
Muscle and Serum Metabolomics for Different Chicken Breeds under Commercial Conditions by GC–MS

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Sample Collection and Ethical Approval
2.3. Preparation of Muscle Samples for GC–MS Analysis
2.4. Preparation of Serum Samples for GC–MS Analysis
2.5. Preparation of Quality Control (QC) Samples
2.6. Preparation of Standard Solution
2.7. GC–MS Analysis
2.8. GC–MS Data Processing
2.9. Statistical Analysis
2.10. Method Validation of Extraction Protocols
2.11. Quantification of Metabolites in Pectoralis Major and Serum
2.12. Calculation of the Recovery of the Internal Standards
3. Results
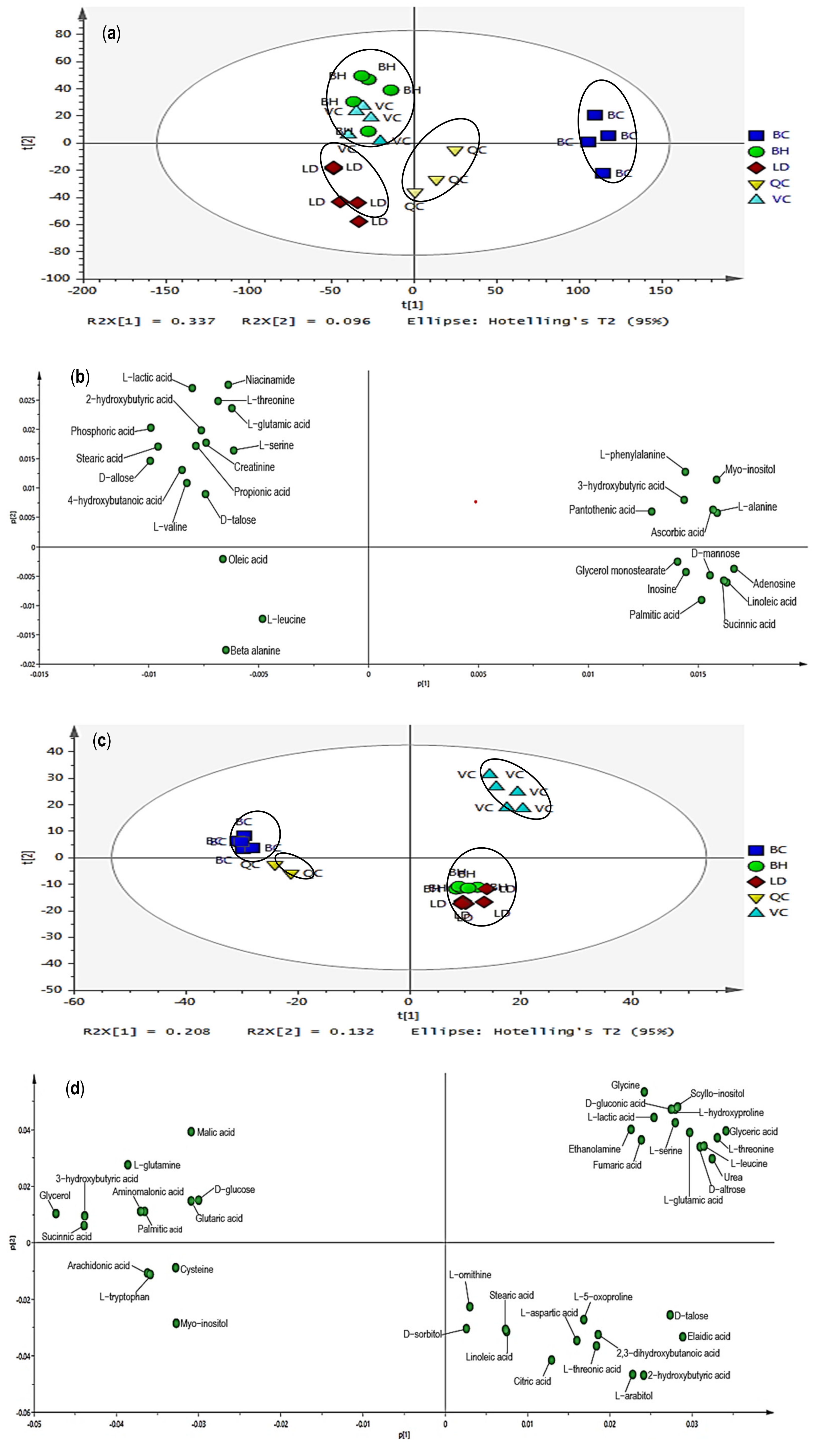
3.1. GC–MS Chromatogram of Pectoralis Major and Serum
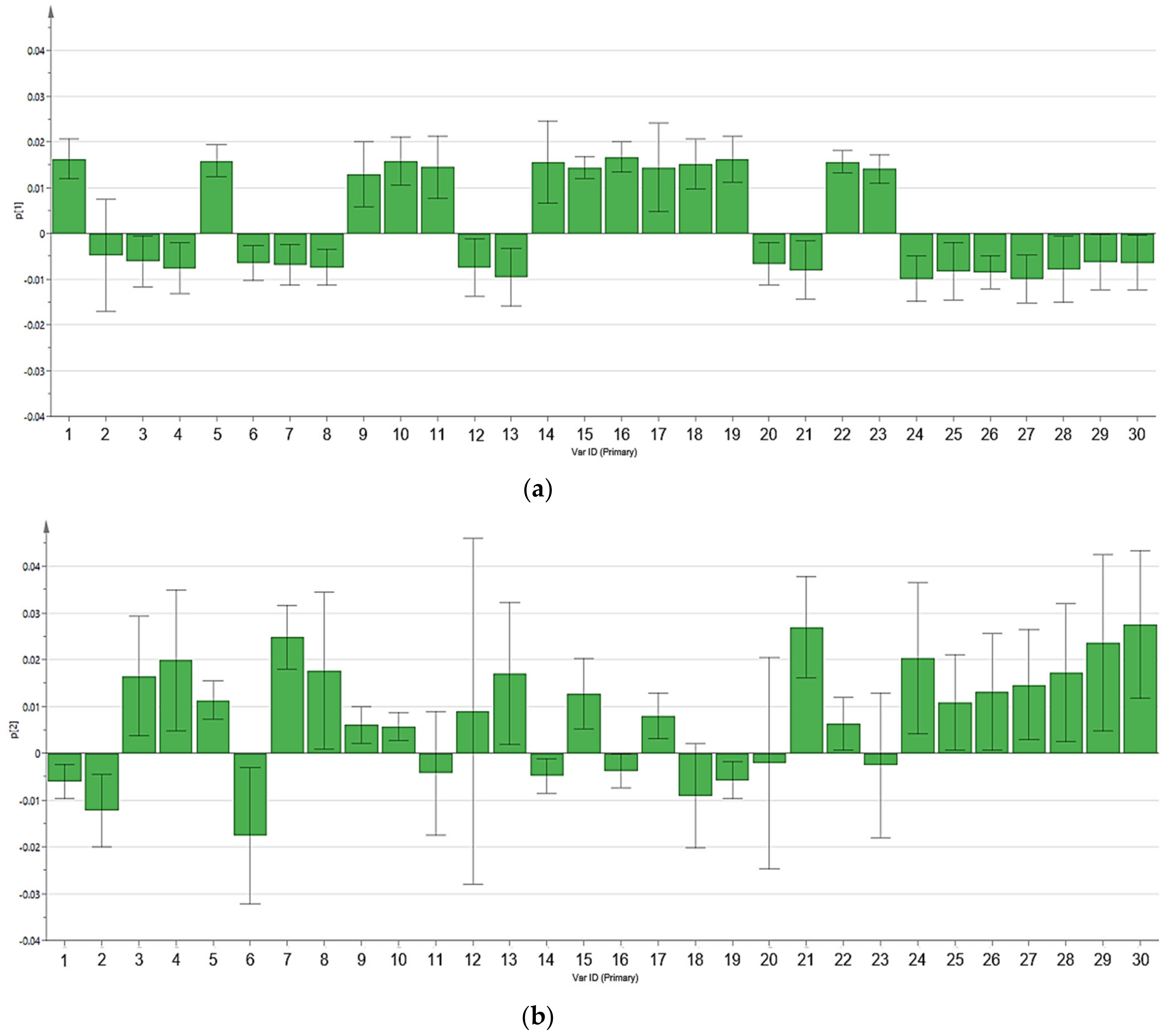
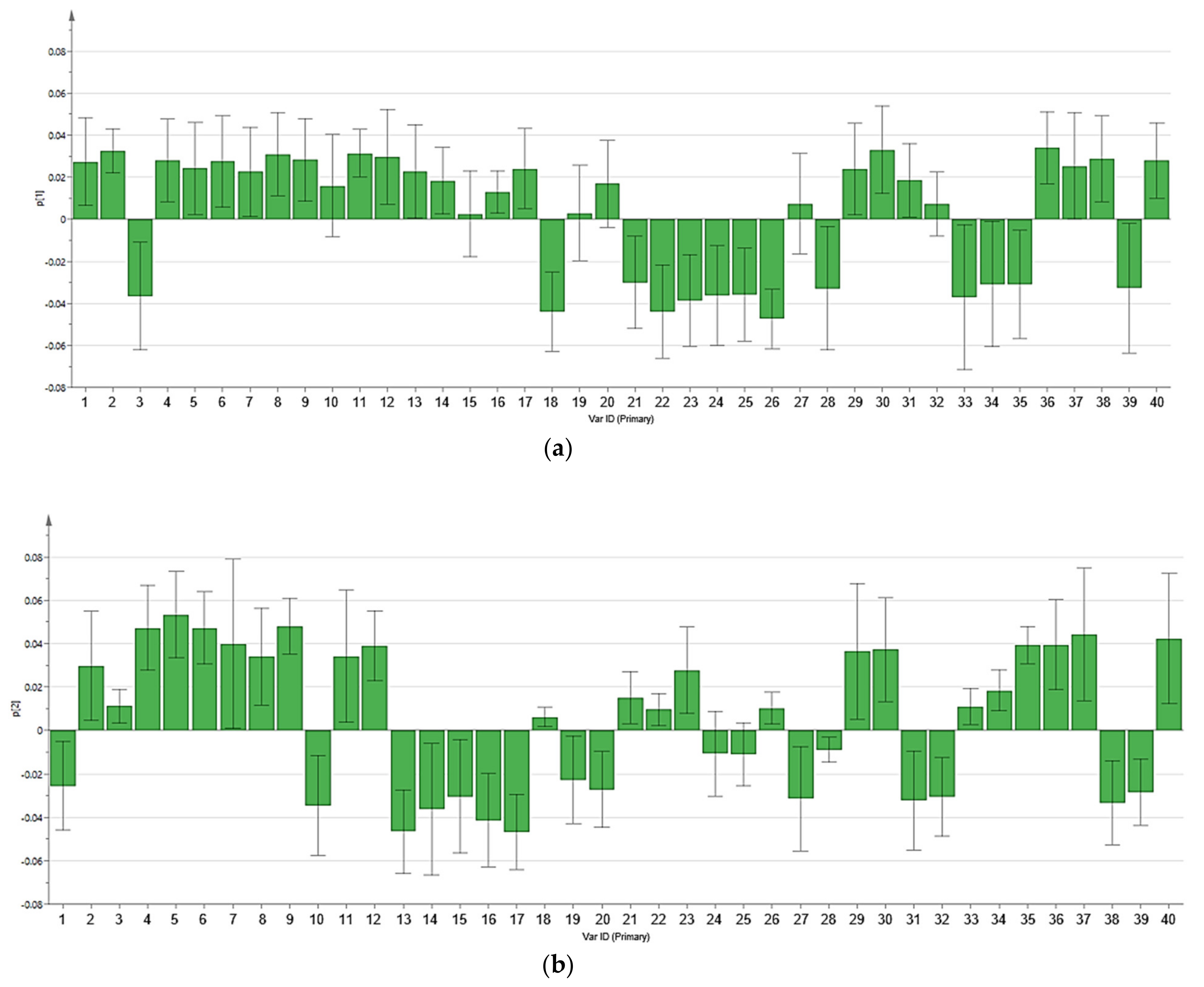
3.2. Multivariate Data Analysis
3.3. Reliability of the Extraction Protocols
3.4. Quantification and Semi-Quantification of Samples
3.5. Proximate Analysis of Commercial Feed Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Semwogerere, F.; Neethling, J.; Muchenje, V.; Hoffman, L.C. Meat Quality, Fatty Acid Profile, and Sensory Attributes of Spent Laying Hens Fed Expeller Press Canola Meal or a Conventional Diet. Poult. Sci. 2019, 98, 3557–3570. [Google Scholar] [CrossRef] [PubMed]
- Petracci, M.; Bianchi, M.; Mudalal, S.; Cavani, C. Functional Ingredients for Poultry Meat Products. Trends Food Sci. Technol. 2013, 33, 27–39. [Google Scholar] [CrossRef]
- Feng, Y.; Cai, Y.; Fu, X.; Zheng, L.; Xiao, Z.; Zhao, M. Comparison of Aroma-Active Compounds in Broiler Broth and Native Chicken Broth by Aroma Extract Dilution Analysis (AEDA), Odor Activity Value (OAV) and Omission Experiment. Food Chem. 2018, 265, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Mohd Shahmi Hakimi, M.; Alias, Z.; Wan Khadijah, W.E.; Abdullah, R.B.; Rahman, M.M. Effect of Diet Containing Palm Kernel Cake and Coconut Meal Cake on Growth Performance and Carcass Quality of Free-Range Chicken in Malaysia (Kesan Daripada Diet Yang Mengandungi Isirung Kernal Sawit Dan Isirung Kelapa Pada. Sains Malays. 2019, 48, 991–998. [Google Scholar] [CrossRef]
- Azahan, E.A.E. The Red and Black-Red Native Chickens of Malaysia. Malays. Agric. Res. Dev. Inst. Res. J. 1994, 22, 73–78. [Google Scholar]
- Aini, I. Indigenous Chicken Production in South-East Asia. Worlds Poult. Sci. J. 1990, 46, 51–57. [Google Scholar] [CrossRef]
- The Star. Available online: https://www.thestar.com.my/news/nation/2019/07/15/beware-of-coloured-chicken/ (accessed on 10 May 2021).
- Malay Mail. Available online: https://www.malaymail.com/news/malaysia/2019/05/27/buyers-beware-ahead-of-raya-breeders-warn-of-fake-kampung-chicken-in-market/1756703 (accessed on 10 May 2021).
