
Lactic Acid Bacteria Exopolysaccharides Producers: A Sustainable Tool for Functional Foods

 ,
,  and
and
Abstract
:1. Introduction
2. Bacterial Synthesis of EPS
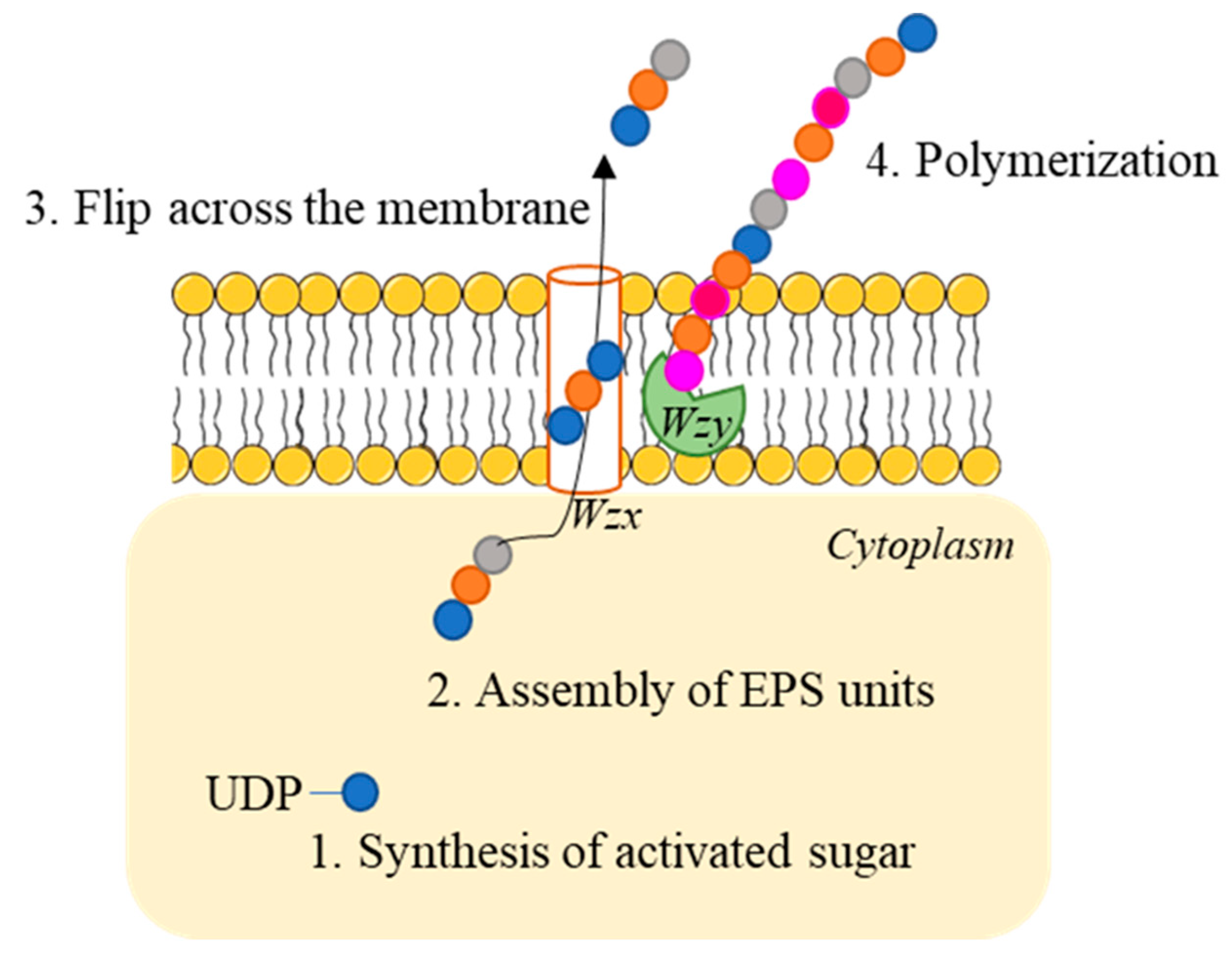
2.1. HoPS Biosynthetic Pathways
2.2. HePS Biosynthetic Pathways
- (1)
- Activated sugar generation: Sugar nucleotides are produced from the glucose-6-phosphate, fructose 6-phosphate or glucose-1-phosphate produced in the Leloir pathway during lactose catabolism [15]. This reaction is catalyzed by priming-GT (a membrane bound polyprenyl-P sugar-1-P transferase).
- (2)
- Assembly of EPS units: sugar nucleotides are added via membrane-bound GT.
- (3)
- Transport across the membrane: repeating units are flipped across the membrane via a flippase (Wzx). This enzyme is bound to the membrane and shows 12 transmembrane domains.
- (4)
- Polimerization: it is catalysed by the Wzy polymerase, which adds single repeating units generating a new glycosidic bond to the reducing end of the chain.
2.3. Genetic of EPS Production
3. EPS-Producing Lactic Acid Bacteria
3.1. In Vitro Screening of EPS-Producing LAB
3.2. EPS Isolation and Purification
3.3. EPS Quantification and Characterization
4. EPS and Health Benefits
4.1. EPS and Antioxidant Properties
4.2. EPS and Cholesterol-Lowering Effect
4.3. EPS and Antibacterial Activity
4.4. Prebiotic Effects of EPS
5. Application of EPS in Dairy Industry
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Sanlibaba, P.; Cakmak, G.A. Exopolysaccharides Production by Lactic Acid Bacteria. Appl. Microbiol. Open Access 2016, 2, 451–469. [Google Scholar] [CrossRef]
- Donot, F.; Fontana, A.; Baccou, J.C.; Schorr-Galindo, S. Microbial Exopolysaccharides: Main Examples of Synthesis, Excretion, Genetics and Extraction. Carbohydr. Polym. 2012, 87, 951–962. [Google Scholar] [CrossRef]
- Flemming, H.-C. EPS—Then and Now. Microorganisms 2016, 4, 41. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.S.; Mody, K.; Jha, B. Bacterial Exopolysaccharides—A Perception. J. Basic Microbiol. 2007, 47, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Galle, S.; Schwab, C.; Dal Bello, F.; Coffey, A.; Gänzle, M.G.; Arendt, E.K. Influence of In-Situ Synthesized Exopolysaccharides on the Quality of Gluten-Free Sorghum Sourdough Bread. Int. J. Food Microbiol. 2012, 155, 105–112. [Google Scholar] [CrossRef]
- Ryan, P.M.; Ross, R.P.; Fitzgerald, G.F.; Caplice, N.M.; Stanton, C. Sugar-Coated: Exopolysaccharide Producing Lactic Acid Bacteria for Food and Human Health Applications. Food Funct. 2015, 6, 679–693. [Google Scholar] [CrossRef]
- Caggianiello, G.; Kleerebezem, M.; Spano, G. Exopolysaccharides Produced by Lactic Acid Bacteria: From Health-Promoting Benefits to Stress Tolerance Mechanisms. Appl. Microbiol. Biotechnol. 2016, 100, 3877–3886. [Google Scholar] [CrossRef]
- Lynch, K.M.; Zannini, E.; Coffey, A.; Arendt, E.K. Lactic Acid Bacteria Exopolysaccharides in Foods and Beverages: Isolation, Properties, Characterization, and Health Benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 155–176. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A Taxonomic Note on the Genus Lactobacillus: Description of 23 Novel Genera, Emended Description of the Genus Lactobacillus beijerinck 1901, and Union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Castro-Bravo, N.; Wells, J.M.; Margolles, A.; Ruas-Madiedo, P. Interactions of Surface Exopolysaccharides from Bifidobacterium and Lactobacillus within the Intestinal Environment. Front. Microbiol. 2018, 9, 2426. [Google Scholar] [CrossRef] [Green Version]
- Van Kranenburg, R.; Vos, H.R.; Van Swam, I.I.; Kleerebezem, M.; De Vos, W.M. Functional Analysis of Glycosyltransferase Genes from Lactococcus lactis and Other Gram-Positive Cocci: Complementation, Expression, and Diversity. J. Bacteriol. 1999, 181, 6347–6353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mozzi, F.; Vaningelgem, F.; Hébert, E.M.; Van Der Meulen, R.; Moreno, M.R.F.; Font De Valdez, G.; De Vuyst, L. Diversity of Heteropolysaccharide-Producing Lactic Acid Bacterium Strains and Their Biopolymers. Appl. Environ. Microbiol. 2006, 72, 4431–4435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torino, M.I.; de Valdez, G.F.; Mozzi, F. Biopolymers from Lactic Acid Bacteria. Novel Applications in Foods and Beverages. Front. Microbiol. 2015, 6, 834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapot-Chartier, M.P.; Kulakauskas, S. Cell Wall Structure and Function in Lactic Acid Bacteria. Microb. Cell Fact. 2014, 13 (Suppl. 1), S9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leemhuis, H.; Pijning, T.; Dobruchowska, J.M.; van Leeuwen, S.S.; Kralj, S.; Dijkstra, B.W.; Dijkhuizen, L. Glucansucrases: Three-Dimensional Structures, Reactions, Mechanism, α-Glucan Analysis and Their Implications in Biotechnology and Food Applications. J. Biotechnol. 2013, 163, 250–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeidan, A.A.; Poulsen, V.K.; Janzen, T.; Buldo, P.; Derkx, P.M.F.; Øregaard, G.; Neves, A.R. Polysaccharide Production by Lactic Acid Bacteria: From Genes to Industrial Applications. FEMS Microbiol. Rev. 2017, 41, S168–S200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deo, D.; Davray, D.; Kulkarni, R. A Diverse Repertoire of Exopolysaccharide Biosynthesis Gene Clusters in Lactobacillus Revealed by Comparative Analysis in 106 Sequenced Genomes. Microorganisms 2019, 7, 444. [Google Scholar] [CrossRef] [Green Version]
- Remus, D.M.; van Kranenburg, R.; van Swam, I.I.; Taverne, N.; Bongers, R.S.; Wels, M.; Wells, J.M.; Bron, P.A.; Kleerebezem, M. Impact of 4 Lactobacillus plantarum Capsular Polysaccharide Clusters on Surface Glycan Composition and Host Cell Signaling. Microb. Cell Fact. 2012, 11, 149. [Google Scholar] [CrossRef] [Green Version]
- Knoshaug, E.P.; Ahlgren, J.A.; Trempy, J.E. Exopolysaccharide Expression in Lactococcus lactis subsp. cremoris Ropy 352: Evidence for Novel Gene Organization. Appl. Environ. Microbiol. 2007, 73, 897–905. [Google Scholar] [CrossRef] [Green Version]
- Goh, Y.J.; Goin, C.; O’Flaherty, S.; Altermann, E.; Hutkins, R. Specialized Adaptation of a Lactic Acid Bacterium to the Milk Environment: The Comparative Genomics of Streptococcus thermophilus LMD-9. Microb. Cell Fact. 2011, 10 (Suppl. 1), S22. [Google Scholar] [CrossRef] [Green Version]
- Mistou, M.Y.; Sutcliffe, I.C.; Van Sorge, N.M. Bacterial Glycobiology: Rhamnose-Containing Cell Wall Polysaccharides in Gram-Positive Bacteria. FEMS Microbiol. Rev. 2016, 40, 464–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thevenard, B.; Besset, C.; Choinard, S.; Fourcassié, P.; Boyaval, P.; Monnet, V.; Rul, F. Response of S. thermophilus LMD-9 to Bacitracin: Involvement of a BceRS/AB-like Module and of the Rhamnose-Glucose Polysaccharide Synthesis Pathway. Int. J. Food Microbiol. 2014, 177, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Mahony, J.; Kot, W.; Murphy, J.; Ainsworth, S.; Neve, H.; Hansen, L.H.; Heller, K.J.; Sørensen, S.J.; Hammer, K.; Cambillau, C.; et al. Investigation of the Relationship between Lactococcal Host Cell Wall Polysaccharide Genotype and 936 Phage Receptor Binding Protein Phylogeny. Appl. Environ. Microbiol. 2013, 79, 4385–4392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badel, S.; Bernardi, T.; Michaud, P. New Perspectives for Lactobacilli Exopolysaccharides. Biotechnol. Adv. 2011, 29, 54–66. [Google Scholar] [CrossRef]
