
Antifungal Efficacy of Redox-Active Natamycin against Some Foodborne Fungi—Comparison with Aspergillus fumigatus

 ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Chemicals
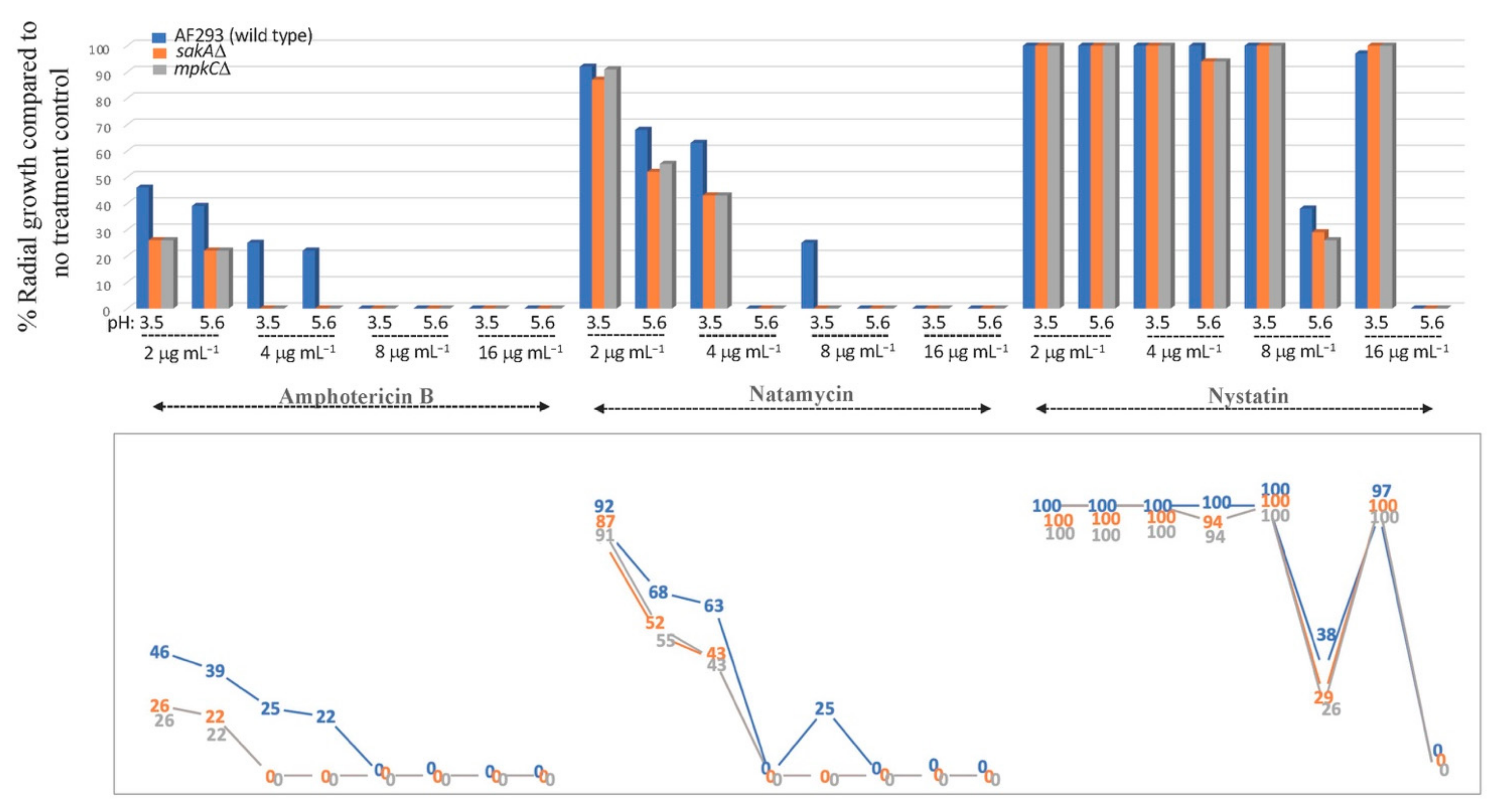
2.3. Overcoming FLU Tolerance of A. fumigatus MAPK Mutants (sakAΔ, mpkCΔ) by NAT
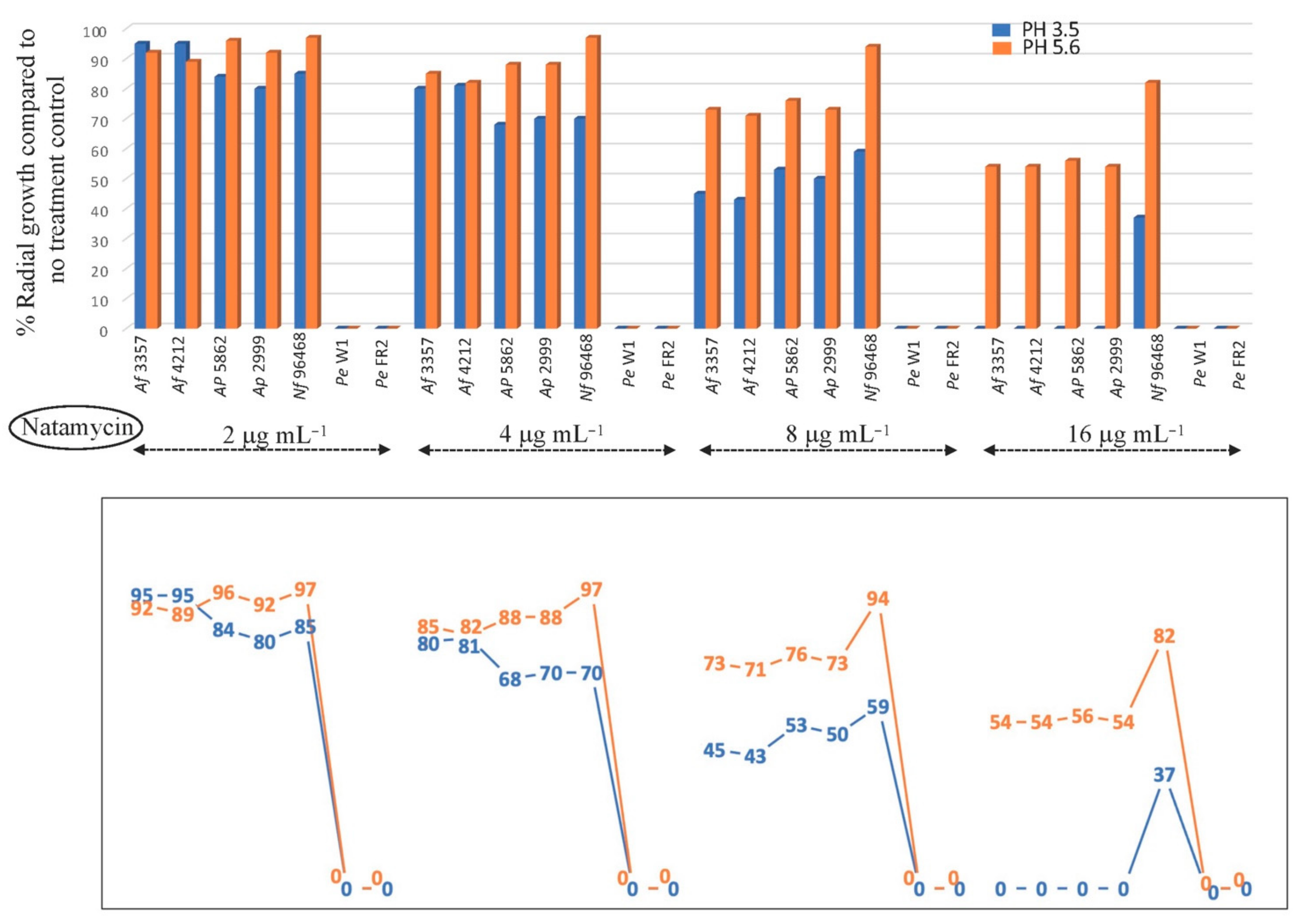
2.4. Effect of pH (3.5 and 5.6) on the Antifungal Efficacy of NAT, AMB, or NYS
2.5. Antifungal Synergism (Chemosensitization) between NAT and KA
2.6. Effect of Food Ingredients on the Efficacy of NAT Tested on Organic Fruit Juice Agar
2.7. Statistical Analysis
3. Results and Discussion
3.1. Redox Activity of NAT: Overcoming FLU Tolerance of A. fumigatus Antioxidant Signaling Mutants
3.2. Differential NAT Susceptibility between Human Pathogen (A. fumigatus) and Foodborne Fungal Contaminants (A. flavus, A. parasiticus, P. expansum, N. fischeri) under Different pHs: High (pH 3.5) versus Low (pH 5.6) Acidity Conditions
3.2.1. Fungal Susceptibility to Polyene Drugs (NAT, AMB, NYS): PDA Test
3.2.2. Fungal Susceptibility to Polyene Drugs (NAT, AMB, NYS): Food Matrices (Organic Fruit Juices) Test
3.3. Enhanced Antifungal Activity of Polyenes with KA in Filamentous Fungi
4. Summary and Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- King, L.J. A8 One Health and Food Safety. In Improving Food Safety Through a One Health Approach: Workshop Summary; Institute of Medicine (US); National Academies Press (US): Washington, DC, USA, 2012. Available online: https://www.ncbi.nlm.nih.gov/books/NBK114498/ (accessed on 26 April 2021).
