.; Kortman, G.A.M.; Boekhorst, J.; Schneeberger, S.; Karanja, S.; Hennet, T.; Zimmermann, M.B. Maternal human milk oligosaccharide profile modulates the impact of an intervention with iron and galacto-oligosaccharides in Kenyan infants. Nutrients 2019, 11, 2596. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Tao, J.; Zhou, J.; Fan, Q.; Liu, M.; Hu, Y.; Xu, Y.; Zhang, L.; Yuan, J.; Li, W. Fucosylated human milk oligosaccharides and N-glycans in the milk of Chinese mothers regulate the gut microbiome of their breast-fed infants during different lactation stages. MSystems 2018, 3, e00206-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korpela, K.; Salonen, A.; Hickman, B.; Kunz, C.; Sprenger, N.; Kukkonen, K.; Savilahti, E.; Kuitunen, M.; de Vos, W.M. Fucosylated oligosaccharides in mother’s milk alleviate the effects of caesarean birth on infant gut microbiota. Sci. Rep. 2018, 8, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, J.C.C.; Lewis, Z.T.; Krishnan, S.; Bernstein, R.M.; Moore, S.E.; Prentice, A.M.; Mills, D.A.; Lebrilla, C.B.; Zivkovic, A.M. Growth and morbidity of Gambian infants are influenced by maternal milk oligosaccharides and infant gut microbiota. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef]
- Underwood, M.A.; Davis, J.C.C.; Kalanetra, K.M.; Gehlot, S.; Patole, S.; Tancredi, D.J.; Mills, D.A.; Lebrilla, C.B.; Simmer, K. Digestion of human milk oligosaccharides by Bifidobacterium breve in the premature infant. J. Pediatr. Gastroenterol. Nutr. 2017, 65, 449. [Google Scholar] [CrossRef]
- Matsuki, T.; Tajima, S.; Hara, T.; Yahagi, K.; Ogawa, E.; Kodama, H. Infant formula with galacto-oligosaccharides (OM55N) stimulates the growth of indigenous bifidobacteria in healthy term infants. Benef. Microbes 2016, 7, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Smith-Brown, P.; Morrison, M.; Krause, L.; Davies, P.S.W. Mothers secretor status affects development of childrens microbiota composition and function: A pilot study. PLoS ONE 2016, 11, e0161211. [Google Scholar] [CrossRef] [PubMed]
- Lewis, Z.T.; Totten, S.M.; Smilowitz, J.T.; Popovic, M.; Parker, E.; Lemay, D.G.; Van Tassell, M.L.; Miller, M.J.; Jin, Y.-S.; German, J.B. Maternal fucosyltransferase 2 status affects the gut bifidobacterial communities of breastfed infants. Microbiome 2015, 3, 1–21. [Google Scholar] [CrossRef] [Green Version]
- De Leoz, M.L.A.; Kalanetra, K.M.; Bokulich, N.A.; Strum, J.S.; Underwood, M.A.; German, J.B.; Mills, D.A.; Lebrilla, C.B. Human milk glycomics and gut microbial genomics in infant feces show a correlation between human milk oligosaccharides and gut microbiota: A proof-of-concept study. J. Proteome Res. 2015, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Li, M.; Wu, S.; Lebrilla, C.B.; Chapkin, R.S.; Ivanov, I.; Donovan, S.M. Fecal microbiota composition of breast-fed infants is correlated with human milk oligosaccharides consumed. J. Pediatr. Gastroenterol. Nutr. 2015, 60, 825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coppa, G.V.; Gabrielli, O.; Zampini, L.; Galeazzi, T.; Ficcadenti, A.; Padella, L.; Santoro, L.; Soldi, S.; Carlucci, A.; Bertino, E. Oligosaccharides in 4 different milk groups, Bifidobacteria, and Ruminococcus obeum. J. Pediatr. Gastroenterol. Nutr. 2011, 53, 80–87. [Google Scholar] [CrossRef]
- Marizzoni, M.; Gurry, T.; Provasi, S.; Greub, G.; Lopizzo, N.; Ribaldi, F.; Festari, C.; Mazzelli, M.; Mombelli, E.; Salvatore, M. Comparison of bioinformatics pipelines and operating systems for the analyses of 16S rRNA gene amplicon sequences in human fecal samples. Front. Microbiol. 2020, 11, 1262. [Google Scholar] [CrossRef]
- Leyva, L.L.; Brereton, N.J.B.; Koski, K.G. Emerging frontiers in human milk microbiome research and suggested primers for 16S rRNA gene analysis. Comput. Struct. Biotechnol. J. 2020, 19, 121–133. [Google Scholar] [CrossRef]