- Khalil, E.S.; Manap, M.Y.A.; Mustafa, S.; Alhelli, A.M.; Shokryazdan, P. Probiotic Properties of Exopolysaccharide-Producing Lactobacillus Strains Isolated from Tempoyak. Molecules 2018, 23, 398. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Li, F.; Li, L.; Huang, L.; Li, Q. Genetic and Biochemical Characterization of an Exopolysaccharide With in Vitro Antitumoral Activity Produced by Lactobacillus fermentum YL-11. Front. Microbiol. 2019, 10, 2898. [Google Scholar] [CrossRef]
- Tukenmez, U.; Aktas, B.; Aslim, B.; Yavuz, S. The Relationship between the Structural Characteristics of Lactobacilli-EPS and Its Ability to Induce Apoptosis in Colon Cancer Cells in Vitro. Sci. Rep. 2019, 9, 8268. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, H.; Miyamoto, T. Production of Exopolysaccharide by Lactobacillus plantarum and the Prebiotic Activity of the Exopolysaccharide. Food Sci. Technol. Res. 2010, 16, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Macedo, M.G.; Laporte, M.F.; Lacroix, C. Quantification of Exopolysaccharide, Lactic Acid, and Lactose Concentrations in Culture Broth by near-Infrared Spectroscopy. J. Agric. Food Chem. 2002, 50, 1774–1779. [Google Scholar] [CrossRef]
- Maeda, H.; Zhu, X.; Suzuki, S.; Suzuki, K.; Kitamura, S. Structural Characterization and Biological Activities of an Exopolysaccharide Kefiran Produced by Lactobacillus kefiranofaciens WT-2B T. J. Agric. Food Chem. 2004, 52, 5533–5538. [Google Scholar] [CrossRef]
- Sasikumar, K.; Kozhummal Vaikkath, D.; Devendra, L.; Nampoothiri, K.M. An Exopolysaccharide (EPS) from a Lactobacillus plantarum BR2 with Potential Benefits for Making Functional Foods. Bioresour. Technol. 2017, 241, 1152–1156. [Google Scholar] [CrossRef] [PubMed]
- Kšonžeková, P.; Bystrický, P.; Vlčková, S.; Pätoprstý, V.; Pulzová, L.; Mudroňová, D.; Kubašková, T.; Csank, T.; Tkáčiková, L. Exopolysaccharides of Lactobacillus reuteri: Their Influence on Adherence of E. Coli to Epithelial Cells and Inflammatory Response. Carbohydr. Polym. 2016, 141, 10–19. [Google Scholar] [CrossRef]
- Nachtigall, C.; Surber, G.; Herbi, F.; Wefers, D.; Jaros, D.; Rohm, H. Production and Molecular Structure of Heteropolysaccharides from Two Lactic Acid Bacteria. Carbohydr. Polym. 2020, 236, 116019. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, A.; Tong, Y.; Wu, Q.; Zhang, J.; Shah, N.P. Transcriptomic Insights into the Growth Phase- and Sugar-Associated Changes in the Exopolysaccharide Production of a High EPS-Producing Streptococcus thermophilus ASCC 1275. Front. Microbiol. 2018, 9, 1919. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Guo, Q.; Zhang, H.; Wu, Y.; Hang, X.; Ai, L. Exopolysaccharide Produced by Streptococcus thermophiles S-3: Molecular, Partial Structural and Rheological Properties. Carbohydr. Polym. 2018, 194, 132–138. [Google Scholar] [CrossRef]
- Kanamarlapudi, S.L.R.K.; Muddada, S. Characterization of Exopolysaccharide Produced by Streptococcus thermophilus CC30. BioMed Res. Int. 2017, 2017, 4201809. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Li, J.; Zhao, F.; Wang, G.; Qin, Q.; Hao, Y. The Influence of Fermentation Condition on Production and Molecular Mass of EPS Produced by Streptococcus thermophilus 05-34 in Milk-Based Medium. Food Chem. 2016, 197, 367–372. [Google Scholar] [CrossRef]
- Zhang, J.; Cao, Y.; Wang, J.; Guo, X.; Zheng, Y.; Zhao, W.; Mei, X.; Guo, T.; Yang, Z. Physicochemical Characteristics and Bioactivities of the Exopolysaccharide and Its Sulphated Polymer from Streptococcus thermophilus GST-6. Carbohydr. Polym. 2016, 146, 368–375. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, C.; Li, S.; Zhang, Y.; Yang, Z. Growth and Exopolysaccharide Production by Streptococcus thermophilus ST1 in Skim Milk. Braz. J. Microbiol. 2011, 42, 1470–1478. [Google Scholar] [CrossRef] [Green Version]
- Marshall, V.M.; Laws, A.P.; Gu, Y.; Levander, F.; Rådström, P.; De Vuyst, L.; Degeest, B.; Vaningelgem, F.; Dunn, H.; Elvin, M. Exopolysaccharide-Producing Strains of Thermophilic Lactic Acid Bacteria Cluster into Groups According to Their EPS Structure. Lett. Appl. Microbiol. 2001, 32, 433–437. [Google Scholar] [CrossRef]
- Almiron-Roig, E.; Mulholland, F.; Gasson, M.J.; Griffin, A.M. The Complete Cps Gene Cluster from Streptococcus thermophilus NCFB 2393 Involved in the Biosynthesis of a New Exopolysaccharide. Microbiology 2000, 146, 2793–2802. [Google Scholar] [CrossRef] [Green Version]
- Adebayo-Tayo, B.; Fashogbon, R. In Vitro Antioxidant, Antibacterial, in Vivo Immunomodulatory, Antitumor and Hematological Potential of Exopolysaccharide Produced by Wild Type and Mutant Lactobacillus delbureckii subsp. bulgaricus. Heliyon 2020, 6, e03268. [Google Scholar] [CrossRef]
- Bancalari, E.; Alinovi, M.; Bottari, B.; Caligiani, A.; Mucchetti, G.; Gatti, M. Ability of a Wild Weissella Strain to Modify Viscosity of Fermented Milk. Front. Microbiol. 2020, 10, 3086. [Google Scholar] [CrossRef]
- Tang, W.; Dong, M.; Wang, W.; Han, S.; Rui, X.; Chen, X.; Jiang, M.; Zhang, Q.; Wu, J.; Li, W. Structural Characterization and Antioxidant Property of Released Exopolysaccharides from Lactobacillus delbrueckii ssp. bulgaricus SRFM-1. Carbohydr. Polym. 2017, 173, 654–664. [Google Scholar] [CrossRef] [PubMed]
- Van Calsteren, M.R.; Gagnon, F.; Nishimura, J.; Makino, S. Structure Determination of the Neutral Exopolysaccharide Produced by Lactobacillus delbrueckii subsp. bulgaricus OLL1073R-1. Carbohydr. Res. 2015, 413, 115–122. [Google Scholar] [CrossRef]
- Abdellah, M.; Ahcne, H.; Benalia, Y.; Saad, B.; Abdelmalek, B. Evaluation of Biofilm Formation by Exopolysaccharide-Producer Strains of Thermophilic Lactic Acid Bacteria Isolated from Algerian Camel Milk. Emir. J. Food Agric. 2015, 27, 513. [Google Scholar] [CrossRef]
- You, X.; Li, Z.; Ma, K.; Zhang, C.; Chen, X.; Wang, G.; Yang, L.; Dong, M.; Rui, X.; Zhang, Q.; et al. Structural Characterization and Immunomodulatory Activity of an Exopolysaccharide Produced by Lactobacillus helveticus LZ-R-5. Carbohydr. Polym. 2020, 235, 115977. [Google Scholar] [CrossRef] [PubMed]
- Min, W.H.; Fang, X.B.; Wu, T.; Fang, L.; Liu, C.L.; Wang, J. Characterization and Antioxidant Activity of an Acidic Exopolysaccharide from Lactobacillus plantarum JLAU103. J. Biosci. Bioeng. 2019, 127, 758–766. [Google Scholar] [CrossRef]
- Wang, J.; Wu, T.; Fang, X.; Min, W.; Yang, Z. Characterization and Immunomodulatory Activity of an Exopolysaccharide Produced by Lactobacillus plantarum JLK0142 Isolated from Fermented Dairy Tofu. Int. J. Biol. Macromol. 2018, 115, 985–993. [Google Scholar] [CrossRef] [PubMed]
- Ziadi, M.; Bouzaiene, T.; M’Hir, S.; Zaafouri, K.; Mokhtar, F.; Hamdi, M.; Boisset-Helbert, C. Evaluation of the Efficiency of Ethanol Precipitation and Ultrafiltration on the Purification and Characteristics of Exopolysaccharides Produced by Three Lactic Acid Bacteria. BioMed Res. Int. 2018, 2018, 1896240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Calsteren, M.R.; Pau-Roblot, C.; Bégin, A.; Roy, D. Structure Determination of the Exopolysaccharide Produced by Lactobacillus rhamnosus Strains RW-9595M and R. Biochem. J. 2002, 363, 7–17. [Google Scholar] [CrossRef]