- Centers for Disease Control and Prevention. Aspergillosis. Available online: https://www.cdc.gov/fungal/diseases/aspergillosis/index.html (accessed on 21 April 2021).
- Maschmeyer, G.; Haas, A.; Cornely, O.A. Invasive aspergillosis: Epidemiology, diagnosis and management in immunocompromised patients. Drugs 2007, 67, 1567–1601. [Google Scholar] [CrossRef]
- Rivero-Menendez, O.; Alastruey-Izquierdo, A.; Mellado, E.; Cuenca-Estrella, M. Triazole Resistance in Aspergillus spp.: A Worldwide Problem? J. Fungi 2016, 2, 21. [Google Scholar] [CrossRef] [Green Version]
- Benedict, K.; Chiller, T.M.; Mody, R.K. Invasive Fungal Infections Acquired from Contaminated Food or Nutritional Supplements: A Review of the Literature. Foodborne Pathog. Dis. 2016, 13, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Perfect, J.R. The antifungal pipeline: A reality check. Nat. Rev. Drug Discov. 2017, 16, 603–616. [Google Scholar] [CrossRef] [Green Version]
- Steinbach, W.J.; Stevens, D.A.; Denning, D.W. Combination and Sequential Antifungal Therapy for Invasive Aspergillosis: Review of Published In Vitro and In Vivo Interactions and 6281 Clinical Cases from 1966 to 2001. Clin. Infect. Dis. 2003, 37, S188–S244. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Cheng, L.W.; Chan, K.L.; Tam, C.C.; Mahoney, N.; Friedman, M.; Shilman, M.M.; Land, K.M. Antifungal Drug Repurposing. Antibiotics 2020, 9, 812. [Google Scholar] [CrossRef]
- Zhang, J.; Li, L.; Lv, Q.; Yan, L.; Wang, Y.; Jiang, Y. The Fungal CYP51s: Their Functions, Structures, Related Drug Resistance, and Inhibitors. Front. Microbiol. 2019, 10, 691. [Google Scholar] [CrossRef]
- Lakhani, P.; Patil, A.; Majumdar, S. Challenges in the Polyene- and Azole-Based Pharmacotherapy of Ocular Fungal Infections. J. Ocul. Pharmacol. Ther. 2019, 35, 6–22. [Google Scholar] [CrossRef]
- Vazquez, J.A.; Arganoza, M.T.; Boikov, D.; Yoon, S.; Sobel, J.D.; Akins, R.A. Stable Phenotypic Resistance of Candida Species to Amphotericin B Conferred by Preexposure to Subinhibitory Levels of Azoles. J. Clin. Microbiol. 1998, 36, 2690–2695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajendran, R.; Mowat, E.; Jones, B.; Williams, C.; Ramage, G. Prior in vitro exposure to voriconazole confers resistance to amphotericin B in Aspergillus fumigatus biofilms. Int. J. Antimicrob. Agents 2015, 46, 342–345. [Google Scholar] [CrossRef] [Green Version]
- Lamoth, F.; Alexander, B.D.; Juvvadi, P.R.; Steinbach, W.J. Antifungal activity of compounds targeting the Hsp90-calcineurin pathway against various mould species. J. Antimicrob. Chemother. 2015, 70, 1408–1411. [Google Scholar] [CrossRef] [Green Version]
- Delattin, N.; Cammue, B.P.; Thevissen, K. Reactive oxygen species-inducing antifungal agents and their activity against fungal biofilms. Future Med. Chem. 2014, 6, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Chan, K.; Faria, N.C.G.; Martins, M.D.L.; Campbell, B. Targeting the Oxidative Stress Response System of Fungi with Redox-Potent Chemosensitizing Agents. Front. Microbiol. 2012, 3, 88. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Faria, N.; Martins, M.; Chan, K.; Campbell, B. Enhancement of Antimycotic Activity of Amphotericin B by Targeting the Oxidative Stress Response of Candida and Cryptococcus with Natural Dihydroxybenzaldehydes. Front. Microbiol. 2012, 3, 261. [Google Scholar] [CrossRef] [Green Version]