- Walker, A.W.; Martin, J.C.; Scott, P.; Parkhill, J.; Flint, H.J.; Scott, K.P. 16S rRNA gene-based profiling of the human infant gut microbiota is strongly influenced by sample processing and PCR primer choice. Microbiome 2015, 3, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Velásquez-Mejía, E.P.; de la Cuesta-Zuluaga, J.; Escobar, J.S. Impact of DNA extraction, sample dilution, and reagent contamination on 16S rRNA gene sequencing of human feces. Appl. Microbiol. Biotechnol. 2018, 102, 403–411. [Google Scholar] [CrossRef]
- Mancabelli, L.; Milani, C.; Lugli, G.A.; Fontana, F.; Turroni, F.; van Sinderen, D.; Ventura, M. The impact of primer design on amplicon-based metagenomic profiling accuracy: Detailed insights into bifidobacterial community structure. Microorganisms 2020, 8, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlaeppi, K.; Ronchi, F.; Leib, S.L.; Erb, M.; Ramette, A. Evaluation of primer pairs for microbiome profiling from soils to humans within the One Health framework. Mol. Ecol. Resour. 2020, 20, 1558–1571. [Google Scholar]
- Bukin, Y.S.; Galachyants, Y.P.; Morozov, I.V.; Bukin, S.V.; Zakharenko, A.S.; Zemskaya, T.I. The effect of 16S rRNA region choice on bacterial community metabarcoding results. Sci. Data 2019, 6, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirichoat, A.; Sankuntaw, N.; Engchanil, C.; Buppasiri, P.; Faksri, K.; Namwat, W.; Chantratita, W.; Lulitanond, V. Comparison of different hypervariable regions of 16S rRNA for taxonomic profiling of vaginal microbiota using next-generation sequencing. Arch. Microbiol. 2021, 203, 1159–1166. [Google Scholar] [CrossRef]
- Bode, L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology 2012, 22, 1147–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Muñoz, M.E.; Arrieta, M.-C.; Ramer-Tait, A.E.; Walter, J. A critical assessment of the “sterile womb” and “in utero colonization” hypotheses: Implications for research on the pioneer infant microbiome. Microbiome 2017, 5, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Vandenplas, Y.; Carnielli, V.P.; Ksiazyk, J.; Luna, M.S.; Migacheva, N.; Mosselmans, J.M.; Picaud, J.C.; Possner, M.; Singhal, A.; Wabitsch, M. Factors affecting early-life intestinal microbiota development. Nutrition 2020, 78, 110812. [Google Scholar] [CrossRef] [PubMed]
- Moubareck, C.A. Human Milk Microbiota and Oligosaccharides: A Glimpse into Benefits, Diversity, and Correlations. Nutrients 2021, 13, 1123. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.B.; Konya, T.; Maughan, H.; Guttman, D.S.; Field, C.J.; Chari, R.S.; Sears, M.R.; Becker, A.B.; Scott, J.A.; Kozyrskyj, A.L. Gut microbiota of healthy Canadian infants: Profiles by mode of delivery and infant diet at 4 months. Cmaj 2013, 185, 385–394. [Google Scholar] [CrossRef] [Green Version]
- Wiciński, M.; Sawicka, E.; Gębalski, J.; Kubiak, K.; Malinowski, B. Human milk oligosaccharides: Health benefits, potential applications in infant formulas, and pharmacology. Nutrients 2020, 12, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGuire, M.K.; Meehan, C.L.; McGuire, M.A.; Williams, J.E.; Foster, J.; Sellen, D.W.; Kamau-Mbuthia, E.W.; Kamundia, E.W.; Mbugua, S.; Moore, S.E. What’s normal? Oligosaccharide concentrations and profiles in milk produced by healthy women vary geographically. Am. J. Clin. Nutr. 2017, 105, 1086–1100. [Google Scholar] [CrossRef]
- Biddulph, C.; Holmes, M.; Kuballa, A.; Davies, P.S.W.; Koorts, P.; Carter, R.J.; Maher, J. Human Milk Oligosaccharide Profiles and Associations with Maternal Nutritional Factors: A Scoping Review. Nutrients 2021, 13, 965. [Google Scholar] [CrossRef]