- Ayala-Hernández, I.; Hassan, A.; Goff, H.D.; Mira de Orduña, R.; Corredig, M. Production, Isolation and Characterization of Exopolysaccharides Produced by Lactococcus lactis subsp. cremoris JFR1 and Their Interaction with Milk Proteins: Effect of PH and Media Composition. Int. Dairy J. 2008, 18, 1109–1118. [Google Scholar] [CrossRef]
- Leroy, F.; De Vuyst, L. Advances in Production and Simplified Methods for Recovery and Quantification of Exopolysaccharides for Applications in Food and Health. J. Dairy Sci. 2016, 99, 3229–3238. [Google Scholar] [CrossRef] [Green Version]
- Jolly, L.; Vincent, S.J.F.; Duboc, P.; Neeser, J.R. Exploiting Exopolysaccharides from Lactic Acid Bacteria. Antonie van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2002, 82, 367–374. [Google Scholar] [CrossRef]
- Cirrincione, S.; Breuer, Y.; Mangiapane, E.; Mazzoli, R.; Pessione, E. ’Ropy’ Phenotype, Exopolysaccharides and Metabolism: Study on Food Isolated Potential Probiotics LAB. Microbiol. Res. 2018, 214, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Schwab, C.; Mastrangelo, M.; Corsetti, A.; Gänzle, M. Formation of Oligosaccharides and Polysaccharides by Lactobacillus reuteri LTH5448 and Weissella cibaria 10M in Sorghum Sourdoughs. Cereal Chem. 2008, 85, 679–684. [Google Scholar] [CrossRef]
- Vaningelgem, F.; Zamfir, M.; Mozzi, F.; Adriany, T.; Vancanneyt, M.; Swings, J.; De Vuyst, L. Biodiversity of Exopolysaccharides Produced by Streptococcus thermophilus Strains Is Reflected in Their Production and Their Molecular and Functional Characteristics. Appl. Environ. Microbiol. 2004, 70, 900–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruas-Madiedo, P.; De Los Reyes-Gavilán, C.G. Invited Review: Methods for the Screening, Isolation, and Characterization of Exopolysaccharides Produced by Lactic Acid Bacteria. J. Dairy Sci. 2005, 88, 843–856. [Google Scholar] [CrossRef] [Green Version]
- Rabha, B.; Nadra, R.-S.; Ahmed, B. Effect of Some Fermentation Substrates and Growth Temperature on Exopolysaccharide Production by Streptococcus thermophilus BN1. Int. J. Biosci. Biochem. Bioinform. 2012, 2, 44–47. [Google Scholar] [CrossRef] [Green Version]
- Petry, S.; Furlan, S.; Crepeau, M.J.; Cerning, J.; Desmazeaud, M. Factors Affecting Exocellular Polysaccharide Production by Lactobacillus delbrueckii subsp. bulgaricus Grown in a Chemically Defined Medium. Appl. Environ. Microbiol. 2000, 66, 3427–3431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Bi, J. Modification of Characteristics of Kefiran by Changing the Carbon Source of Lactobacillus kefiranofaciens. J. Sci. Food Agric. 2008, 88, 763–769. [Google Scholar] [CrossRef]
- Zajšek, K.; Goršek, A.; Kolar, M. Cultivating Conditions Effects on Kefiran Production by the Mixed Culture of Lactic Acid Bacteria Imbedded within Kefir Grains. Food Chem. 2013, 139, 970–977. [Google Scholar] [CrossRef] [PubMed]
- Degeest, B.; Janssens, B.; De Vuyst, L. Exopolysaccharide (EPS) Biosynthesis by Lactobacillus sakei 0-1: Production Kinetics, Enzyme Activities and EPS Yields. J. Appl. Microbiol. 2001, 91, 470–477. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, S.; Zhang, C.; Luo, Y.; Zhang, H.; Yang, Z. Growth and Exopolysaccharide Production by Lactobacillus fermentum F6 in Skim Milk. Afr. J. Biotechnol. 2011, 10, 2080–2091. [Google Scholar] [CrossRef]
- Pham, P.L.; Dupont, I.; Roy, D.; Lapointe, G.; Cerning, J. Production of Exopolysaccharide by Lactobacillus rhamnosus R and Analysis of Its Enzymatic Degradation during Prolonged Fermentation. Appl. Environ. Microbiol. 2000, 66, 2302–2310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, T.Y.; Chien, M.F.C. Exopolysaccharides Production as Affected by Lactic Acid Bacteria and Fermentation Time. Food Chem. 2007, 100, 1419–1423. [Google Scholar] [CrossRef]
- Mende, S.; Krzyzanowski, L.; Weber, J.; Jaros, D.; Rohm, H. Growth and Exopolysaccharide Yield of Lactobacillus delbrueckii ssp. bulgaricus DSM 20081 in Batch and Continuous Bioreactor Experiments at Constant PH. J. Biosci. Bioeng. 2012, 113, 185–191. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; De Vin, F.; Vaningelgem, F.; Degeest, B. Recent Developments in the Biosynthesis and Applications of Heteropolysaccharides from Lactic Acid Bacteria. Int. Dairy J. 2001, 11, 687–707. [Google Scholar] [CrossRef]
- Leo, F.; Hashida, S.; Kumagai, D.; Uchida, K.; Motoshima, H.; Arai, I.; Asakuma, S.; Fukuda, K.; Urashima, T. Studies on a Neutral Exopolysaccharide of Lactobacillus fermentum TDS030603. J. Appl. Glycosci. 2007, 54, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Ruas-Madiedo, P.; Tuinier, R.; Kanning, M.; Zoon, P. Role of Exopolysaccharides Produced by Lactococcus lactis subsp. cremoris on the Viscosity of Fermented Milks. Int. Dairy J. 2002, 12, 689–695. [Google Scholar] [CrossRef]
- Ahmed, Z.; Wang, Y.; Anjum, N.; Ahmad, H.; Ahmad, A.; Raza, M. Characterization of New Exopolysaccharides Produced by Coculturing of L. kefiranofaciens with Yoghurt Strains. Int. J. Biol. Macromol. 2013, 59, 377–383. [Google Scholar] [CrossRef]
- Bertsch, A.; Roy, D.; LaPointe, G. Enhanced Exopolysaccharide Production by Lactobacillus rhamnosus in Co-Culture with Saccharomyces cerevisiae. Appl. Sci. 2019, 9, 4026. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.J.; Chen, Z.; Chen, P.T.; Ng, I.S. Production, Characterization and Antibacterial Activity of Exopolysaccharide from a Newly Isolated Weissella Cibaria under Sucrose Effect. J. Biosci. Bioeng. 2018, 126, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Zannini, E.; Mauch, A.; Galle, S.; Gänzle, M.; Coffey, A.; Arendt, E.K.; Taylor, J.P.; Waters, D.M. Barley Malt Wort Fermentation by Exopolysaccharide-Forming Weissella Cibaria MG1 for the Production of a Novel Beverage. J. Appl. Microbiol. 2013, 115, 1379–1387. [Google Scholar] [CrossRef] [PubMed]
- Dertli, E.; Mercan, E.; Arici, M.; Yilmaz, M.T.; Sağdiç, O. Characterisation of Lactic Acid Bacteria from Turkish Sourdough and Determination of Their Exopolysaccharide (EPS) Production Characteristics. LWT-Food Sci. Technol. 2016, 71, 116–124. [Google Scholar] [CrossRef]
- Imran, M.Y.M.; Reehana, N.; Jayaraj, K.A.; Ahamed, A.A.P.; Dhanasekaran, D.; Thajuddin, N.; Alharbi, N.S.; Muralitharan, G. Statistical Optimization of Exopolysaccharide Production by Lactobacillus Plantarum NTMI05 and NTMI20. Int. J. Biol. Macromol. 2016, 93, 731–745. [Google Scholar] [CrossRef]
- Abdalrahim, S.; Zohri, A.N.A.; Khider, M.; Kamal El-Dean, A.M.; Abulreesh, H.H.; Ahmad, I.; Elbanna, K. Phenotypic and Genotypic Characterization of Exopolysaccharide Producing Bacteria Isolated from Fermented Fruits, Vegetables and Dairy Products. J. Pure Appl. Microbiol. 2019, 13, 1349–1362. [Google Scholar] [CrossRef] [Green Version]
- Salazar, N.; Prieto, A.; Leal, J.A.; Mayo, B.; Bada-Gancedo, J.C.; de los Reyes-Gavilán, C.G.; Ruas-Madiedo, P. Production of Exopolysaccharides by Lactobacillus and Bifidobacterium Strains of Human Origin, and Metabolic Activity of the Producing Bacteria in Milk. J. Dairy Sci. 2009, 92, 4158–4168. [Google Scholar] [CrossRef] [Green Version]
- Ruas-Madiedo, P.; Gueimonde, M.; Arigoni, F.; De Los Reyes-Gavilán, C.G.; Margolles, A. Bile Affects the Synthesis of Exopolysaccharides by Bifidobacterium animalis. Appl. Environ. Microbiol. 2009, 75, 1204–1207. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Shah, N.P. Comparative MRNA-Seq Analysis Reveals the Improved EPS Production Machinery in Streptococcus Thermophilus ASCC 1275 during Optimized Milk Fermentation. Front. Microbiol. 2018, 9, 445. [Google Scholar] [CrossRef] [Green Version]