- Bowyer, P.; Denning, D.W. Environmental fungicides and triazole resistance in Aspergillus. Pest Manag. Sci. 2014, 70, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Sun, Y.; Chen, W.; Liu, W.; Wan, Z.; Bu, D.; Li, R. The T788G Mutation in the cyp51C Gene Confers Voriconazole Resistance in Aspergillus flavus Causing Aspergillosis. Antimicrob. Agents Chemother. 2012, 56, 2598–2603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, B.C.; Molyneux, R.J.; Schatzki, T.F. Current Research on Reducing Pre- and Post-harvest Aflatoxin Contamination of U.S. Almond, Pistachio, and Walnut. J. Toxicol. Toxin Rev. 2003, 22, 225–266. [Google Scholar] [CrossRef]
- Doukas, E.G.; Markoglou, A.N.; Vontas, J.G.; Ziogas, B.N. Effect of DMI-resistance mechanisms on cross-resistance patterns, fitness parameters and aflatoxin production in Aspergillus parasiticus Speare. Fungal Genet. Biol. 2012, 49, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Wang, F.; Xiao, C.L. Efficacy of Natamycin Against Gray Mold of Stored Mandarin Fruit Caused by Isolates of Botrytis cinerea With Multiple Fungicide Resistance. Plant Dis. 2020, 104, 787–792. [Google Scholar] [CrossRef]
- Haack, S.E.; Ivors, K.L.; Holmes, G.J.; Förster, H.; Adaskaveg, J.E. Natamycin, a New Biofungicide for Managing Crown Rot of Strawberry Caused by QoI-Resistant Colletotrichum acutatum. Plant Dis. 2018, 102, 1687–1695. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zhao, H.; Lyu, L.; Huang, Z.; Fan, S.; Wu, W.; Li, W. Synergistic effect of natural antifungal agents for postharvest diseases of blackberry fruits. J. Sci. Food Agric. 2019, 99, 3343–3349. [Google Scholar] [CrossRef]
- Medina, A.; Jiménez, M.; Mateo, R.; Magan, N. Efficacy of natamycin for control of growth and ochratoxin A production by Aspergillus carbonarius strains under different environmental conditions. J. Appl. Microbiol. 2007, 103, 2234–2239. [Google Scholar] [CrossRef]
- Chen, D.; Forster, H.; Adaskaveg, J. Baseline Sensitivities of Major Citrus, Pome, and Stone Fruits Postharvest Pathogens to Natamycin and Estimation of the Resistance Potential in Penicillium digitatum. Plant Dis. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Forster, H.; Nguyen, K.; Adaskaveg, J. Organic Acids as Sanitizers for Natamycin and other Fungicides in Postharvest Recirculating Application Systems for Citrus. Plant Dis. 2021, in press. [Google Scholar] [CrossRef]
- Van Leeuwen, M.R.; Golovina, E.A.; Dijksterhuis, J. The polyene antimycotics nystatin and filipin disrupt the plasma membrane, whereas natamycin inhibits endocytosis in germinating conidia of Penicillium discolor. J. Appl. Microbiol. 2009, 106, 1908–1918. [Google Scholar] [CrossRef] [PubMed]