- Fontanesi, L. Meat Authenticity and Tracebility. In Lawrie’s Meat Science; Woodhead Publishing: Cambridge, UK, 2017; pp. 585–633. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics—The Link between Genotypes and Phenotypes. Funct. Genom. 2002, 48, 155–171. [Google Scholar] [CrossRef]
- Cevallos-Cevallos, J.M.; Reyes-De-Corcuera, J.I.; Etxeberria, E.; Danyluk, M.D.; Rodrick, G.E. Metabolomic Analysis in Food Science: A Review. Trends Food Sci. Technol. 2009, 20, 557–566. [Google Scholar] [CrossRef]
- Caboni, P.; Maxia, D.; Scano, P.; Addis, M.; Dedola, A.; Pes, M.; Murgia, A.; Casula, M.; Profumo, A.; Pirisi, A. A Gas Chromatography-Mass Spectrometry Untargeted Metabolomics Approach to Discriminate Fiore Sardo Cheese Produced from Raw or Thermized Ovine Milk. J. Dairy Sci. 2019, 102, 5005–5018. [Google Scholar] [CrossRef]
- Man, K.Y.; Chan, C.O.; Tang, H.H.; Dong, N.-P.; Capozzi, F.; Wong, K.H.; Kwok, K.W.H.; Chan, H.M.; Mok, D.K.W. Mass Spectrometry-Based Untargeted Metabolomics Approach for Differentiation of Beef of Different Geographic Origins. Food Chem. 2021, 338, 127847. [Google Scholar] [CrossRef] [PubMed]
- Ueda, S.; Iwamoto, E.; Kato, Y.; Shinohara, M.; Shirai, Y.; Yamanoue, M. Comparative Metabolomics of Japanese Black Cattle Beef and Other Meats Using Gas Chromatography–Mass Spectrometry. Biosci. Biotechnol. Biochem. 2018, 83, 137–147. [Google Scholar] [CrossRef]
- Surowiec, I.; Fraser, P.D.; Patel, R.; Halket, J.; Bramley, P.M. Metabolomic Approach for the Detection of Mechanically Recovered Meat in Food Products. Food Chem. 2011, 125, 1468–1475. [Google Scholar] [CrossRef]
- Mabuchi, R.; Adachi, M.; Kikutani, H.; Tanimoto, S. Discriminant Analysis of Muscle Tissue Type in Yellowtail Seriola Quinqueradiata Muscle Based on Metabolic Component Profiles. Food Sci. Technol. Res. 2018, 24, 883–891. [Google Scholar] [CrossRef]
- Rochfort, S.J.; Ezernieks, V.; Maher, A.D.; Ingram, B.A.; Olsen, L. Mussel Metabolomics—Species Discrimination and Provenance Determination. Food Res. Int. 2013, 54, 1302–1312. [Google Scholar] [CrossRef]
- Beauclercq, S.; Nadal-Desbarats, L.; Hennequet-Antier, C.; Collin, A.; Tesseraud, S.; Bourin, M.; Le Bihan-Duval, E.; Berri, C. Serum and Muscle Metabolomics for the Prediction of Ultimate PH, a Key Factor for Chicken-Meat Quality. J. Proteome Res. 2016, 15, 1168–1178. [Google Scholar] [CrossRef]
- Mabuchi, R.; Ishimaru, A.; Tanaka, M.; Kawaguchi, O.; Tanimoto, S. Metabolic Profiling of Fish Meat by Gc-Ms Analysis, and Correlations with Taste Attributes Obtained Using an Electronic Tongue. Metabolites 2019, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Trimigno, A.; Münger, L.; Picone, G.; Freiburghaus, C.; Pimentel, G.; Vionnet, N.; Pralong, F.; Capozzi, F.; Badertscher, R.; Vergères, G. GC-MS Based Metabolomics and NMR Spectroscopy Investigation of Food Intake Biomarkers for Milk and Cheese in Serum of Healthy Humans. Metabolites 2018, 8, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mykhailenko, O.; Ivanauskas, L.; Bezruk, I.; Lesyk, R.; Georgiyants, V. Comparative Investigation of Amino Acids Content in the Dry Extracts of Juno Bucharica, Gladiolus Hybrid Zefir, Iris Hungarica, Iris Variegata and Crocus Sativus Raw Materials of Ukrainian Flora. Sci. Pharm. 