- Stack, H.M.; Kearney, N.; Stanton, C.; Fitzgerald, G.F.; Ross, R.P. Association of Beta-Glucan Endogenous Production with Increased Stress Tolerance of Intestinal Lactobacilli. Appl. Environ. Microbiol. 2010, 76, 500–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degeest, B.; Mozzi, F.; De Vuyst, L. Effect of Medium Composition and Temperature and PH Changes on Exopolysaccharide Yields and Stability during Streptococcus thermophilus LY03 Fermentations. Int. J. Food Microbiol. 2002, 79, 161–174. [Google Scholar] [CrossRef]
- Yang, Z.; Li, S.; Zhang, X.; Zeng, X.; Li, D.; Zhao, Y.; Zhang, J. Capsular and Slime-Polysaccharide Production by Lactobacillus rhamnosus JAAS8 Isolated from Chinese Sauerkraut: Potential Application in Fermented Milk Products. J. Biosci. Bioeng. 2010, 110, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, Z.; Wang, Y.; Anjum, N.; Ahmad, A.; Khan, S.T. Characterization of Exopolysaccharide Produced by Lactobacillus kefiranofaciens ZW3 Isolated from Tibet Kefir-Part II. Food Hydrocoll. 2013, 30, 343–350. [Google Scholar] [CrossRef]
- Saadat, Y.R.; Khosroushahi, A.Y.; Gargari, B.P. A Comprehensive Review of Anticancer, Immunomodulatory and Health Beneficial Effects of the Lactic Acid Bacteria Exopolysaccharides. Carbohydr. Polym. 2019, 217, 79–89. [Google Scholar] [CrossRef]
- Ortega-Morales, B.O.; Santiago-García, J.L.; Chan-Bacab, M.J.; Moppert, X.; Miranda-Tello, E.; Fardeau, M.L.; Carrero, J.C.; Bartolo-Pérez, P.; Valadéz-González, A.; Guezennec, J. Characterization of Extracellular Polymers Synthesized by Tropical Intertidal Biofilm Bacteria. J. Appl. Microbiol. 2007, 102, 254–264. [Google Scholar] [CrossRef] [Green Version]
- Borucki, M.K.; Peppin, J.D.; White, D.; Loge, F.; Call, D.R. Variation in Biofilm Formation among Strains of Listeria monocytogenes. Appl. Environ. Microbiol. 2003, 69, 7336–7342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Yin, R. Primary Study on Extracellular Polysaccharide Producing Bacteria in Different Environments. J. Anhui Univ. Nat. Sci. Ed. 2011, 35, 94–100. [Google Scholar]
- Lauer Cruz, K.; de Souza da Motta, A. Characterization of Biofilm Production by Pseudomonas fluorescens Isolated from Refrigerated Raw Buffalo Milk. J. Food Sci. Technol. 2019, 56, 4595–4604. [Google Scholar] [CrossRef]
- Ates, O. Systems Biology of Microbial Exopolysaccharides Production. Front. Bioeng. Biotechnol. 2015, 3, 200. [Google Scholar] [CrossRef] [Green Version]
- Teusink, B.; Wiersma, A.; Jacobs, L.; Notebaart, R.; Smid, E. Understanding the Adaptive Growth Strategy of Lactobacillus plantarum by In Silico Optimisation. PLoS Comput. Biol. 2009, 5, e1000410. [Google Scholar] [CrossRef]
- Siezen, R.J.; van Hylckama Vlieg, J.E.T. Genomic Diversity and Versatility of Lactobacillus plantarum, a Natural Metabolic Engineer. Microb. Cell Fact. 2011, 10 (Suppl. 1), S3. [Google Scholar] [CrossRef] [Green Version]
- Hao, P.; Zheng, H.; Yu, Y.; Ding, G.; Gu, W.; Chen, S.; Yu, Z.; Ren, S.; Oda, M.; Konno, T.; et al. Complete Sequencing and Pan-Genomic Analysis of Lactobacillus delbrueckii subsp. bulgaricus Reveal Its Genetic Basis for Industrial Yogurt Production. PLoS ONE 2011, 6, e15964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.Y.; Lee, S.H.; Kim, J.M.; Park, M.S.; Bae, J.W.; Hahn, Y.; Madsen, E.L.; Jeon, C.O. Metagenomic Analysis of Kimchi, a Traditional Korean Fermented Food. Appl. Environ. Microbiol. 2011, 77, 2264–2274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieuwerts, S.; Molenaar, D.; Van Hijum, S.A.F.T.; Beerthuyzen, M.; Stevens, M.J.A.; Janssen, P.W.M.; Ingham, C.J.; De Bok, F.A.M.; De Vos, W.M.; Van Hylckama Vlieg, J.E.T. Mixed-Culture Transcriptome Analysis Reveals the Molecular Basis of Mixed-Culture Growth in Streptococcus thermophilus and Lactobacillus bulgaricus. Appl. Environ. Microbiol. 2010, 76, 7775–7784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouzar, F.; Cerning, J.; Desmazeaud, M. Exopolysaccharide Production and Texture-Promoting Abilities of Mixed-Strain Starter Cultures in Yogurt Production. J. Dairy Sci. 1997, 80, 2310–2317. [Google Scholar] [CrossRef]
- Rimada, P.S.; Abraham, A.G. Comparative Study of Different Methodologies to Determine the Exopolysaccharide Produced by Kefir Grains in Milk and Whey. Lait 2003, 83, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Notararigo, S.; Nácher-Vázquez, M.; Ibarburu, I.; Werning, M.; De Palencia, P.F.; Dueñas, M.T.; Aznar, R.; López, P.; Prieto, A. Comparative Analysis of Production and Purification of Homo- and Hetero-Polysaccharides Produced by Lactic Acid Bacteria. Carbohydr. Polym. 2013, 93, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Li, W.; Chen, X.; Feng, M.; Rui, X.; Jiang, M.; Dong, M. Microbiological, Physicochemical and Rheological Properties of Fermented Soymilk Produced with Exopolysaccharide (EPS) Producing Lactic Acid Bacteria Strains. LWT-Food Sci. Technol. 2014, 57, 477–485. [Google Scholar] [CrossRef]
- Abdhul, K.; Ganesh, M.; Shanmughapriya, S.; Kanagavel, M.; Anbarasu, K.; Natarajaseenivasan, K. Antioxidant Activity of Exopolysaccharide from Probiotic Strain Enterococcus faecium (BDU7) from Ngari. Int. J. Biol. Macromol. 2014, 70, 450–454. [Google Scholar] [CrossRef]
- Donnarumma, G.; Molinaro, A.; Cimini, D.; De Castro, C.; Valli, V.; De Gregorio, V.; De Rosa, M.; Schiraldi, C. Lactobacillus crispatus L1: High Cell Density Cultivation and Exopolysaccharide Structure Characterization to Highlight Potentially Beneficial Effects against Vaginal Pathogens. BMC Microbiol. 2014, 14, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pino, A.; Bartolo, E.; Caggia, C.; Cianci, A.; Randazzo, C.L. Detection of Vaginal Lactobacilli as Probiotic Candidates. Sci. Rep. 2019, 9, 3355. [Google Scholar] [CrossRef] [PubMed]
- Behare, P.V.; Singh, R.; Nagpal, R.; Rao, K.H. Exopolysaccharides Producing Lactobacillus fermentum Strain for Enhancing Rheological and Sensory Attributes of Low-Fat Dahi. J. Food Sci. Technol. 2013, 50, 1228–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimopoulou, M.; Vuillemin, M.; Campbell-Sills, H.; Lucas, P.M.; Ballestra, P.; Miot-Sertier, C.; Favier, M.; Coulon, J.; Moine, V.; Doco, T.; et al. Exopolysaccharide (EPS) Synthesis by Oenococcus oeni: From Genes to Phenotypes. PLoS ONE 2014, 9, e98898. [Google Scholar] [CrossRef] [Green Version]
- Ruijssenaars, H.J.; Stingele, F.; Hartmans, S. Biodegradability of Food-Associated Extracellular Polysaccharides. Curr. Microbiol. 2000, 40, 194–199. [Google Scholar] [CrossRef]
- Katina, K.; Maina, N.H.; Juvonen, R.; Flander, L.; Johansson, L.; Virkki, L.; Tenkanen, M.; Laitila, A. In Situ Production and Analysis of Weissella confusa Dextran in Wheat Sourdough. Food Microbiol. 2009, 26, 734–743. [Google Scholar] [CrossRef]
- Yilmaz, M.T.; Dertli, E.; Toker, O.S.; Tatlisu, N.B.; Sagdic, O.; Arici, M. Effect of in Situ Exopolysaccharide Production on Physicochemical, Rheological, Sensory, and Microstructural Properties of the Yogurt Drink Ayran: An Optimization Study Based on Fermentation Kinetics. J. Dairy Sci. 2015, 98, 1604–1624. [Google Scholar] [CrossRef]