- Dalhoff, A. Does the use of antifungal agents in agriculture and food foster polyene resistance development? A reason for concern. J. Glob. Antimicrob. Resist. 2018, 13, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Rusul, G.; Marth, E.H. Growth and aflatoxin production by Aspergillus parasiticus in a medium at different pH values and with or without pimaricin. Z. Lebensm. Unters. Forsch. 1988, 187, 436–439. [Google Scholar] [CrossRef]
- Teixeira, G.H.A.; O’Keefe, S.F. Short communication: Mycosporine-like amino acids protect natamycin against photodegradation in milk exposed to fluorescent or light-emitting diode light. J. Dairy Sci. 2019, 102, 4972–4977. [Google Scholar] [CrossRef] [PubMed]
- Xue, T.; Nguyen, C.K.; Romans, A.; May, G.S. A Mitogen-Activated Protein Kinase That Senses Nitrogen Regulates Conidial Germination and Growth in Aspergillus fumigatus. Eukaryot Cell 2004, 3, 557–560. [Google Scholar] [CrossRef] [Green Version]
- Reyes, G.; Romans, A.; Nguyen, C.K.; May, G.S. Novel Mitogen-Activated Protein Kinase MpkC of Aspergillus fumigatus Is Required for Utilization of Polyalcohol Sugars. Eukaryot Cell 2006, 5, 1934–1940. [Google Scholar] [CrossRef] [Green Version]
- Li, H.X.; Xiao, C.L. Characterization of Fludioxonil-Resistant and Pyrimethanil-Resistant Phenotypes of Penicillium expansum from Apple. Phytopathology 2008, 98, 427–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Chan, K.L.; Mahoney, N.; Campbell, B.C. Antifungal activity of redox-active benzaldehydes that target cellular antioxidation. Ann. Clin. Microbiol. Antimicrob. 2011, 10, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkman, T.W. Statistics to Use. Available online: http://www.physics.csbsju.edu/stats/ (accessed on 11 May 2021).
- Beites, T.; Pires, S.D.S.; Santos, C.L.; Osório, H.; Moradas-Ferreira, P.; Mendes, M.V. Crosstalk between ROS Homeostasis and Secondary Metabolism in S. natalensis ATCC 27448: Modulation of Pimaricin Production by Intracellular ROS. PLoS ONE 2011, 6, e27472. [Google Scholar] [CrossRef] [PubMed]
- Beites, T.; Rodríguez-García, A.; Moradas-Ferreira, P.; Aparicio, J.F.; Mendes, M.V. Genome-wide analysis of the regulation of pimaricin production in Streptomyces natalensis by reactive oxygen species. Appl. Microbiol. Biotechnol. 2014, 98, 2231–2241. [Google Scholar] [CrossRef]
- Maeda, T.; Wurgler-Murphy, S.M.; Saito, H. A two-component system that regulates an osmosensing MAP kinase cascade in yeast. Nature 1994, 369, 242–245. [Google Scholar] [CrossRef]
- Kojima, K.; Takano, Y.; Yoshimi, A.; Tanaka, C.; Kikuchi, T.; Okuno, T. Fungicide activity through activation of a fungal signalling pathway. Mol. Microbiol. 2004, 53, 1785–1796. [Google Scholar] [CrossRef]
- Rep, M.; Krantz, M.; Thevelein, J.M.; Hohmann, S. The Transcriptional Response of Saccharomyces cerevisiae to Osmotic Shock: Hot1p and Msn2p/Msn4p are required for the induction of subsets of high osmolarity glycerol pathway-dependent genes. J. Biol. Chem. 2000, 275, 8290–8300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rep, M.; Proft, M.; Remize, F.; Tamás, M.; Serrano, R.; Thevelein, J.M.; Hohmann, S. The Saccharomyces cerevisiae Sko1p transcription factor mediates HOG pathway-dependent osmotic regulation of a set of genes encoding enzymes implicated in protection from oxidative damage. Mol. Microbiol. 2001, 40, 1067–1083. [Google Scholar] [CrossRef] [Green Version]
