
Enhanced Soluble Expression of Linoleic Acid Isomerase by Coordinated Regulation of Promoter and Fusion Tag in Escherichia coli

Abstract
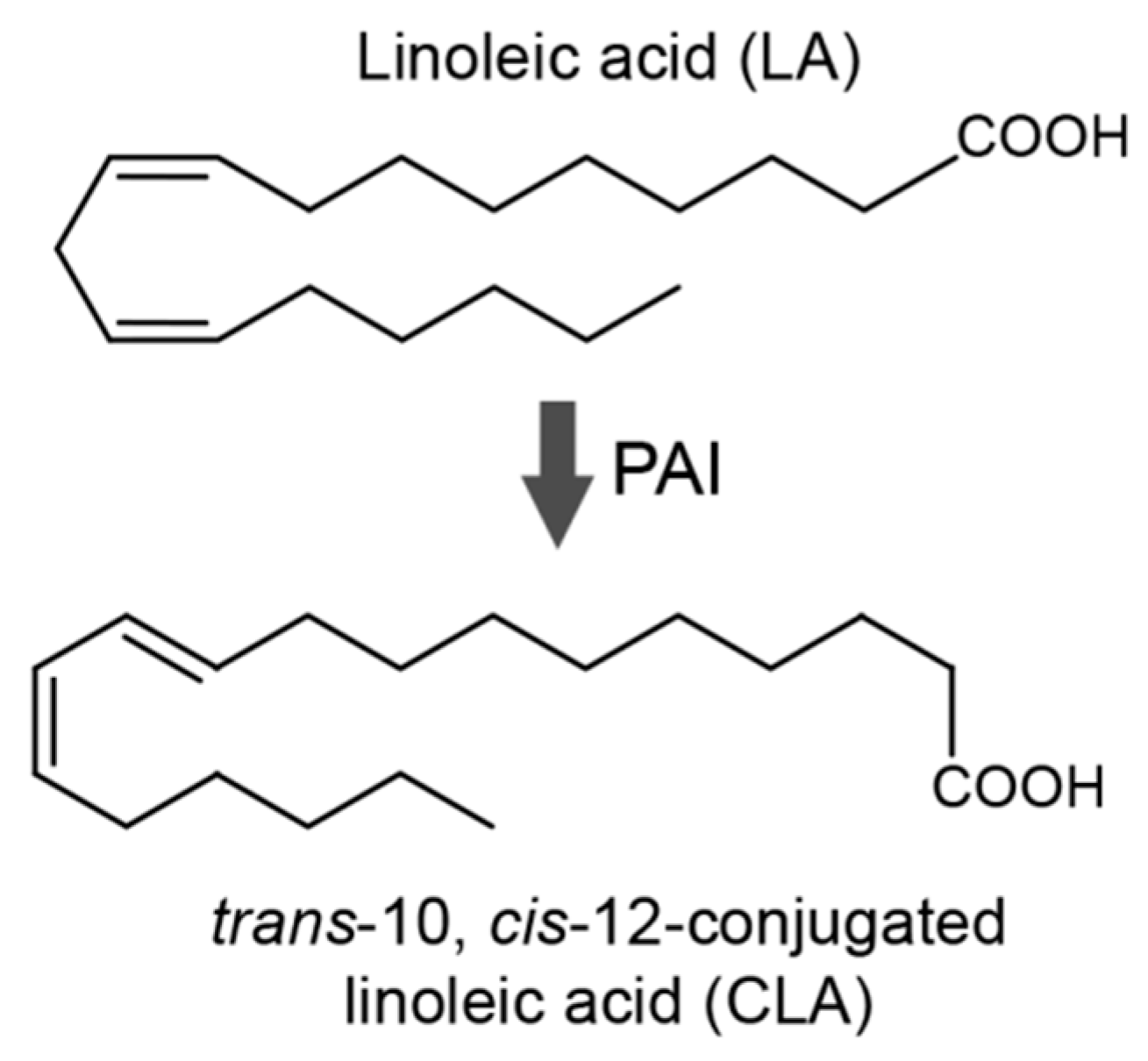
:1. Introduction
2. Materials and Methods
2.1. Materials
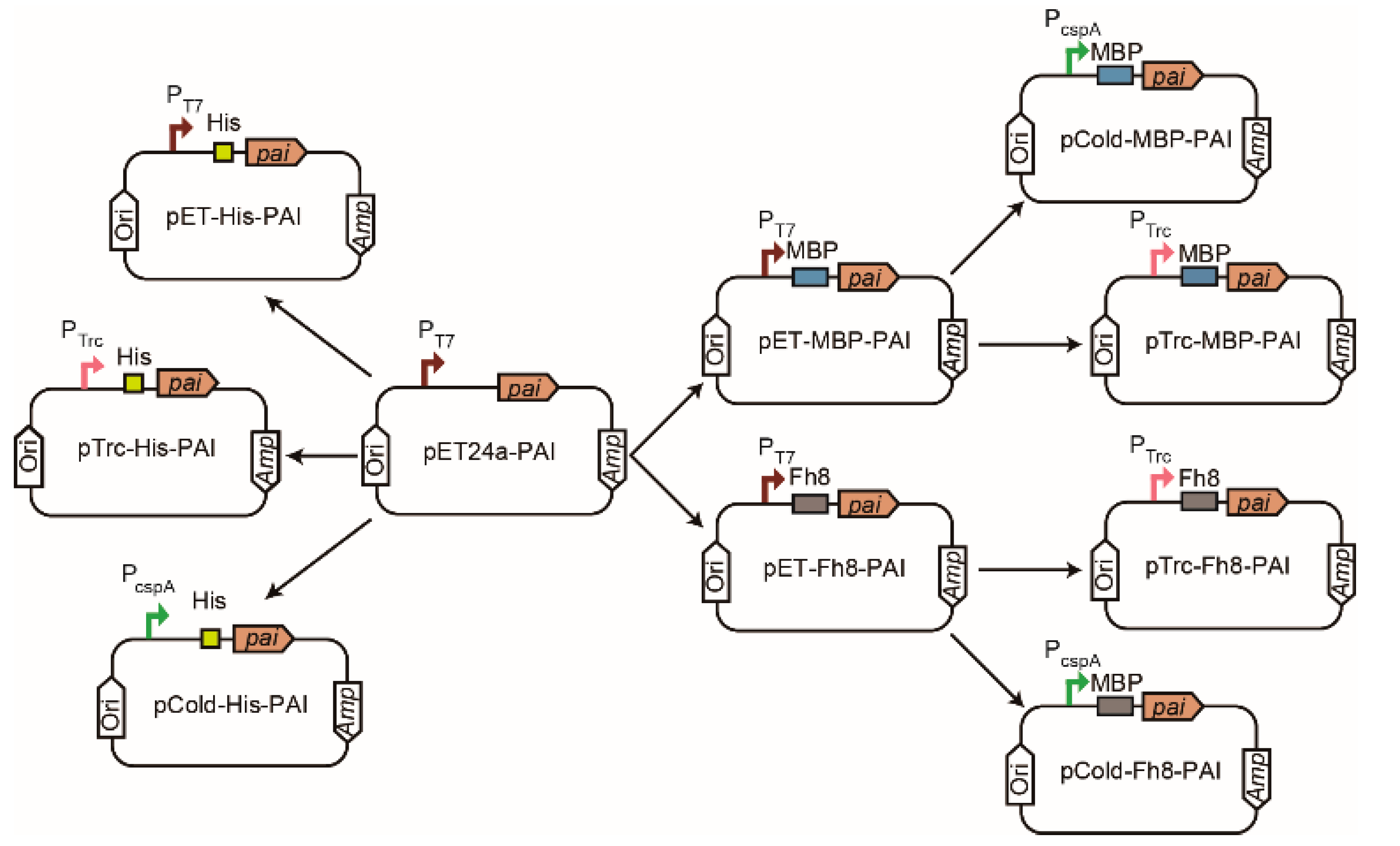