- Angelin, J.; Kavitha, M. Exopolysaccharides from Probiotic Bacteria and Their Health Potential. Int. J. Biol. Macromol. 2020, 162, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Niu, M.; Song, D.; Song, X.; Zhao, J.; Wu, Y.; Lu, B.; Niu, G. Preparation, Partial Characterization and Biological Activity of Exopolysaccharides Produced from Lactobacillus fermentum S1. J. Biosci. Bioeng. 2020, 129, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Zannini, E.; Waters, D.M.; Coffey, A.; Arendt, E.K. Production, Properties, and Industrial Food Application of Lactic Acid Bacteria-Derived Exopolysaccharides. Appl. Microbiol. Biotechnol. 2016, 100, 1121–1135. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- Liu, Z.; Ren, Z.; Zhang, J.; Chuang, C.C.; Kandaswamy, E.; Zhou, T.; Zuo, L. Role of ROS and Nutritional Antioxidants in Human Diseases. Front. Physiol. 2018, 9, 477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, V.; Shah, C.; Mokashe, N.; Chavan, R.; Yadav, H.; Prajapati, J. Probiotics as Potential Antioxidants: A Systematic Review. J. Agric. Food Chem. 2015, 63, 3615–3626. [Google Scholar] [CrossRef] [PubMed]
- Prete, R.; Garcia-Gonzalez, N.; Di Mattia, C.D.; Corsetti, A.; Battista, N. Food-Borne Lactiplantibacillus plantarum Protect Normal Intestinal Cells against Inflammation by Modulating Reactive Oxygen Species and IL-23/IL-17 Axis. Sci. Rep. 2020, 10, 16340. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, C.; Li, D.; Zhao, Y.; Zhang, X.; Zeng, X.; Yang, Z.; Li, S. Antioxidant Activity of an Exopolysaccharide Isolated from Lactobacillus plantarum C88. Int. J. Biol. Macromol. 2013, 54, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Seo, B.-J.; Bajpai, V.K.; Rather, I.A.; Park, Y.-H. Partially Purified Exopolysaccharide from Lactobacillus plantarum YML009 with Total Phenolic Content, Antioxidant and Free Radical Scavenging Efficacy. Indian J. Pharm. Educ. Res. 2015, 49, 282–292. [Google Scholar] [CrossRef] [Green Version]
- Trabelsi, I.; Ktari, N.; Ben Slima, S.; Triki, M.; Bardaa, S.; Mnif, H.; Ben Salah, R. Evaluation of Dermal Wound Healing Activity and in Vitro Antibacterial and Antioxidant Activities of a New Exopolysaccharide Produced by Lactobacillus sp.Ca6. Int. J. Biol. Macromol. 2017, 103, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Şengül, N.; Aslím, B.; Uçar, G.; Yücel, N.; Işik, S.; Bozkurt, H.; Sakaoğullarí, Z.; Atalay, F. Effects of Exopolysaccharide-Producing Probiotic Strains on Experimental Colitis in Rats. Dis. Colon Rectum 2006, 49, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.F.; Tseng, K.C.; Chiang, S.S.; Lee, B.H.; Hsu, W.H.; Pan, T.M. Immunomodulatory and Antioxidant Potential of Lactobacillus Exopolysaccharides. J. Sci. Food Agric. 2011, 91, 2284–2291. [Google Scholar] [CrossRef]
- Polak-Berecka, M.; Waśko, A.; Szwajgier, D.; Choma, A. Bifidogenic and Antioxidant Activity of Exopolysaccharides Produced by Lactobacillus rhamnosus E/N Cultivated on Different Carbon Sources. Polish J. Microbiol. 2013, 62, 181–188. [Google Scholar] [CrossRef]
- Li, J.Y.; Jin, M.M.; Meng, J.; Gao, S.M.; Lu, R.R. Exopolysaccharide from Lactobacillus planterum LP6: Antioxidation and the Effect on Oxidative Stress. Carbohydr. Polym. 2013, 98, 1147–1152. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Ji, J.; Chen, X.; Jiang, M.; Rui, X.; Dong, M. Structural Elucidation and Antioxidant Activities of Exopolysaccharides from Lactobacillus helveticus MB2-1. Carbohydr. Polym. 2014, 102, 351–359. [Google Scholar] [CrossRef]
- Rani, R.P.; Anandharaj, M.; David Ravindran, A. Characterization of a Novel Exopolysaccharide Produced by Lactobacillus gasseri FR4 and Demonstration of Its in Vitro Biological Properties. Int. J. Biol. Macromol. 2018, 109, 772–783. [Google Scholar] [CrossRef]
- Guo, Y.; Pan, D.; Li, H.; Sun, Y.; Zeng, X.; Yan, B. Antioxidant and Immunomodulatory Activity of Selenium Exopolysaccharide Produced by Lactococcus lactis subsp. lactis. Food Chem. 2013, 138, 84–89. [Google Scholar] [CrossRef]
- Dilna, S.V.; Surya, H.; Aswathy, R.G.; Varsha, K.K.; Sakthikumar, D.N.; Pandey, A.; Nampoothiri, K.M. Characterization of an Exopolysaccharide with Potential Health-Benefit Properties from a Probiotic Lactobacillus plantarum RJF4. LWT-Food Sci. Technol. 2015, 64, 1179–1186. [Google Scholar] [CrossRef]
- Yılmaz, T.; Şimşek, Ö. Potential Health Benefits of Ropy Exopolysaccharides Produced by Lactobacillus plantarum. Molecules 2020, 25, 3293. [Google Scholar] [CrossRef]
- Jiang, Y.; Jiang, X.; Wang, P.; Mou, H.; Hu, X.; Liu, S. The Antitumor and Antioxidative Activities of Polysaccharides Isolated from Isaria farinosa B05. Microbiol. Res. 2008, 163, 424–430. [Google Scholar] [CrossRef]
- Pan, D.; Mei, X. Antioxidant Activity of an Exopolysaccharide Purified from Lactococcus lactis subsp. lactis 12. Carbohydr. Polym. 2010, 80, 908–914. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, X.; Jiang, Y.; Zhao, W.; Guo, T.; Cao, Y.; Teng, J.; Hao, X.; Zhao, J.; Yang, Z. Antioxidant Status and Gut Microbiota Change in an Aging Mouse Model as Influenced by Exopolysaccharide Produced by Lactobacillus plantarum YW11 Isolated from Tibetan Kefir. J. Dairy Sci. 2017, 100, 6025–6041. [Google Scholar] [CrossRef]
- Li, B.; Du, P.; Smith, E.E.; Wang, S.; Jiao, Y.; Guo, L.; Huo, G.; Liu, F. In Vitro and in Vivo Evaluation of an Exopolysaccharide Produced by Lactobacillus helveticus KLDS1.8701 for the Alleviative Effect on Oxidative Stress. Food Funct. 2019, 10, 1707–1717. [Google Scholar] [CrossRef] [PubMed]
- Tok, E.; Aslim, B. Cholesterol Removal by Some Lactic Acid Bacteria That Can Be Used as Probiotic. Microbiol. Immunol. 2010, 54, 257–264. [Google Scholar] [CrossRef]
- Costabile, A.; Buttarazzi, I.; Kolida, S.; Quercia, S.; Baldini, J.; Swann, J.R.; Brigidi, P.; Gibson, G.R. An in Vivo Assessment of the Cholesterol-Lowering Efficacy of Lactobacillus plantarum ECGC 13110402 in Normal to Mildly Hypercholesterolaemic Adults. PLoS ONE 2017, 12, e0187964. [Google Scholar] [CrossRef]
- Bhat, B.; Bajaj, B.K. Hypocholesterolemic Potential and Bioactivity Spectrum of an Exopolysaccharide from a Probiotic Isolate Lactobacillus paracasei M7. Bioact. Carbohydr. Diet. Fibre 2019, 19, 100191. [Google Scholar] [CrossRef]
- Nakajima, H.; Suzuki, Y.; Hirota, T. Cholesterol Lowering Activity of Ropy Fermented Milk. J. Food Sci. 1992, 57, 1327–1329. [Google Scholar] [CrossRef]
- Ai, L.; Zhang, H.; Guo, B.; Chen, W.; Wu, Z.; Wu, Y. Preparation, Partial Characterization and Bioactivity of Exopolysaccharides from Lactobacillus casei LC2W. Carbohydr. Polym. 2008, 74, 353–357. [Google Scholar] [CrossRef]
- Lim, J.; Kale, M.; Kim, D.H.; Kim, H.S.; Chon, J.W.; Seo, K.H.; Lee, H.G.; Yokoyama, W.; Kim, H. Antiobesity Effect of Exopolysaccharides Isolated from Kefir Grains. J. Agric. Food Chem. 2017, 65, 10011–10019. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, N.; Xi, A.; Ahmed, Z.; Zhang, B.; Bai, X. Effects of Lactobacillus plantarum MA2 Isolated from Tibet Kefir on Lipid Metabolism and Intestinal Microflora of Rats Fed on High-Cholesterol Diet. Appl. Microbiol. Biotechnol. 2009, 84, 341–347. [Google Scholar] [CrossRef]
- Akalin, A.S.; Gönç, S.; Düzel, S. Influence of Yogurt and Acidophilus Yogurt on Serum Cholesterol Levels in Mice. J. Dairy Sci. 1997, 80, 2721–2725. [Google Scholar] [CrossRef]