2020, 88, 8. [Google Scholar] [CrossRef] [Green Version]
- Grabež, V.; Bjelanović, M.; Rohloff, J.; Martinović, A.; Berg, P.; Tomović, V.; Rogić, B.; Egelandsdal, B. The Relationship between Volatile Compounds, Metabolites and Sensory Attributes: A Case Study Using Lamb and Sheep Meat. Small Rumin. Res. 2019, 181, 12–20. [Google Scholar] [CrossRef]
- Shi, Y.; Li, X.; Huang, A. A Metabolomics-Based Approach Investigates Volatile Flavor Formation and Characteristic Compounds of the Dahe Black Pig Dry-Cured Ham. Meat Sci. 2019, 158, 107904. [Google Scholar] [CrossRef]
- Murugesu, S.; Ibrahim, Z.; Ahmed, Q.U.; Yusoff, N.I.N.; Uzir, B.F.; Perumal, V.; Abas, F.; Saari, K.; El-Seedi, H.; Khatib, A. Characterization of α-Glucosidase Inhibitors from Clinacanthus Nutans Lindau Leaves by Gas Chromatography-Mass Spectrometry-Based Metabolomics and Molecular Docking Simulation. Molecules 2018, 23, 2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, S.; Yi, L.; Yun, Y.; Zhang, X.; Liang, Y. A Combination of GC-MS and Chemometrics Reveals Metabolic Differences between Serum and Plasma. Anal. Methods 2015, 7, 1751–1757. [Google Scholar] [CrossRef]
- Yamada, T.; Kamiya, M.; Higuchi, M. Gas Chromatography–Mass Spectrometry-based Metabolomic Analysis of Wagyu and Holstein Beef. Metabolites 2020, 10, 95. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, L.; Byrne, T.; Johansson, E.; Trygg, J.; Vikstrom, C. Multi-and Megavariate Data Analysis Basic Principles and Applications, 2nd ed.; Umetrics Academy: Umeå, Sweden, 2006; pp. 33–53. [Google Scholar]
- Guntarti, A.; Ningrum, K.P.; Gandjar, I.G.; Salamah, N. Authentication of Sprague Dawley Rats (Rattus Norvegicus) Fat with GC-MS (Gas Chromatography-Mass Spectrometry) Combined with Chemometrics. Int. J. Appl. Pharm. 2021, 13, 134–139. [Google Scholar] [CrossRef]
- Ivanne, C.; Roy, L.; John, L.; Roberto, M.; La, M.; John, M.; Sandrine, W.; Claus, P. NMR-Based Metabolic Characterization of Chicken Tissues and Biofluids: A Model for Avian Research. Metabolomics 2016, 12, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Mueller, S.; Taddei, L.; Albiker, D.; Kreuzer, M.; Siegrist, M.; Messikommer, R.E.; Gangnat, I.D.M. Growth, Carcass, and Meat Quality of 2 Dual-Purpose Chickens and a Layer Hybrid Grown for 67 or 84 D Compared with Slow-Growing Broilers. J. Appl. Poult. Res. 2020, 29, 185–196. [Google Scholar] [CrossRef]