2.2. Construction of the Expression Vectors
2.3. Recombinant Protein Expression
2.4. SDS-PAGE Analysis of Recombinant Proteins
2.5. Measurement of Total PAI Activity with Cell Lysate
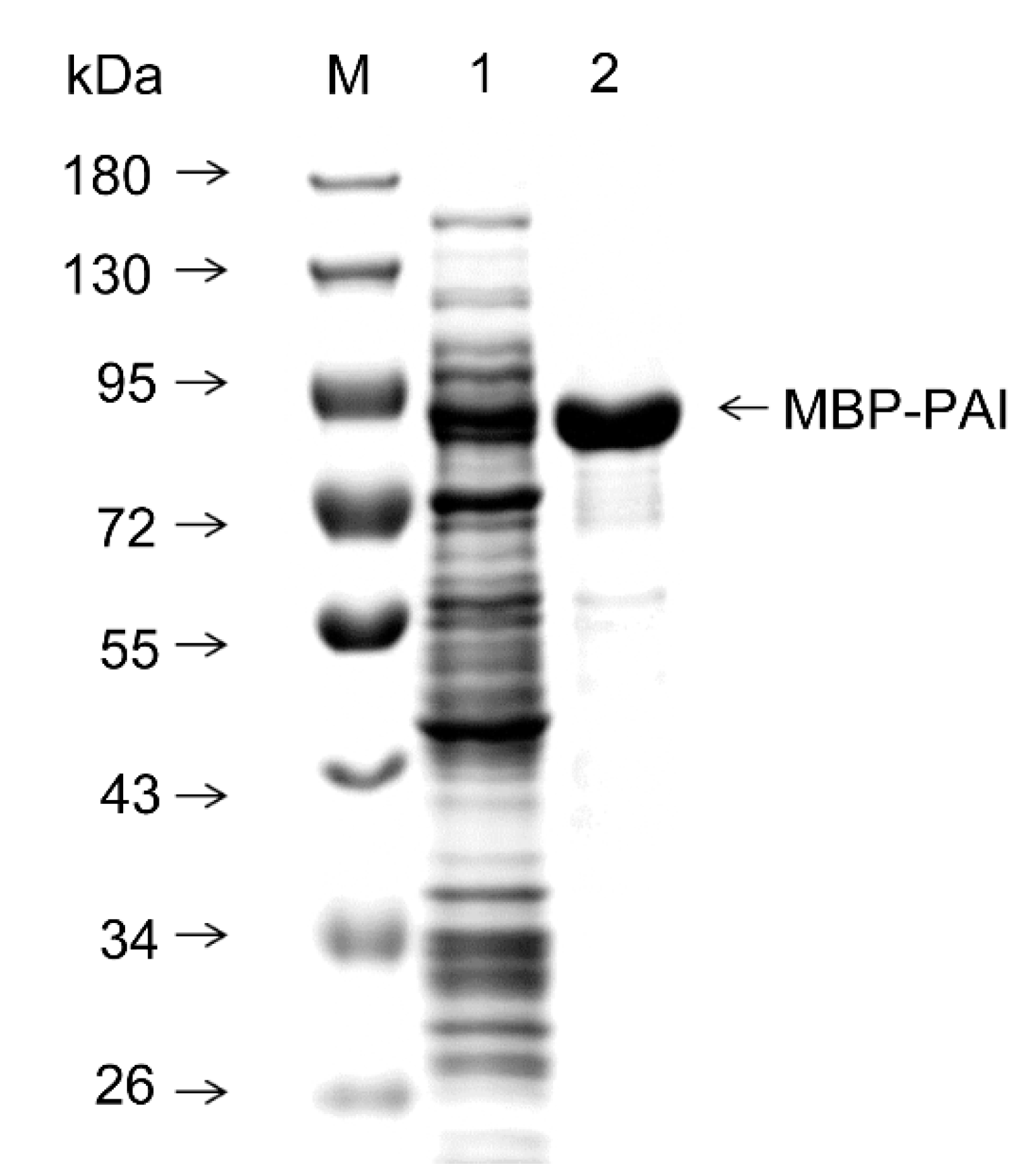
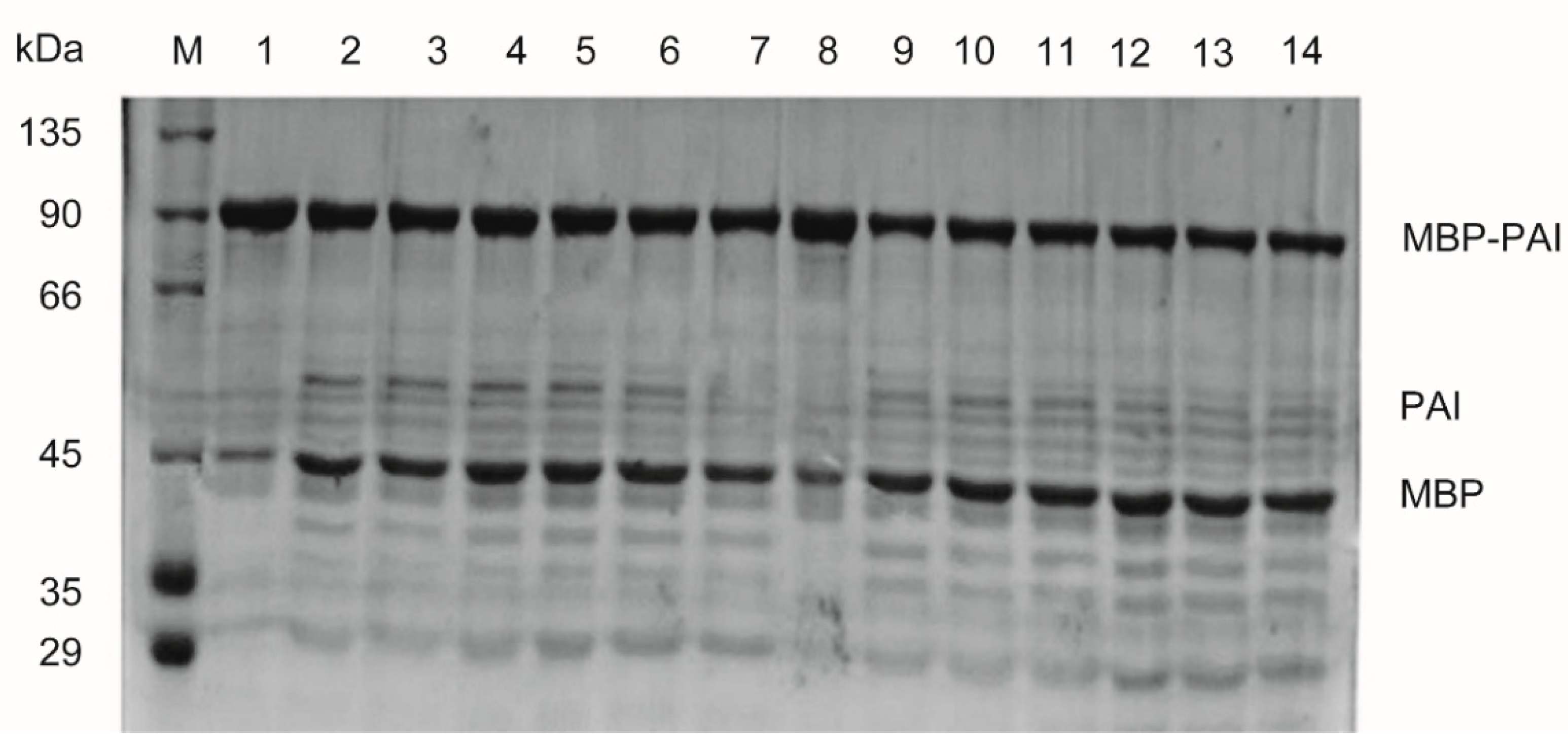