- Liu, J.-R.; Wang, S.-Y.; Chen, M.-J.; Chen, H.-L.; Yueh, P.-Y.; Lin, C.-W. Hypocholesterolaemic Effects of Milk-Kefir and Soyamilk-Kefir in Cholesterol-Fed Hamsters. Br. J. Nutr. 2006, 95, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Gunness, P.; Gidley, M.J. Mechanisms Underlying the Cholesterol-Lowering Properties of Soluble Dietary Fibre Polysaccharides. Food Funct. 2010, 1, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Lye, H.S.; Rusul, G.; Liong, M.T. Removal of Cholesterol by Lactobacilli via Incorporation and Conversion to Coprostanol. J. Dairy Sci. 2010, 93, 1383–1392. [Google Scholar] [CrossRef] [Green Version]
- Prete, R.; Long, S.L.; Gallardo, A.L.; Gahan, C.G.; Corsetti, A.; Joyce, S.A. Beneficial Bile Acid Metabolism from Lactobacillus plantarum of Food Origin. Sci. Rep. 2020, 10, 1165. [Google Scholar] [CrossRef] [Green Version]
- Jeong, D.; Kim, D.H.; Kang, I.B.; Kim, H.; Song, K.Y.; Kim, H.S.; Seo, K.H. Characterization and Antibacterial Activity of a Novel Exopolysaccharide Produced by Lactobacillus kefiranofaciens DN1 Isolated from Kefir. Food Control 2017, 78, 436–442. [Google Scholar] [CrossRef]
- Ayyash, M.; Abu-Jdayil, B.; Itsaranuwat, P.; Galiwango, E.; Tamiello-Rosa, C.; Abdullah, H.; Esposito, G.; Hunashal, Y.; Obaid, R.S.; Hamed, F. Characterization, Bioactivities, and Rheological Properties of Exopolysaccharide Produced by Novel Probiotic Lactobacillus plantarum C70 Isolated from Camel Milk. Int. J. Biol. Macromol. 2020, 144, 938–946. [Google Scholar] [CrossRef]
- Dertli, E.; Colquhoun, I.J.; Gunning, A.P.; Bongaerts, R.J.; Le Gall, G.; Bonev, B.B.; Mayer, M.J.; Narbad, A. Structure and Biosynthesis of Two Exopolysaccharides Produced by Lactobacillus johnsonii FI9785. J. Biol. Chem. 2013, 288, 31938–31951. [Google Scholar] [CrossRef] [Green Version]
- Spanò, A.; Laganà, P.; Visalli, G.; Maugeri, T.L.; Gugliandolo, C. In Vitro Antibiofilm Activity of an Exopolysaccharide from the Marine Thermophilic Bacillus licheniformis T14. Curr. Microbiol. 2016, 72, 518–528. [Google Scholar] [CrossRef]
- Xing, K.; Chen, X.G.; Kong, M.; Liu, C.S.; Cha, D.S.; Park, H.J. Effect of Oleoyl-Chitosan Nanoparticles as a Novel Antibacterial Dispersion System on Viability, Membrane Permeability and Cell Morphology of Escherichia Coli and Staphylococcus aureus. Carbohydr. Polym. 2009, 76, 17–22. [Google Scholar] [CrossRef]
- Rajoka, M.S.R.; Jin, M.; Haobin, Z.; Li, Q.; Shao, D.; Jiang, C.; Huang, Q.; Yang, H.; Shi, J.; Hussain, N. Functional Characterization and Biotechnological Potential of Exopolysaccharide Produced by Lactobacillus rhamnosus Strains Isolated from Human Breast Milk. LWT-Food Sci. Technol. 2018, 89, 638–647. [Google Scholar] [CrossRef]
- Sarikaya, H.; Aslim, B.; Yuksekdag, Z. Assessment of Anti-Biofilm Activity and Bifidogenic Growth Stimulator (BGS) Effect of Lyophilized Exopolysaccharides (l-EPSs) from Lactobacilli Strains. Int. J. Food Prop. 2017, 20, 362–371. [Google Scholar] [CrossRef] [Green Version]
- Abid, Y.; Casillo, A.; Gharsallah, H.; Joulak, I.; Lanzetta, R.; Corsaro, M.M.; Attia, H.; Azabou, S. Production and Structural Characterization of Exopolysaccharides from Newly Isolated Probiotic Lactic Acid Bacteria. Int. J. Biol. Macromol. 2018, 108, 719–728. [Google Scholar] [CrossRef]
- Rosca, I.; Petrovici, A.R.; Peptanariu, D.; Nicolescu, A.; Dodi, G.; Avadanei, M.; Ivanov, I.C.; Bostanaru, A.C.; Mares, M.; Ciolacu, D. Biosynthesis of Dextran by Weissella confusa and Its In Vitro Functional Characteristics. Int. J. Biol. Macromol. 2018, 107, 1765–1772. [Google Scholar] [CrossRef]
- Kanmani, P.; Suganya, K.; Satish Kumar, R.; Yuvaraj, N.; Pattukumar, V.; Paari, K.A.; Arul, V. Synthesis and Functional Characterization of Antibiofilm Exopolysaccharide Produced by Enterococcus faecium Mc13 Isolated from the Gut of Fish. Appl. Biochem. Biotechnol. 2013, 169, 1001–1015. [Google Scholar] [CrossRef]
- Jiang, P.; Li, J.; Han, F.; Duan, G.; Lu, X.; Gu, Y.; Yu, W. Antibiofilm Activity of an Exopolysaccharide from Marine Bacterium Vibrio sp. QY101. PLoS ONE 2011, 6, e18514. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Castillo, V.; Marcial, G.; Albarracín, L.; Tomokiyo, M.; Clua, P.; Takahashi, H.; Kitazawa, H.; Garcia-Cancino, A.; Villena, J. The Exopolysaccharide of Lactobacillus fermentum UCO-979C Is Partially Involved in Its Immunomodulatory Effect and Its Ability to Improve the Resistance against Helicobacter Pylori Infection. Microorganisms 2020, 8, 479. [Google Scholar] [CrossRef] [Green Version]
- Korcz, E.; Kerényi, Z.; Varga, L. Dietary Fibers, Prebiotics, and Exopolysaccharides Produced by Lactic Acid Bacteria: Potential Health Benefits with Special Regard to Cholesterol-Lowering Effects. Food Funct. 2018, 9, 3057–3068. [Google Scholar] [CrossRef]
- Gagliardi, A.; Totino, V.; Cacciotti, F.; Iebba, V.; Neroni, B.; Bonfiglio, G.; Trancassini, M.; Passariello, C.; Pantanella, F.; Schippa, S. Rebuilding the Gut Microbiota Ecosystem. Int. J. Environ. Res. Public Health 2018, 15, 1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, G.R.; Roberfroid, M.B. Dietary Modulation of the Human Colonic Microbiota: Introducing the Concept of Prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Salazar, N.; Gueimonde, M.; de los Reyes-Gavilán, C.G.; Ruas-Madiedo, P. Exopolysaccharides Produced by Lactic Acid Bacteria and Bifidobacteria as Fermentable Substrates by the Intestinal Microbiota. Crit. Rev. Food Sci. Nutr. 2016, 56, 1440–1453. [Google Scholar] [CrossRef]
- Dal Bello, F.; Walter, J.; Hertel, C.; Hammes, W.P. In Vitro Study of Prebiotic Properties of Levan-Type Exopolysaccharides from Lactobacilli and Non-Digestible Carbohydrates Using Denaturing Gradient Gel Electrophoresis. Syst. Appl. Microbiol. 2001, 24, 232–237. [Google Scholar] [CrossRef]
- Hongpattarakere, T.; Cherntong, N.; Wichienchot, S.; Kolida, S.; Rastall, R.A. In Vitro Prebiotic Evaluation of Exopolysaccharides Produced by Marine Isolated Lactic Acid Bacteria. Carbohydr. Polym. 2012, 87, 846–852. [Google Scholar] [CrossRef]
- Das, D.; Baruah, R.; Goyal, A. A Food Additive with Prebiotic Properties of an α-d-Glucan from Lactobacillus plantarum DM5. Int. J. Biol. Macromol. 2014, 69, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Zhou, J.; Xu, Q.; Dong, M.; Fan, X.; Rui, X.; Zhang, Q.; Chen, X.; Jiang, M.; Wu, J.; et al. In Vitro Digestion and Fermentation of Released Exopolysaccharides (r-EPS) from Lactobacillus delbrueckii ssp. bulgaricus SRFM-1. Carbohydr. Polym. 2020, 230, 115593. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Han, S.; Zhou, J.; Xu, Q.; Dong, M.; Fan, X.; Rui, X.; Zhang, Q.; Chen, X.; Jiang, M.; et al. Selective Fermentation of Lactobacillus delbrueckii ssp. bulgaricus SRFM-1 Derived Exopolysaccharide by Lactobacillus and Streptococcus Strains Revealed Prebiotic Properties. J. Funct. Foods 2020, 69, 103952. [Google Scholar] [CrossRef]
- Russo, P.; López, P.; Capozzi, V.; de Palencia, P.F.; Dueñas, M.T.; Spano, G.; Fiocco, D. Beta-Glucans Improve Growth, Viability and Colonization of Probiotic Microorganisms. Int. J. Mol. Sci. 2012, 13, 6026–6039. [Google Scholar] [CrossRef] [Green Version]
- Lindström, C.; Holst, O.; Nilsson, L.; Öste, R.; Andersson, K.E. Effects of Pediococcus parvulus 2.6 and Its Exopolysaccharide on Plasma Cholesterol Levels and Inflammatory Markers in Mice. AMB Express 2012, 2, 66. [Google Scholar] [CrossRef] [Green Version]