- Henriet, S.S.V.; Jans, J.; Simonetti, E.; Kwon-Chung, K.J.; Rijs, A.J.M.M.; Hermans, P.W.M.; Holland, S.M.; de Jonge, M.I.; Warris, A. Chloroquine Modulates the Fungal Immune Response in Phagocytic Cells From Patients With Chronic Granulomatous Disease. J. Infect. Dis. 2013, 207, 1932–1939. [Google Scholar] [CrossRef]
- Gsaller, F.; Furukawa, T.; Carr, P.D.; Rash, B.; Jöchl, C.; Bertuzzi, M.; Bignell, E.M.; Bromley, M.J. Mechanistic Basis of pH-Dependent 5-Flucytosine Resistance in Aspergillus fumigatus. Antimicrob. Agents Chemother. 2018, 62, e02593-17. [Google Scholar] [CrossRef] [Green Version]
- Holmquist, G.U.; Walker, H.W.; Stahr, H.M. Influence of Temperature, pH, Water Activity and Antifungal Agents on Growth of Aspergillus flavus and A. parasiticus. J. Food Sci. 1983, 48, 778–782. [Google Scholar] [CrossRef]
- Nguefack, J.; Leth, V.; Amvam Zollo, P.H.; Mathur, S.B. Evaluation of five essential oils from aromatic plants of Cameroon for controlling food spoilage and mycotoxin producing fungi. Int. J. Food Microbiol. 2004, 94, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Chaillot, J.; Tebbji, F.; García, C.; Wurtele, H.; Pelletier, R.; Sellam, A. pH-Dependant Antifungal Activity of Valproic Acid against the Human Fungal Pathogen Candida albicans. Front. Microbiol. 2017, 8, 1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badhani, B.; Sharma, N.; Kakkar, R. Gallic acid: A versatile antioxidant with promising therapeutic and industrial applications. RSC Adv. 2015, 5, 27540–27557. [Google Scholar] [CrossRef]
- Guillén, F.; Evans, C.S. Anisaldehyde and Veratraldehyde Acting as Redox Cycling Agents for H2O2 Production by Pleurotus eryngii. Appl. Environ. Microbiol. 1994, 60, 2811–2817. [Google Scholar] [CrossRef] [Green Version]
- Jacob, C. A scent of therapy: Pharmacological implications of natural products containing redox-active sulfur atoms. Nat. Prod. Rep 2006, 23, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Campbell, B.C.; Mahoney, N.; Chan, K.L.; Molyneux, R.J.; May, G.S. Enhancement of fludioxonil fungicidal activity by disrupting cellular glutathione homeostasis with 2,5-dihydroxybenzoic acid. FEMS Microbiol. Lett. 2007, 270, 284–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalazar, A.L.; Gliemmo, M.F.; Soria, M.; Campos, C.A. Modelling growth/no growth interface of Zygosaccharomyces bailii in simulated acid sauces as a function of natamycin, xanthan gum and sodium chloride concentrations. Food Res. Int. 2019, 116, 916–924. [Google Scholar] [CrossRef]
- Arroyo-López, F.N.; Bautista-Gallego, J.; Romero-Gil, V.; Rodríguez-Gómez, F.; Garrido-Fernández, A. Growth/no growth interfaces of table olive related yeasts for natamycin, citric acid and sodium chloride. Int. J. Food Microbiol. 2012, 155, 257–262. [Google Scholar] [CrossRef]