- Xiao, Z.; Ge, C.; Zhou, G.; Zhang, W.; Liao, G. 1H NMR-Based Metabolic Characterization of Chinese Wuding Chicken Meat. Food Chem. 2019, 274, 574–582. [Google Scholar] [CrossRef]
- Panel on Additives and Products or Substances used in Animal Feed EFSA. Scientific Opinion on the Safety and Efficacy of AviMatrix® (Benzoic Acid, Calcium Formate and Fumaric Acid) for Chickens for Fattening, Chickens Reared for Laying, Minor Avian Species for Fattening and Minor Avian Species Reared to Point of Lay. EFSA J. 2015, 13, 3794. [Google Scholar] [CrossRef]
- Peng, M.L.; Han, J.; Li, L.L.; Ma, H.T. Metabolomics Reveals the Mechanism of (-)-Hydroxycitric Acid Promotion of Protein Synthesis and Inhibition of Fatty Acid Synthesis in Broiler Chickens. Animal 2018, 12, 774–783. [Google Scholar] [CrossRef]
- Donsbough, A.L.; Powell, S.; Waguespack, A.; Bidner, T.D.; Southern, L.L. Uric Acid, Urea, and Ammonia Concentrations in Serum and Uric Acid Concentration in Excreta as Indicators of Amino Acid Utilization in Diets for Broilers. Poult. Sci. 2010, 89, 287–294. [Google Scholar] [CrossRef]
- Kong, Y.; Yang, X.; Ding, Q.; Zhang, Y.Y.; Sun, B.G.; Chen, H.T.; Sun, Y. Comparison of Non-Volatile Umami Components in Chicken Soup and Chicken Enzymatic Hydrolysate. Food Res. Int. 2017, 102, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Zotte, A.D.; Gleeson, E.; Franco, D.; Cullere, M.; Lorenzo, M. Proximate Composition, Amino Acid Profile, and Oxidative Stability of Slow-Growing Indigenous Chickens Compared with Commercial Broiler Chickens. Foods 2020, 9, 546. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Burghardt, R.C.; Johnson, G.A.; Kim, S.W.; Knabe, D.A.; Li, P.; Li, X.; McKnight, J.R.; Satterfield, M.C.; et al. Proline and Hydroxyproline Metabolism: Implications for Animal and Human Nutrition. Amino Acids 2011, 40, 1053–1063. [Google Scholar] [CrossRef] [Green Version]
- Lengkidworraphiphat, P.; Wongpoomchai, R.; Bunmee, T.; Chariyakornkul, A.; Chaiwang, N.; Jaturasitha, S. Taste-Active and Nutritional Components of Thai Native Chicken Meat: A Perspective of Consumer Satisfaction. Food Sci. Anim. Resour. 2021, 41, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Okarini, I.A.; Purnomo, H.; Aulanni’am; Radiati, L.E. Proximate, Total Phenolic, Antioxidant Activity and Amino Acids Profile of Bali Indigenous Chicken, Spent Laying Hen and Broiler Breast Fillet. Int. J. Poult. Sci. 2013, 12, 415–420. [Google Scholar] [CrossRef] [Green Version]
- Kidd, M.T.; Poernama, F.; Wibowo, T.; Maynard, C.W.; Liu, S.Y. Dietary Branched-Chain Amino Acid Assessment in Broilers from 22 to 35 Days of Age. J. Anim. Sci. Biotechnol. 2021, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Maynard, C.W.; Liu, S.Y.; Lee, J.T.; Caldas, J.V.; Diehl, J.J.E.; Rochell, S.J.; Kidd, M.T. Evaluation of Branched-Chain Amino Acids in Male Cobb MV × 500 Broiler Chickens by Using Box-Behnken Response Surface Design. Anim. Feed Sci. Technol. 2021, 271, 114710. [Google Scholar] [CrossRef]