2.6. Purification of MBP-PAI and Separation of PAI from the Fusion Tag
2.7. Measurement of Purified MBP-PAI Activity
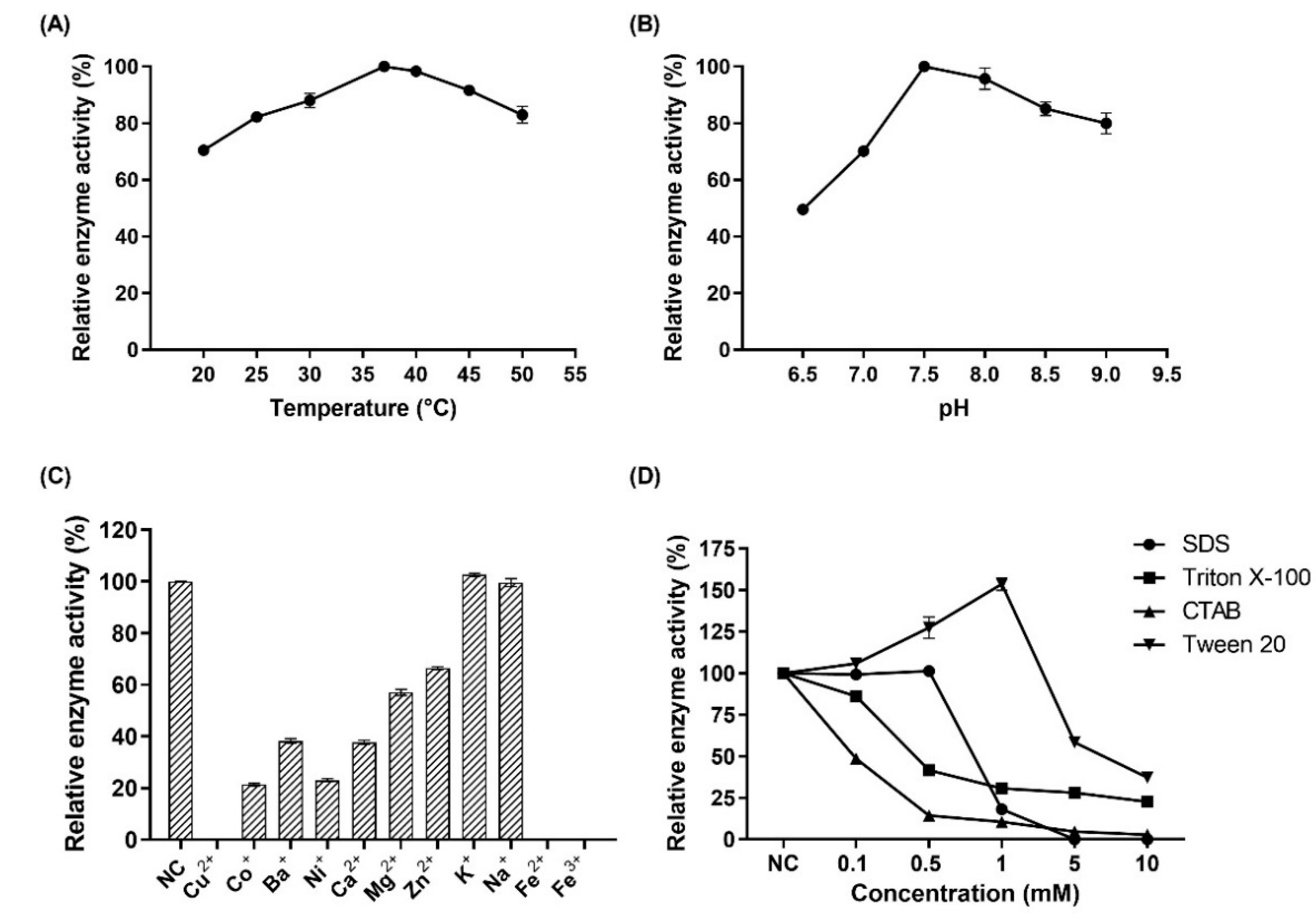