- Garnier, L.; Salas, M.L.; Pinon, N.; Wiernasz, N.; Pawtowski, A.; Coton, E.; Mounier, J.; Valence, F. Technical note: High-throughput method for antifungal activity screening in a cheese-mimicking model. J. Dairy Sci. 2018, 101, 4971–4976. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.; Chan, K.; Kim, J.H. Chemosensitization as a Means to Augment Commercial Antifungal Agents. Front. Microbiol. 2012, 3, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Chang, P.-K.; Chan, K.L.; Faria, N.C.G.; Mahoney, N.; Kim, Y.K.; Martins, M.D.L.; Campbell, B.C. Enhancement of Commercial Antifungal Agents by Kojic Acid. Int. J. Mol. Sci. 2012, 13, 13867–13880. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungi | Strain Characteristics | Source |
|---|---|---|
| A. flavus 3357 | Mycotoxigenic (aflatoxin (AF) producer), human pathogen (aspergillosis), reference strain for genome sequencing | NRRL 1 |
| A. flavus 4212 | Mycotoxigenic (AF producer), human pathogen (aspergillosis) | NRRL |
| A. parasiticus 2999 | Mycotoxigenic (AF producer) | NRRL |
| A. parasiticus 5862 | Mycotoxigenic (AF producer) | NRRL |
| A. fumigatus AF293 | Human pathogen (aspergillosis), reference clinical strain for genome sequencing | UT 2, [31] |
| A. fumigatus sakAΔ | Mitogen-activated protein kinase (MAPK) gene deletion mutant derived from AF293 | UT, [31] |
| A. fumigatus mpkCΔ | MAPK gene deletion mutant derived from AF293 | UT, [32] |
| P. expansum W1 | Mycotoxigenic (patulin producer; parental strain) | WSU 3, [33] |
| P. expansum FR2 | Fludioxonil (FLU)-resistant mutant derived from W1 | WSU, [33] |
| N. fischeri 96468 | Food spoilage fungus, heat resistant | ATCC 4 |
| NAT (μg mL−1) | Values | PDA (pH 3.5) | Juice 1 | Juice 2 | Juice 3 | Juice 4 | Juice 5 | Juice 6 | Juice 7 | Juice 8 | Juice 9 | Juice 10 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A. fumigatus | Average | 90.0 | 86.3 | 72.3 | 77.0 | 77.7 | 79.3 | 67.3 | 90.0 | 72.7 | 66.7 | 54.7 |
| 2 | SD | 2.6 | 5.8 | 2.3 | 7.0 | 4.0 | 4.0 | 3.1 | 0.0 | 2.5 | 6.7 | 10.8 |
| p-value | - | 0.37 | 0.00 * | 0.04 * | 0.01 * | 0.02 * | 0.00 * | 1.00 | 0.00 * | 0.01 * | 0.01 * | |
| 4 | Average | 49.7 | 62.3 | 41.7 | 45.3 | 39.7 | 52.0 | 24.3 | 81.7 | 19.3 | 19.0 | 33.0 |
| SD | 11.5 | 5.8 | 2.3 | 8.5 | 11.5 | 3.5 | 7.5 | 2.3 | 5.1 | 0.0 | 14.7 | |
| p-value | - | 0.16 | 0.31 | 0.63 | 0.35 | 0.75 | 0.03 * | 0.01 * | 0.01 * | 0.01 * | 0.20 | |
| 8 | Average | 8.3 | 33.3 | 19.0 | 24.3 | 33.0 | 16.7 | 15.7 | 49.3 | 10.3 | 19.0 | 33.0 |
| SD | 14.4 | 6.8 | 1.7 | 2.1 | 0.0 | 2.9 | 0.6 | 5.1 | 2.3 | 0.0 | 14.7 | |
| p-value | - | 0.05 | 0.27 | 0.13 | 0.04 * | 0.38 | 0.43 | 0.01 * | 0.82 | 0.27 | 0.11 | |
| 16 | Average | 0.0 | 18.0 | 15.7 | 17.7 | 29.0 | 13.3 | 11.7 | 0.0 | 9.0 | 19.0 | 33.0 |
| SD | 0.0 | 0.0 | 2.9 | 2.1 | 3.5 | 2.3 | 0.6 | 0.0 | 0.0 | 0.0 | 14.7 | |
| p-value | - | 0.00 * | 0.00 * | 0.00 * | 0.00 * | 0.00 * | 0.00 * | ND | 0.00 * | 0.00 * | 0.02 * | |