- Clements, J.R.S.; Darnell, B. Myo-Inositol Content of Common Foods: Development of a High-Myo-Inositol Diet. Am. J. Clin. Nutr. 1980, 33, 1954–1967. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-uarquin, F.; Rodehutscord, M.; Huber, K. Myo-Inositol: Its Metabolism and Potential Implications for Poultry Nutrition—A Review. Poult. Sci. 2020, 99, 893–905. [Google Scholar] [CrossRef] [PubMed]
- Cowieson, A.J.; Zhai, H.X. Research Note: The Effect of Sequential Displacement of Dietary Dextrose with Myo-Inositol on Broiler Chicken Growth Performance, Bone Characteristics, Ileal Nutrient Digestibility, and Total Tract Nutrient Retention. Poult. Sci. 2021, 100, 993–997. [Google Scholar] [CrossRef]
- Qaid, M.M.; Al-garadi, M.A. Protein and Amino Acid Metabolism in Poultry during and after Heat Stress: A Review. Animals 2021, 11, 1167. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Classification | Authentic Village Chicken | Broiler (Cobb) | Colored Broiler (Hubbard) | Spent Laying Hen (Dekalb) |
|---|---|---|---|---|
| Amino acid | 9 (25.7) | 10 (23.8) | 7 (23.3) | 10 (27.8) |
| Fatty acid | 7 (20.0) | 9 (21.4) | 6 (20.0) | 9 (25.0) |
| Organic acid | 6 (17.1) | 5 (11.9) | 4 (13.3) | 5 (13.9) |
| Sugar | 3 (8.6) | 4 (9.5) | 4 (13.3) | 4 (11.1) |
| Sugar alcohol | 2 (5.7) | 2 (4.8) | 1 (3.3) | 1 (2.8) |
| Vitamin | 1 (2.9) | 3 (7.1) | 2 (6.7) | 1 (2.8) |
| Other | 7 (20.0) | 9 (21.4) | 6 (20.0) | 6 (16.7) |
| Total | 35 (100) | 42 (100) | 30 (100) | 36 (100) |
| Classification | Authentic Village Chicken | Broiler (Cobb) | Colored Broiler (Hubbard) | Spent Laying Hen (Dekalb) |
|---|---|---|---|---|
| Amino acid | 14 (28.6) | 19 (36.5) | 14 (34.1) | 12 (27.9) |
| Fatty acid | 5 (10.2) | 5 (9.6) | 7 (17.1) | 6 (14.0) |
| Organic acid | 10 (20.4) | 10 (19.2) | 6 (14.6) | 9 (20.9) |
| Sugar | 5 (10.2) | 5 (9.6) | 4 (9.8) | 5 (11.6) |
| Sugar alcohol | 6 (12.2) | 6 (11.5) | 4 (9.8) | 6 (14.0) |
| Other | 9 (18.4) | 7 (13.5) | 6 (14.6) | 5 (11.6) |
| Total | 49 (100) | 52 (100) | 41 (100) | 43 (100) |
| No. | RT (min) | Tentative Metabolites | Molecular Weight (g/mol) | Derivatized Molecular Weight (g/mol) | Probability (%) | Molecular Formula | Derivatized Molecular Formula |
|---|---|---|---|---|---|---|---|
| 1 | 7.005 | 2−hydroxybutyric acid | 104.10 | 248.13 | 92.87 | C4H8O3 | C10H24O3Si2 |
| 2 | 7.892 | 3−hydroxybutyric acid | 104.10 | 248.13 | 93.39 | C4H8O3 | C10H24O3Si2 |
| 3 | 10.201 | 4−hydroxybutanoic acid | 104.10 | 248.13 | 95.20 | C4H8O3 | C10H24O3Si2 |
| 4 | 55.193 | Adenosine | 267.24 | 555.26 | 92.54 | C10H13N5O4 | C22H45N5O4Si4 |
| 5 | 36.315 | Ascorbic acid | 176.12 | 464.19 | 95.73 | C6H8O6 | C18H40O6Si4 |