- Lambo-Fodje, A.M.; Öste, R.; Nyman, M.E.G.-L. Short-Chain Fatty Acid Formation in the Hindgut of Rats Fed Native and Fermented Oat Fibre Concentrates. Br. J. Nutr. 2006, 96, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Fraher, M.H.; O’Toole, P.W.; Quigley, E.M.M. Techniques Used to Characterize the Gut Microbiota: A Guide for the Clinician. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 312–322. [Google Scholar] [CrossRef]
- Hyötyläinen, T. Novel Methodologies in Metabolic Profiling with a Focus on Molecular Diagnostic Applications. Expert Rev. Mol. Diagn. 2012, 12, 527–538. [Google Scholar] [CrossRef]
- Xu, Y.; Cui, Y.; Yue, F.; Liu, L.; Shan, Y.; Liu, B.; Zhou, Y.; Lü, X. Exopolysaccharides Produced by Lactic Acid Bacteria and Bifidobacteria: Structures, Physiochemical Functions and Applications in the Food Industry. Food Hydrocoll. 2019, 94, 475–499. [Google Scholar] [CrossRef]
- Di Cagno, R.; De Pasquale, I.; De Angelis, M.; Buchin, S.; Rizzello, C.G.; Gobbetti, M. Use of Microparticulated Whey Protein Concentrate, Exopolysaccharide-Producing Streptococcus thermophilus, and Adjunct Cultures for Making Low-Fat Italian Caciotta-Type Cheese. J. Dairy Sci. 2014, 97, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Purohit, D.H.; Hassan, A.N.; Bhatia, E.; Zhang, X.; Dwivedi, C. Rheological, Sensorial, and Chemopreventive Properties of Milk Fermented with Exopolysaccharide-Producing Lactic Cultures. J. Dairy Sci. 2009, 92, 847–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amatayakul, T.; Halmos, A.L.; Sherkat, F.; Shah, N.P. Physical Characteristics of Yoghurts Made Using Exopolysaccharide-Producing Starter Cultures and Varying Casein to Whey Protein Ratios. Int. Dairy J. 2006, 16, 40–51. [Google Scholar] [CrossRef]
- Han, X.; Yang, Z.; Jing, X.; Yu, P.; Zhang, Y.; Yi, H.; Zhang, L. Improvement of the Texture of Yogurt by Use of Exopolysaccharide Producing Lactic Acid Bacteria. BioMed Res. Int. 2016, 2016, 7945675. [Google Scholar] [CrossRef] [PubMed]
- Petry, S.; Furlan, S.; Waghorne, E.; Saulnier, L.; Cerning, J.; Maguin, E. Comparison of the Thickening Properties of Four Lactobacillus delbrueckii subsp. bulgaricus Strains and Physicochemical Characterization of Their Exopolysaccharides. FEMS Microbiol. Lett. 2003, 221, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Ayyash, M.; Abu-Jdayil, B.; Hamed, F.; Shaker, R. Rheological, Textural, Microstructural and Sensory Impact of Exopolysaccharide-Producing Lactobacillus plantarum Isolated from Camel Milk on Low-Fat Akawi Cheese. LW-Food Sci. Technol. 2018, 87, 423–431. [Google Scholar] [CrossRef]
- Ale, E.C.; Perezlindo, M.J.; Pavón, Y.; Peralta, G.H.; Costa, S.; Sabbag, N.; Bergamini, C.; Reinheimer, J.A.; Binetti, A.G. Technological, Rheological and Sensory Characterizations of a Yogurt Containing an Exopolysaccharide Extract from Lactobacillus fermentum Lf2, a New Food Additive. Food Res. Int. 2016, 90, 259–267. [Google Scholar] [CrossRef] [PubMed]
- London, L.E.E.; Chaurin, V.; Auty, M.A.E.; Fenelon, M.A.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C. Use of Lactobacillus mucosae DPC 6426, an Exopolysaccharide-Producing Strain, Positively Influences the Techno-Functional Properties of Yoghurt. Int. Dairy J. 2015, 40, 33–38. [Google Scholar] [CrossRef]
- Prasanna, P.H.P.; Grandison, A.S.; Charalampopoulos, D. Microbiological, Chemical and Rheological Properties of Low Fat Set Yoghurt Produced with Exopolysaccharide (EPS) Producing Bifidobacterium Strains. Food Res. Int. 2013, 51, 15–22. [Google Scholar] [CrossRef]
- Medrano, M.; Hamet, M.F.; Abraham, A.G.; Pérez, P.F. Kefiran Protects Caco-2 Cells from Cytopathic Effects Induced by Bacillus cereus Infection. Antonie van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2009, 96, 505–513. [Google Scholar] [CrossRef]
- Escalante, A.; Elena Rodríguez, M.; Martínez, A.; López-Munguía, A.; Bolívar, F.; Gosset, G. Characterization of Bacterial Diversity in Pulque, a Traditional Mexican Alcoholic Fermented Beverage, as Determined by 16S RDNA Analysis. FEMS Microbiol. Lett. 2004, 235, 273–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behare, P.; Singh, R.; Singh, R.P. Exopolysaccharide-Producing Mesophilic Lactic Cultures for Preparation of Fat-Free Dahi-an Indian Fermented Milk. J. Dairy Res. 2009, 76, 90–97. [Google Scholar] [CrossRef]
- Hassan, A.N. ADSA Foundation Scholar Award: Possibilities and Challenges of Exopolysaccharide-Producing Lactic Cultures in Dairy Foods. J. Dairy Sci. 2008, 91, 1282–1298. [Google Scholar] [CrossRef]
- Costa, N.E.; Hannon, J.A.; Guinee, T.P.; Auty, M.A.E.; McSweeney, P.L.H.; Beresford, T.P. Effect of Exopolysaccharide Produced by Isogenic Strains of Lactococcus lactis on Half-Fat Cheddar Cheese. J. Dairy Sci. 2010, 93, 3469–3486. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.N.; Corredig, M.; Frank, J.F.; Elsoda, M. Microstructure and Rheology of an Acid-Coagulated Cheese (Karish) Made with an Exopolysaccharide-Producing Streptococcus thermophilus Strain and Its Exopolysaccharide Non-Producing Genetic Variant. J. Dairy Res. 2004, 71, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Characklis, W.G.; Wilderer, P.A. The Structure and Function of Biofilms; Wiley: Chichester, UK, 1989. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| EPS | Producers | Biological Properties |
|---|---|---|
| HoPS | ||
| Dextran (glucose) | Leuconostoc mesenteroides, Lactobacillus reuteri, Lacticaseibacillus casei, Latilactobacillus sakei, Limosilactobacillus fermentum, Lentilactobacillus parabuchneri | Food industry: emulsifier and stabilizer, improvement of softness, crumb texture, loaf volume in bakery products, improvement of moisture retention and viscosity in confectionary |
| Reuteran (glucose) | Lactobacillus reuteri | Bakery industry |
| Levan (fructose) | Bacillus subtilis, Streptococcus salivarius, Streptococcus mutans | Health benefits: prebiotic activity, antitumor property, hypocholesterolemic agent Food industry: bio-thickener |
| Inulin-type (fructose) | Streptococcus mutans, Lactobacillus reuteri | Health benefits: prebiotic activity Food industry: sugar and fat replacer, texture modifier in low-fat dairy products enhancer of creaminess |
| Alternan (glucose) | L. mesenteroides, Leuconostoc citreum | Food industry: Sweetener in confectionary, stabilizer, emulsifier and prebiotics |
| Curdlan (glucose) | Alcaligenes faecalis, Rhizobium meliloti, Agrobacterium radiobacter | Starter culture, gelling agent, immoblization matrix |
| HePS | ||
| Glucose and galactose | Lactiplantibacillus plantarum, Streptococcus thermophilus, Enterococcus faecium | Health benefits: immunomodulatory activity |
| Arabinose, mannose, glucose and galactose | Lpb. plantarum | Food industry: improvement of texture and rheological properties of various food stuffs |
| Glucose, arabinose, galactose, mannose and xylose | Bacillus tequilensis | Food industry: stabilizer and thickener |
| Glucose, mannose, galactose, rhamnose, and a small fraction fucose | Lactobacillus gasseri | Food industry: antioxidant agent, viscosifying agent and antimicrobial agent |
| Arabinose, rhamnose, fucose, xylose, mannose, fructose, galactose and glucose | Lpb. plantarum | Food industry: natural antioxidant or functional additive |