| Foodborne fungi | Average | 62.7 | 82.1 | 73.9 | 77.1 | 82.7 | 80.6 | 72.6 | 82.0 | 74.7 | 75.7 | 73.7 |
| 2 | SD | 43.2 | 19.4 | 19.0 | 13.4 | 22.4 | 13.0 | 17.0 | 14.0 | 22.8 | 18.1 | 8.4 |
| p-value | - | 0.30 | 0.54 | 0.42 | 0.30 | 0.32 | 0.58 | 0.28 | 0.53 | 0.48 | 0.52 | |
| 4 | Average | 52.7 | 73.6 | 56.4 | 56.4 | 77.6 | 57.4 | 52.4 | 59.0 | 60.7 | 65.6 | 55.4 |
| SD | 36.4 | 22.1 | 14.5 | 14.3 | 19.1 | 20.2 | 13.1 | 40.5 | 19.2 | 14.4 | 12.9 | |
| p-value | - | 0.22 | 0.81 | 0.81 | 0.14 | 0.77 | 0.99 | 0.77 | 0.62 | 0.40 | 0.86 | |
| 8 | Average | 35.7 | 66.4 | 42.3 | 40.6 | 67.3 | 42.7 | 36.7 | 50.4 | 46.9 | 52.6 | 41.4 |
| SD | 25.0 | 18.3 | 7.0 | 10.0 | 13.3 | 9.4 | 7.4 | 35.2 | 10.6 | 8.6 | 14.3 | |
| p-value | - | 0.02 * | 0.52 | 0.64 | 0.01 * | 0.50 | 0.92 | 0.38 | 0.30 | 0.12 | 0.61 | |
| 16 | Average | 5.3 | 55.7 | 30.3 | 30.1 | 54.1 | 26.6 | 22.1 | 40.6 | 33.4 | 39.3 | 35.1 |
| SD | 14.0 | 12.7 | 8.4 | 10.6 | 6.6 | 7.9 | 10.4 | 28.1 | 7.8 | 9.5 | 17.0 | |
| p-value | - | 0.00 * | 0.00 * | 0.00 * | 0.00 * | 0.00 * | 0.03 * | 0.01 * | 0.00 * | 0.00 * | 0.00 * |
| Juice No. NAT (μg mL−1) | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|---|---|---|---|---|---|---|---|---|---|
| Human pathogen (A. fumigatus) | ||||||||||
| 2 | 9 | 4 | 6 | 7 | 8 | 3 | 10 | 5 | 2 | 1 |
| 4 | 9 | 6 | 7 | 5 | 8 | 3 | 10 | 2 | 1 | 4 |
| 8 | 9 | 4 | 6 | 7 | 3 | 2 | 10 | 1 | 4 | 7 |
| 16 | 7 | 5 | 6 | 9 | 4 | 3 | 1 | 2 | 8 | 10 |
| Average 2 | 8.50 | 4.75 | 6.25 | 7.00 | 5.75 | 2.75 | 7.75 | 2.50 | 3.75 | 5.50 |
| Final 3 | 10 | 4 | 7 | 8 | 6 | 2 | 9 | 1 | 3 | 5 |
| Foodborne contaminants | ||||||||||
| 2 | 9 | 3 | 6 | 10 | 7 | 1 | 8 | 4 | 5 | 2 |
| 4 | 9 | 3 | 3 | 10 | 5 | 1 | 6 | 7 | 8 | 2 |
| 8 | 9 | 4 | 2 | 10 | 5 | 1 | 7 | 6 | 8 | 3 |
| 16 | 10 | 4 | 3 | 9 | 2 | 1 | 8 | 5 | 7 | 6 |
| Average 2 | 9.25 | 3.50 | 3.50 | 9.75 | 4.75 | 1.00 | 7.25 | 5.50 | 7.00 | 3.25 |
| Final 3 | 9 | 3 | 3 | 10 | 5 | 1 | 8 | 6 | 7 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.H.; Tam, C.C.; Chan, K.L.; Cheng, L.W.; Land, K.M.; Friedman, M.; Chang, P.-K. Antifungal Efficacy of Redox-Active Natamycin against Some Foodborne Fungi—Comparison with Aspergillus fumigatus. Foods 2021, 10, 2073. https://doi.org/10.3390/foods10092073
Kim JH, Tam CC, Chan KL, Cheng LW, Land KM, Friedman M, Chang P-K. Antifungal Efficacy of Redox-Active Natamycin against Some Foodborne Fungi—Comparison with Aspergillus fumigatus. Foods. 2021; 10(9):2073. https://doi.org/10.3390/foods10092073
Chicago/Turabian StyleKim, Jong H., Christina C. Tam, Kathleen L. Chan, Luisa W. Cheng, Kirkwood M. Land, Mendel Friedman, and Perng-Kuang Chang. 2021. "Antifungal Efficacy of Redox-Active Natamycin against Some Foodborne Fungi—Comparison with Aspergillus fumigatus" Foods 10, no. 9: 2073. https://doi.org/10.3390/foods10092073
APA StyleKim, J. H., Tam, C. C., Chan, K. L., Cheng, L. W., Land, K. M., Friedman, M., & Chang, P. -K. (2021). Antifungal Efficacy of Redox-Active Natamycin against Some Foodborne Fungi—Comparison with Aspergillus fumigatus. Foods, 10(9), 2073. https://doi.org/10.3390/foods10092073