| 6 | 8.743 | Beta-alanine | 89.09 | 233.13 | 92.87 | C3H7NO2 | C9H23NO2Si2 |
| 7 | 22.055 | Creatinine | 113.12 | 329.66 | 92.16 | C4H7N3O | C13H31N3OSi3 |
| 8 | 35.148 | D−talose | 180.16 | 569.29 | 91.78 | C6H12O6 | C22H55NO6Si5 |
| 9 | 35.616 | D−allose | 180.16 | 569.29 | 91.64 | C6H12O6 | C22H55NO6Si5 |
| 10 | 34.772 | D−mannose | 180.16 | 540.26 | 94.32 | C6H12O6 | C21H52O6Si5 |
| 11 | 58.382 | Glycerol monostearate | 358.56 | 502.39 | 95.93 | C21H42O4 | C27H58O4Si2 |
| 12 | 53.819 | Inosine | 268.23 | 556.24 | 93.76 | C10H12N4O5 | C22H44N4O5Si4 |
| 13 | 5.62 | L−lactic acid | 90.08 | 234.11 | 95.64 | C3H6O3 | C9H22O3Si2 |
| 14 | 6.361 | L−alanine | 89.09 | 233.13 | 90.94 | C3H7NO2 | C9H23NO2Si2 |
| 15 | 24.789 | L−glutamic acid | 147.13 | 363.17 | 91.10 | C5H9NO4 | C14H33NO4Si3 |
| 16 | 7.643 | L−leucine | 131.17 | 203.13 | 94.27 | C6H13NO2 | C9H21NO2Si |
| 17 | 24.576 | L−phenylalanine | 165.19 | 309.16 | 91.11 | C9H11NO2 | C15H27NO2Si2 |
| 18 | 10.942 | L−serine | 105.09 | 249.12 | 94.74 | C3H7NO3 | C9H23NO3Si2 |
| 19 | 16.046 | L−threonine | 119.12 | 335.18 | 91.59 | C4H9NO3 | C13H33NO3Si3 |
| 20 | 9.63 | L−valine | 117.15 | 261.16 | 92.04 | C5H11NO2 | C11H27NO2Si2 |
| 21 | 43.301 | Linoleic acid | 280.45 | 352.28 | 98.84 | C18H32O2 | C21H40O2Si |
| 22 | 40.847 | Myo-inositol | 180.16 | 612.30 | 91.98 | C6H12O6 | C24H60O6Si6 |
| 23 | 18.81 | Niacinamide | 122.12 | 194.09 | 91.08 | C6H6N2O | C9H14N2OSi |
| 24 | 43.472 | Oleic acid | 282.46 | 354.30 | 91.88 | C18H34O2 | C21H42O2Si |
| 25 | 38.502 | Palmitic acid | 256.42 | 328.28 | 93.17 | C16H32O2 | C19H40O2Si |
| 26 | 37.390 | Pantothenic acid | 219.23 | 435.23 | 90.40 | C9H17NO5 | C18H41NO5Si3 |
| 27 | 30.373 | Phosphoric acid | 98.00 | 460.17 | 96.31 | H3PO4 | C15H41O6PSi4 |
| 28 | 31.873 | Propionic acid | 74.08 | 474.15 | 92.78 | C3H6O2 | C15H39O7PSi4 |
| 29 | 44.286 | Stearic acid | 284.48 | 356.31 | 94.06 | C18H36O2 | C21H44O2Si |
| 30 | 12.972 | Succinic acid | 118.09 | 262.11 | 98.30 | C4H6O4 | C10H22O4Si2 |
| No. | RT (min) | Tentative Metabolites | Molecular Weight (g/mol) | Derivatized Molecular Weight (g/mol) | Probability (%) | Molecular Formula | Derivatized Molecular Formula |
|---|---|---|---|---|---|---|---|
| 1 | 14.758 | 2,3-dihydroxybutanoic acid | 120.10 | 336.16 | 91.07 | C4H8O4 | C13H32O4Si3 |
| 2 | 6.932 | 2-hydroxybutyric acid | 104.10 | 248.13 | 94.29 | C4H8O3 | C10H24O3Si2 |
| 3 | 7.856 | 3-hydroxybutyric acid | 104.10 | 248.13 | 96.23 | C4H8O3 | C10H24O3Si2 |
| 4 | 19.059 | Aminomalonic acid | 119.08 | 335.14 | 93.30 | C3H5NO4 | C12H29NO4Si3 |
| 5 | 47.559 | Arachidonic acid | 304.47 | 376.28 | 94.80 | C20H32O2 | C23H40O2Si |
| 6 | 32.129 | Citric acid | 192.12 | 480.19 | 91.79 | C6H8O7 | C18H40O7Si4 |
| 7 | 22.201 | Cysteine | 121.16 | 337.14 | 96.55 | C3H7NO2S | C12H31NO2SSi3 |