| Genus | Strains | EPS Yield (mg/L) | Culture Media | Carbon Source in Media | Monosaccharide Composition | Reference |
|---|---|---|---|---|---|---|
| Streptococcus | ||||||
| S. thermophilus DGCC7919 | 404 | Whey permeate | Lactose | Glucose, Galactose, Rhamnose, Mannose | [33] | |
| S. thermophilus ASCC 1275 | 163–430 | M17 | Glucose or lactose or sucrose | Glucose, Galactose | [34] | |
| S. thermophilus S-3 | 100 | Skim milk | Lactose | Glucose, Galactose | [35] | |
| S. thermophilus CC30 | 1950 | Skim milk | Lactose | Glucose, Galactose | [36] | |
| S. thermophilus 05-34 | 55–120 | Skim milk | Glucose or Galactose or Lactose or Sucrose or Fructose | Glucose, Galactose | [37] | |
| S. thermophilus GST-6 | Not reported | Skim milk | Lactose | Glucose, Galactose | [38] | |
| S. thermophilus ST1 | 136 | Skim milk | Lactose and sucrose | Glucose, Galactose | [39] | |
| S. thermophilus CRL804 | 166 | Skim milk | Lactose | Galactose, Rhamnose | [12] | |
| S. thermophilus SY89, SY102, IMDO1, IMDO2, IMDO3, NCFB 859 | Not reported | Skim milk | Lactose | Glucose, Galactose | [40] | |
| S. thermophilus NCFB 2393 | 300 | Skim milk | Lactose | Glucose, Galactose, Rhamnose | [41] | |
| Lactobacillus | ||||||
| Lb. delbrueckii subsp. bulgaricus | 5570.34–5910.62 | Modified Skim milk | Glucose | Glucose, Galactose, Rhamnose, Ribose, Mannose, Xylose, Arabinose, Fructose | [42] | |
| Lb. delbrueckii subsp. bulgaricus 147 | 960 | Skim milk | Lactose | Glucose, Galactose, Rhamnose, Ribose, Mannose | [43] | |
| Lb. delbrueckii subsp. bulgaricus 2214 | 1880 | Skim milk | Lactose | Glucose, Rhamnose, Mannose | [43] | |
| Lb. delbrueckii subsp. bulgaricus B3 | 449 | MRS | Glucose | Glucose, Fructose, Mannose | [27] | |
| Lb. delbrueckii subsp. bulgaricus SRFM-1 | 141.63 | Milk | Lactose | Glucose, Galactose | [44] | |
| Lb. delbrueckii subsp. bulgaricus OLL1073R-1 | 1546 | Skim milk | Lactose | Glucose, Galactose | [45] | |
| Lb. delbrueckii subsp. bulgaricus | 190–740 | Modified MRS | Lactose and Glucose | Not reported | [46] | |
| Lb. delbrueckii subsp. bulgaricus CRL 852, 865, 874 | 24–150 | Skim milk | Lactose | Glucose, Galactose, Rhamnose | [12] | |
| Lb. delbrueckii subsp. bulgaricus LY03 | Not reported | Skim milk | Lactose | Glucose, Galactose, Rhamnose | [40] | |
| Lb. helveticus LZ-R-5 | 128 | Milk | Lactose | Glucose, Galactose | [47] | |
| Lactiplantibacillus | ||||||
| Lpb. plantarum subsp. plantarum JLAU103 | 75 | Modified MRS | Sorbitol | Glucose, Galactose, Rhamnose, Mannose, Xylose, Arabinose, Fructose, Fucose | [48] | |
| Lpb. plantarum subsp. plantarum GD2 | 397 | MRS | Glucose | Glucose, Mannose, Arabinose | [27] | |
| Lpb. plantarum subsp. plantarum JLK0142 | Not reported | Modified MRS | Sorbitol | Glucose, Galactose | [49] | |
| Lpb. plantarum subsp. plantarum C7 | 198–265 | Modified MRS | Sucrose | Glucose, Mannose | [50] | |
| Limosilactobacillus | ||||||
| Lim. reuteri L26 and Lim. reuteri DSM 17938 | 4300–5000 | Modified MRS | Sucrose | Glucose | [32] | |
| Lim. fermentum YL-11 | 84.5 | Skim milk | Lactose | Glucose, Galactose, Mannose, Arabinose | [26] | |
| Lacticaseibacillus | ||||||
| Lcb. rhamnosus E9 | 298 | MRS | Glucose | Glucose, Mannose, Arabinose | [27] | |
| Lcb. rhamnosus RW-9595M | 931–1275 | MRS | Glucose | Glucose, Galactose, Rhamnose | [51] | |
| Lcb. rhamnosus R | 438–601 | MRS | Glucose or Lactose | Glucose, Galactose, Rhamnose | [51] | |
| Levilactobacillus | ||||||
| Lev. brevis LB63 | 347 | MRS | Glucose | Glucose, Mannose, Arabinose | [27] | |
| Lactococcus | ||||||
| Lact. lactis LL-2A | 354 | Whey permeate | Lactose | Glucose, Galactose, Mannose | [33] | |
| Lact. lactis SLT10 | 280–336 | Modified MRS | Sucrose | Glucose, Mannose, Rhamnose | [50] | |
| Lact. lactis subsp. cremoris JFR1 | Not reported | Skim milk | Lactose | Glucose, Galactose, Rhamnose | [52] | |
| Lact. lactis subsp. cremoris 352 | 204 | Skim milk | Lactose | Glucose, Galactose | [19] |
| Main Influencing Factors | Example | Reference |
|---|---|---|
| Microbial factors | ||
| EPS producers | Species/strains | [8,12,24,35,53] |
| Bacterial growth | Exponential/stationary phase | [38,60,68,82,83] |
| Microbial interactions | Synergistic/antagonist effects in bacterial mixture | [71,84] |
| Medium composition | ||
| Carbon source | Glucose/Sucrose/Lactose etc. | [34,37,39,55,56,60,64,82] |
| Nitrogen source | Organic/inorganic nitrogen | [37,57,60,68,82] |
| Physicochemical parameters | ||
| pH | pH of medium and its variations | [37,54,57,60,68,82] |
| Temperature | Optimal/sub-optimal temperature | [37,39,57,59,60,64,67,68,69,82] |
| Oxygen rate | Aerobiosis/anaerobiosis | [24,60,67,85] |
| Incubation time | Prolonged incubation time | [37,39,55,57,64,66,68,82] |
| EPS-Producers | Foods | Main Effects | Reference |
|---|---|---|---|
| S. thermophilus | Yogurt drink (Ayran) | Increased apparent viscosity | [107] |
| S. thermophilus | Low-fat Italian Cacciotta type cheese | Improved taste, flavor and chew ability | [171] |
| S. thermophilus ST-5581, ST-4239 and ST-PH Lact.lactis subsp. cremoris JFR1 | Fermented milk | Low syneresis, increased viscosity, lowered incidence of colon tumor and activity of cyclooxygenase-2 enzyme | [172] |
| S.thermophilus ASCC 1275 | Set and stirred yogurt | Decreased firmness and syneresis | [173] |
| S. thermophilus zlw TM11 | Yogurt | Improved yogurt texture and lower separation of whey | [174] |
| Lb. delbrueckii subsp. bulgaricus 147 and 2214 | Fermented milk | Improved viscosity, low syneresis and increased water holding capacity | [43] |
| Lb. delbrueckii subsp. bulgaricus CNRZ 1187 and 416 | Fermented milk | Improved viscosity | [175] |
| Lpb. plantarum KX881772 and KX881779 | Low-fat Akawi cheese | Improved water holding capacity, improved elastic structure, reduced hardness and adhesiveness, higher scores in appearance and overall quality | [176] |
| Lim. fermentum Lf2 | Non-fat yogurt | Creamy consistency, increased hardness, improved water holding capacity and low syneresis | [177] |
| Lim. mucosae DPC 6426 | Low-fat yogurt | Improved viscosity and reduced syneresis | [178] |
| Lcb. rhamnosus JAAS8 | Yogurt | Increased apparent viscosity and improved water holding capacity | [83] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prete, R.; Alam, M.K.; Perpetuini, G.; Perla, C.; Pittia, P.; Corsetti, A. Lactic Acid Bacteria Exopolysaccharides Producers: A Sustainable Tool for Functional Foods. Foods 2021, 10, 1653. https://doi.org/10.3390/foods10071653
Prete R, Alam MK, Perpetuini G, Perla C, Pittia P, Corsetti A. Lactic Acid Bacteria Exopolysaccharides Producers: A Sustainable Tool for Functional Foods. Foods. 2021; 10(7):1653. https://doi.org/10.3390/foods10071653
Chicago/Turabian StylePrete, Roberta, Mohammad Khairul Alam, Giorgia Perpetuini, Carlo Perla, Paola Pittia, and Aldo Corsetti. 2021. "Lactic Acid Bacteria Exopolysaccharides Producers: A Sustainable Tool for Functional Foods" Foods 10, no. 7: 1653. https://doi.org/10.3390/foods10071653
APA StylePrete, R., Alam, M. K., Perpetuini, G., Perla, C., Pittia, P., & Corsetti, A. (2021). Lactic Acid Bacteria Exopolysaccharides Producers: A Sustainable Tool for Functional Foods. Foods, 10(7), 1653. https://doi.org/10.3390/foods10071653