- Kunz, C.; Meyer, C.; Collado, M.C.; Geiger, L.; García-Mantrana, I.; Bertua-Ríos, B.; Martínez-Costa, C.; Borsch, C.; Rudloff, S. Influence of gestational age, secretor, and lewis blood group status on the oligosaccharide content of human milk. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.J.; Ajami, N.J.; O’Brien, J.L.; Hutchinson, D.S.; Smith, D.P.; Wong, M.C.; Ross, M.C.; Lloyd, R.E.; Doddapaneni, H.; Metcalf, G.A. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature 2018, 562, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Ottman, N.A. Host Immunostimulation and Substrate Utilization of the Gut Symbiont Akkermansia Muciniphila. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2015. [Google Scholar]
- Derrien, M.; Belzer, C.; de Vos, W.M. Akkermansia muciniphila and its role in regulating host functions. Microb. Pathog. 2017, 106, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Huttenhower, C.; Gevers, D.; Knight, R.; Abubucker, S.; Badger, J.H.; Chinwalla, A.T.; Creasy, H.H.; Earl, A.M.; FitzGerald, M.G.; Fulton, R.S. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207. [Google Scholar]
- Xu, G.; Davis, J.C.C.; Goonatilleke, E.; Smilowitz, J.T.; German, J.B.; Lebrilla, C.B. Absolute quantitation of human milk oligosaccharides reveals phenotypic variations during lactation. J. Nutr. 2017, 147, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Garrido, D.; Kim, J.H.; German, J.B.; Raybould, H.E.; Mills, D.A. Oligosaccharide binding proteins from Bifidobacterium longum subsp. infantis reveal a preference for host glycans. PLoS ONE 2011, 6, e17315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sela, D.A.; Garrido, D.; Lerno, L.; Wu, S.; Tan, K.; Eom, H.-J.; Joachimiak, A.; Lebrilla, C.B.; Mills, D.A. Bifidobacterium longum subsp. infantis ATCC 15697 α-fucosidases are active on fucosylated human milk oligosaccharides. Appl. Environ. Microbiol. 2012, 78, 795–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sela, D.A.; Chapman, J.; Adeuya, A.; Kim, J.H.; Chen, F.; Whitehead, T.R.; Lapidus, A.; Rokhsar, D.S.; Lebrilla, C.B.; German, J.B. The genome sequence of Bifidobacterium longum subsp. infantis reveals adaptations for milk utilization within the infant microbiome. Proc. Natl. Acad. Sci. USA 2008, 105, 18964–18969. [Google Scholar] [CrossRef] [Green Version]
- LoCascio, R.G.; Ninonuevo, M.R.; Freeman, S.L.; Sela, D.A.; Grimm, R.; Lebrilla, C.B.; Mills, D.A.; German, J.B. Glycoprofiling of bifidobacterial consumption of human milk oligosaccharides demonstrates strain specific, preferential consumption of small chain glycans secreted in early human lactation. J. Agric. Food Chem. 2007, 55, 8914–8919. [Google Scholar] [CrossRef]
- Thurl, S.; Munzert, M.; Henker, J.; Boehm, G.; Müller-Werner, B.; Jelinek, J.; Stahl, B. Variation of human milk oligosaccharides in relation to milk groups and lactational periods. Br. J. Nutr. 2010, 104, 1261–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R. Linking long-term dietary patterns with gut microbial enterotypes. Science (80-.) 2011, 334, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Holmes, I.; Harris, K.; Quince, C. Dirichlet multinomial mixtures: Generative models for microbial metagenomics. PLoS ONE 2012, 7, e30126. [Google Scholar] [CrossRef] [Green Version]
- Simeoni, U.; Berger, B.; Junick, J.; Blaut, M.; Pecquet, S.; Rezzonico, E.; Grathwohl, D.; Sprenger, N.; Brüssow, H.; Team, S. Gut microbiota analysis reveals a marked shift to bifidobacteria by a starter infant formula containing a synbiotic of bovine milk-derived oligosaccharides and B ifidobacterium animalis subsp. lactis CNCM I-3446. Environ. Microbiol. 2016, 18, 2185–2195. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Reference | Study Design | Population and Sample Number | HMO BM Analysis | Microbiota Analysis 1 | Outcome Observed (Yes/No) | Main Finding |
|---|---|---|---|---|---|---|
| Tonon et al. [15] | Cross-sectional study | n = 48 pairs of mothers and C-sec and vaginally born BF infants. Sampling at one month postpartum. | HPLC-MS -MSeS: Occurrence of 4 α1-2 fucosylated HMO. |
| Yes | MSeS positively equates the intestinal flora of a child born vaginally or by C-sec. |