| 8 | 37.986 | D-altrose | 180.16 | 540.26 | 95.21 | C6H12O6 | C21H52O6Si5 |
| 9 | 37.117 | D-gluconic acid | 196.16 | 628.30 | 95.51 | C6H12O7 | C24H60O7Si6 |
| 10 | 35.36 | D-glucose | 180.16 | 569.29 | 94.39 | C6H12O6 | C22H55NO6Si5 |
| 11 | 36.102 | D-sorbitol | 182.17 | 614.32 | 92.80 | C6H14O6 | C24H62O6Si6 |
| 12 | 34.808 | D-talose | 180.16 | 540.26 | 95.81 | C6H12O6 | C21H52O6Si5 |
| 13 | 43.442 | Elaidic acid | 282.46 | 354.30 | 95.21 | C18H34O2 | C21H42O2Si |
| 14 | 11.246 | Ethanolamine | 61.08 | 277.17 | 93.99 | C2H7NO | C11H31NOSi3 |
| 15 | 14.114 | Fumaric acid | 116.07 | 260.09 | 93.08 | C4H4O4 | C10H20O4Si2 |
| 16 | 23.13 | Glutaric acid | 132.11 | 319.13 | 90.47 | C5H8O4 | C12H25NO5Si2 |
| 17 | 13.846 | Glyceric acid | 106.08 | 322.15 | 91.52 | C3H6O4 | C12H30O4Si3 |
| 18 | 11.781 | Glycerol | 92.09 | 308.17 | 91.88 | C3H8O3 | C12H32O3Si3 |
| 19 | 6.683 | Glycine | 75.07 | 219.11 | 92.33 | C2H5NO2 | C8H21NO2Si2 |
| 20 | 28.555 | L-arabitol | 152.15 | 512.27 | 91.68 | C5H12O5 | C20H52O5Si5 |
| 21 | 21.07 | L-aspartic acid | 133.10 | 349.16 | 92.53 | C4H7NO4 | C13H31NO4Si3 |
| 22 | 24.728 | L-glutamic acid | 147.13 | 363.17 | 92.22 | C5H9NO4 | C14H33NO4Si3 |
| 23 | 30.172 | L-glutamine | 146.19 | 362.19 | 91.72 | C5H10N2O3 | C14H34N2O3Si3 |
| 24 | 21.198 | L-hydroxyproline | 131.13 | 347.18 | 90.40 | C5H9NO3 | C14H33NO3Si3 |
| 25 | 5.389 | L-lactic acid | 90.08 | 234.11 | 96.28 | C3H6O3 | C9H22O3Si2 |
| 26 | 7.576 | L-leucine | 131.17 | 203.13 | 94.00 | C6H13NO2 | C9H21NO2Si |
| 27 | 31.691 | L-ornithine | 132.16 | 420.25 | 95.20 | C5H12N2O2 | C17H44N2O2Si4 |
| 28 | 10.887 | L-serine | 105.09 | 249.12 | 93.85 | C3H7NO3 | C9H23NO3Si2 |
| 29 | 23.045 | L-threonic acid | 136.10 | 424.20 | 93.96 | C4H8O5 | C16H40O5Si4 |
| 30 | 15.997 | L-threonine | 119.12 | 335.18 | 93.52 | C4H9NO3 | C13H33NO3Si3 |
| 31 | 43.819 | L-tryptophan | 204.22 | 420.77 | 90.83 | C11H12N2O2 | C20H36N2O2Si3 |
| 32 | 43.259 | Linoleic acid | 280.45 | 352.28 | 96.06 | C18H32O2 | C21H40O2Si |
| 33 | 20.675 | L-5-oxoproline | 129.11 | 273.12 | 95.39 | C5H7NO3 | C11H23NO3Si2 |
| 34 | 19.916 | Malic acid | 134.09 | 350.14 | 92.41 | C4H6O5 | C13H30O5Si3 |
| 35 | 40.836 | Myo-inositol | 180.16 | 612.30 | 92.69 | C6H12O6 | C24H60O6Si6 |
| 36 | 38.496 | Palmitic acid | 256.42 | 328.28 | 93.58 | C16H32O2 | C19H40O2Si |
| 37 | 39.104 | Scyllo-inositol | 180.16 | 612.30 | 93.49 | C6H12O6 | C24H60O6Si6 |
| 38 | 12.886 | Succinic acid | 118.09 | 262.11 | 91.01 | C4H6O4 | C10H22O4Si2 |
| 39 | 44.24 | Stearic acid | 284.48 | 356.31 | 93.51 | C18H36O2 | C21H44O2Si |
| 40 | 10.328 | Urea | 60.06 | 204.11 | 92.73 | CH4N2O | C7H20N2OSi2 |
| Metabolites | Authentic Village Chicken | Broiler (Cobb) | Colored Broiler (Hubbard) | Spent Laying Hen (Dekalb) | ||||
|---|---|---|---|---|---|---|---|---|
| µg/mg | RSD (%) | µg/mg | RSD (%) | µg/mg | RSD (%) | µg/mg | RSD (%) | |
| D-allose 2 | 57.9 ± 3.1 b | 5.3 | n/d | - | 94.0 ± 8.8 a | 9.4 | n/d | - |
| L-phenylalanine 1 | 18.2 ± 2.4 b | 13.1 | 36.8 ± 7.4 a | 20.1 | 16.8 ± 1.8 b | 10.9 | 19.0 ± 2.5 b | 13.0 |
| Linoleic acid 3 | 60.1 ± 7.6 b | 12.7 | 108.2 ± 5.1 a | 4.7 | 54.7 ± 7.9 b | 14.4 | 50.6 ± 6.2 b | 12.2 |
| L-leucine 1 | 15.6 ± 1.3 b | 8.1 | n/d | - | n/d | - | 37.8 ± 3.2 a | 8.6 |
| L-valine 1 | 41.6 ± 5.2 a | 12.5 | 17.5 ± 3.6 b | 20.4 | 39.3 ± 4.7 a | 12.0 | 11.1 ± 1.4 b | 12.9 |
| Stearic acid 3 | 33.8 ± 7.6 c | 14.1 | 57.9 ± 6.3 b | 10.9 | 101.1 ± 5.5 a | 5.4 | 20.9 ± 1.5 d | 7.3 |
| Metabolites | Authentic Village Chicken | Broiler (Cobb) | Colored Broiler (Hubbard) | Spent Laying Hen (Dekalb) | ||||
|---|---|---|---|---|---|---|---|---|
| µg/mL | RSD (%) | µg/mL | RSD (%) | µg/mL | RSD (%) | µg/mL | RSD (%) | |
| Arachidonic acid 3 | 38.3 ± 1.5 b | 3.9 | 69.2 ± 5.4 a | 7.9 | 41.5 ± 3.7 b | 9.0 | 21.4 ± 1.6 c | 3.9 |
| Cysteine 1 | n/d | - | 32.6 ± 2.1 | 6.5 | n/d | - | n/d | - |
| D-glucose 2 | 2314.6 ± 99.3 b | 4.3 | 3398.5 ± 44.7 a | 1.3 | 1252.2 ± 96.5 d | 7.7 | 1584.8 ± 79.2 c | 8.7 |
| Fumaric acid 2 | 12.0 ± 1.0 | 8.7 | n/d | - | n/d | - | n/d | - |
| Linoleic acid 3 | 26.1 ± 1.9 c | 7.2 | 31.9 ± 1.8 bc | 5.7 | 66.9 ± 6.5 a | 9.7 | 33.2 ± 3.1 b | 9.4 |
| L-aspartic acid 3 | 30.4 ± 3.0 b | 9.8 | 24.9 ± 2.0 c | 8.0 | 45.1 ± 3.0 a | 6.7 | n/d | - |
| L-leucine 1 | 83.6 ± 6.7 a | 8.0 | 36.9 ± 3.4 c | 9.2 | 53.2 ± 4.1 b | 7.6 | 36.4 ± 2.5 c | 6.8 |
| Stearic acid 3 | 37.3 ± 3.1 c | 8.3 | 46.3 ± 4.2 b | 9.0 | 105.9 ± 7.7 a | 8.3 | 50.4 ± 4.0 b | 7.8 |
| Urea 1 | 90.6 ± 4.0 a | 4.4 | 43.4 ± 4.1 c | 9.5 | 42.3 ± 3.2 c | 7.6 | 64.8 ± 3.8 b | 5.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, C.; Selamat, J.; Jambari, N.N.; Sukor, R.; Murugesu, S.; Khatib, A. Muscle and Serum Metabolomics for Different Chicken Breeds under Commercial Conditions by GC–MS. Foods 2021, 10, 2174. https://doi.org/10.3390/foods10092174
Tan C, Selamat J, Jambari NN, Sukor R, Murugesu S, Khatib A. Muscle and Serum Metabolomics for Different Chicken Breeds under Commercial Conditions by GC–MS. Foods. 2021; 10(9):2174. https://doi.org/10.3390/foods10092174
Chicago/Turabian StyleTan, Chengkeng, Jinap Selamat, Nuzul Noorahya Jambari, Rashidah Sukor, Suganya Murugesu, and Alfi Khatib. 2021. "Muscle and Serum Metabolomics for Different Chicken Breeds under Commercial Conditions by GC–MS" Foods 10, no. 9: 2174. https://doi.org/10.3390/foods10092174
APA StyleTan, C., Selamat, J., Jambari, N. N., Sukor, R., Murugesu, S., & Khatib, A. (2021). Muscle and Serum Metabolomics for Different Chicken Breeds under Commercial Conditions by GC–MS. Foods, 10(9), 2174. https://doi.org/10.3390/foods10092174