| Masi et al. [16] | Cohort study | n = 70 preterm infants (BF or FF-BF + LaBiNIC) (n = 33, NEC) Validation subset: (n = 48: n = 14, NEC). | HPLC-MS -19 most abundant HMOs. -MSeS: presence/absence 2′FL. |
| Yes | Independent of Se status, DSLNT concentration was lower in milk received by infants who showed abnormal microbiome development and developed NEC. |
| Borewicz et al. [17] | Cohort longitudinal Study | n = 24 mother–BF infant pairs. Sampling at 2, 6, and 12 weeks post-partum. | PGC-UPLC-MS and HPAEC-PAD -Total and relative abundance of 18 HMOs. |
| No | Did not observe strong and consistent positive correlations between the HMOs and specific microbial OTUs, including Bifidobacterium. HMO composition is only one of many factors regulating infant gut microbial community. |
| Berger et al. [18] | Randomized double-blinded controlled multicentric clinical trial | n= 175 healthy term FF infants, (n = 87, IF with 2′FL and LNnT and n = 88, normal IF) Control: n = 38 BF infants. Sampling at 3 months post-partum. |
| Yes | The addition of two very specific HMOs (2′FL and LNnT) to IF shifts the microbiota toward the microbiota observed with breastfeeding, the standard in infant nutrition. | |
| Quin et al. [19] | Cohort study | n = 109 mother–BF infant pairs. Sampling at 5 months post-partum. | CE-LIF and HPLC-MS. -MSeS: Relative abundance 2′FL, LDFT, and LNFP1. -16 HMOs bearing sulfate and/or phosphate groups. -Nonsulfonated HMOs: 2′FL, LSTc, and LNP1 |
| Yes | Maternal genetics have a defining role in the establishment of early colonizers (abundance of Enterobacteriaceae was associated with MSeS), but maternal dietary intake during lactation appears to influence the community composition of the infant microbiome. |
| Borewicz et al. [20] | Cohort study | n = 121 healthy, full-term BF infants. Sampling: At, approximately, 1 month postpartum. | UPLC-MS -11 neutral and five acidic HMOs) PAEC-PAD -3′FL |
| Yes | Statistically significant associations between infant fecal microbiota composition and LNFPI and 2′FL levels. Degradation of specific HMOs could be correlated with an increase in relative abundance of various phylotypes within the genus Bifidobacterium and to a lesser extent within the genera Bacteroides and Lactobacillus. |
| Paganini et al. [21] | Double-masked randomized controlled trial study. Cross-sectional study. | n = 80 mother–infant pairs, BF + supplement or not Sampling at baseline of clinical trial, after 3 weeks and after 4 months. | HPAE-PAD -MSeS: presence/absence 2′FL and LNFPI - Total fucosylated sum of 2′FL, 3′FL, LNFPI, LNFPII, and LNFPIII; total sialylated sum of 6′SL, 3′SL, LSTd, LSTa, DSLNT and total non-fucosylated and non-sialylated sum of LNnT, LNT, and LNnH. |
| No | MSeS does not have a major impact on the gut microbiota of the mothers with the exception of a higher abundance of C. perfringens among Se- compared to Se mothers. |
| Bai et al. [22] | Longitud Cohort study | n = 56 mother–vaginally born BF pairs. Sampling at days 6, 42, 120, and 180 post-birth. | LC-QTOF-MS -MSeS: presence/absence LDFT and LNFP I |
| Yes | Bifidobacterial established earlier (and in higher amounts in Se+-fed infants). The relative abundances of this genus continued to increase more than 180 days of lactation in the Se+ group. |
| Korpela et al. [23] | Cohort study | n = 76 mothers–C-sec and vaginally born BF infants. Sampling: BM on day 3 and feces at 3 months. | HPLC-MALDI-TOF and HPAEC -MSeS: 2′FL quantification. |
| Yes | The C-sec born infants of Se+ mothers had a more modest deviation in microbiota composition, compared to those of Se- mothers. |
| Davis et al. [24] | Longitudinal sub-study embedded within a randomized trial. | n = 33 mother–BF infant pairs. Sampling: At 4, 16, and 20 weeks postpartum. | HPLC-TOF. MSeS: α 1-2 fucosylated HMO quantification of 2′ FL, LDFT, TFLNH, DFLNHa, DFLNHc, and IFLNH I. |
| Yes | The microbiome’s ability to break down certain types of oligosaccharides depends on the specific strains that make up the baby’s microbiota. These strains’ variability may contribute to their inability to find functional differences in microbiomes between babies fed by mothers of different secretor status. |
| Underwood et al. [25] | Cohort study | n = 29 preterm BF infants supplemented with B. breve, strain M16-V. Sampling: close to the probiotic start and 3 weeks later. | Nano-HPLC-chip/TOF-MS -MSeS: α(1,2) fucosylated HMOs abundance >6%. |
| No | MSeS was not a significant predictor of response to the administered probiotic B. breve. |
| Matsuki et al. [26] | Randomized, double-blind, placebo-controlled trial | n = 35 FF infants (supplemented with GOS (OM55N). Sampling at the start of the trial and 2 weeks later. |
| Yes | The formula supplementation with GOS (OM55N) stimulated the growth of bifidobacteria and resulted in reduced α-diversity of the gut microbiota. | |
| Smith-Brown et al. [27] | Cohort study | n = 37 BF children 2 years old and 17 eligible mothers (20 excluded due to pregnancy within the previous 12 months) | MSeS was determined from blood and saliva samples using hemagglutination inhibition technique. |
| Yes | Bifidobacterium was increased in the BF children of Se+ mothers compared to Se- mothers. |
| Lewis et al. [28] | Longitudinal cohort study | n = 44 mother–BF infant pairs Sampling: At day 6, 21, 71, and/or 120 postpartum. | Nano-HPLC-chip-TOF-MS. -MSeS: α(1,2) fucosylated HMOs abundance. |
| Yes | Se+ fed infants generally had higher relative amounts of Bifidobacterium and Bacteroides and lower levels of enterobacteria, clostridia, and streptococci. |
| De Leoz et al. [29] | Longitudinal Proof-of-concept study | n = 2 infants (A: BF; B: formula supplementation 4 days and then was solely BF) Sampling: twice/week first month, twice/month second month, and once or twice/month thereafter. | Nano-HPLC-Chip/TOF MS -HMO profile and HMO quantitation to the isomer level (fecal samples). |
| Yes | Fecal HMO profiles correlated with changes in bacterial population. Positive and negative correlations between the fecal isomers of HMO and the relative abundance of bacterial taxa were found at the order level. |
| Wang et al. [30] | Quasi-experimental cohort study | n = 22 mother–infant pairs. (16 BF and 6 FF) Sampling: At 3 months post-partum. | HPLC-Chip/TOF-MS -Until 141 HMOs/sample. |
| Yes | The microbial composition of BF infants is correlated with the presence of HMO in their mother′s milk. |
| Coppa et al. [31] | Cohort study | n = 256 mother–infant pairs. Sampling: At 30 days post-partum. | HPAEC -18 HMOs. -4 BM groups on the basis of the presence or the absence of 2′FL and LNFPII. |
| Yes | No substantial differences in bifidobacteria species composition within infants fed with groups 1, 2, and 3 BM; with group 4 BM (with slight quantity of fucosyloligosaccharides), the microbiota was characterized by a greater frequency of B. adolescentis and the absence of B. catenulatum and harbored a different intestinal microbiota. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez, C.; Fente, C.; Regal, P.; Lamas, A.; Lorenzo, M.P. Human Milk Oligosaccharides (HMOs) and Infant Microbiota: A Scoping Review. Foods 2021, 10, 1429. https://doi.org/10.3390/foods10061429
Sánchez C, Fente C, Regal P, Lamas A, Lorenzo MP. Human Milk Oligosaccharides (HMOs) and Infant Microbiota: A Scoping Review. Foods. 2021; 10(6):1429. https://doi.org/10.3390/foods10061429
Chicago/Turabian StyleSánchez, Cristina, Cristina Fente, Patricia Regal, Alexandre Lamas, and María Paz Lorenzo. 2021. "Human Milk Oligosaccharides (HMOs) and Infant Microbiota: A Scoping Review" Foods 10, no. 6: 1429. https://doi.org/10.3390/foods10061429
APA StyleSánchez, C., Fente, C., Regal, P., Lamas, A., & Lorenzo, M. P. (2021). Human Milk Oligosaccharides (HMOs) and Infant Microbiota: A Scoping Review. Foods, 10(6), 1429. https://doi.org/10.3390/foods10061429