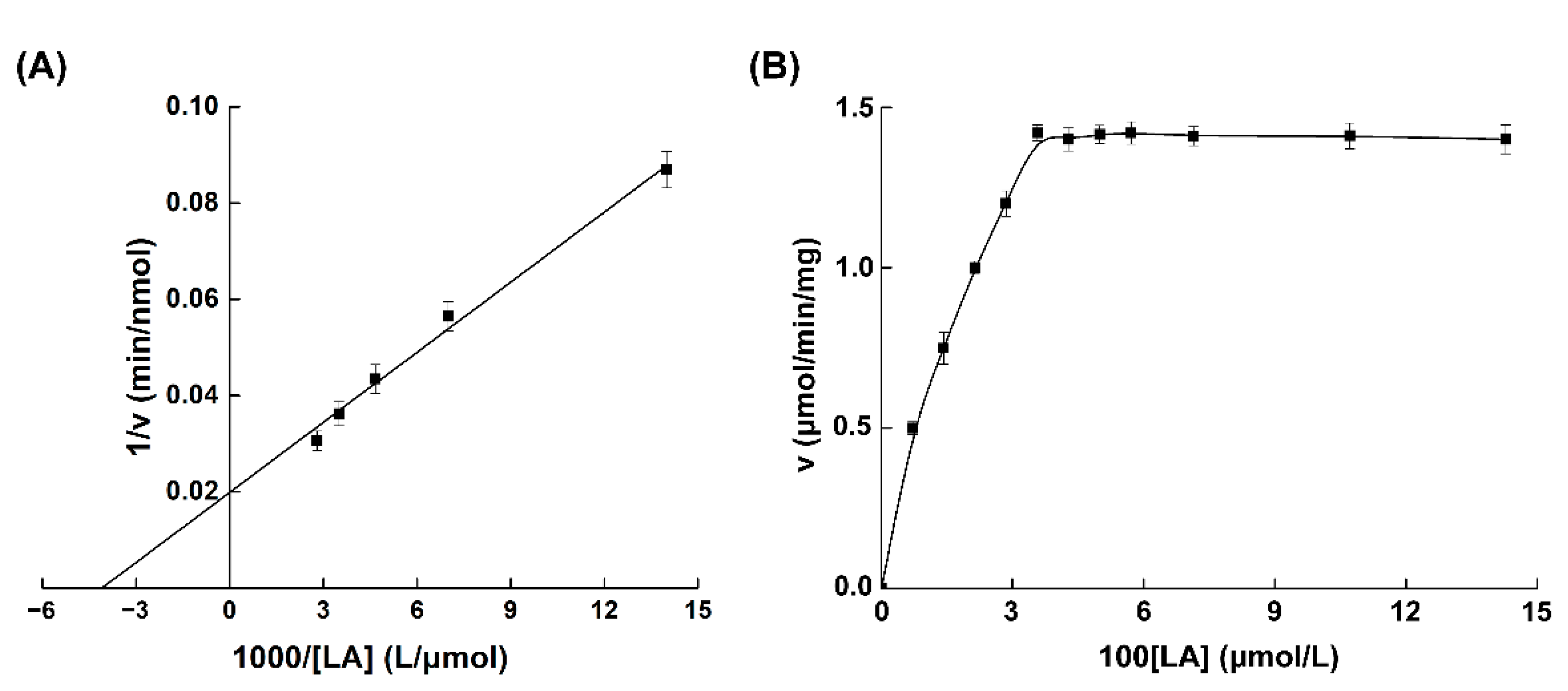
2.8. Analysis of the Enzymatic Properties of MBP-PAI
3. Results and Discussion
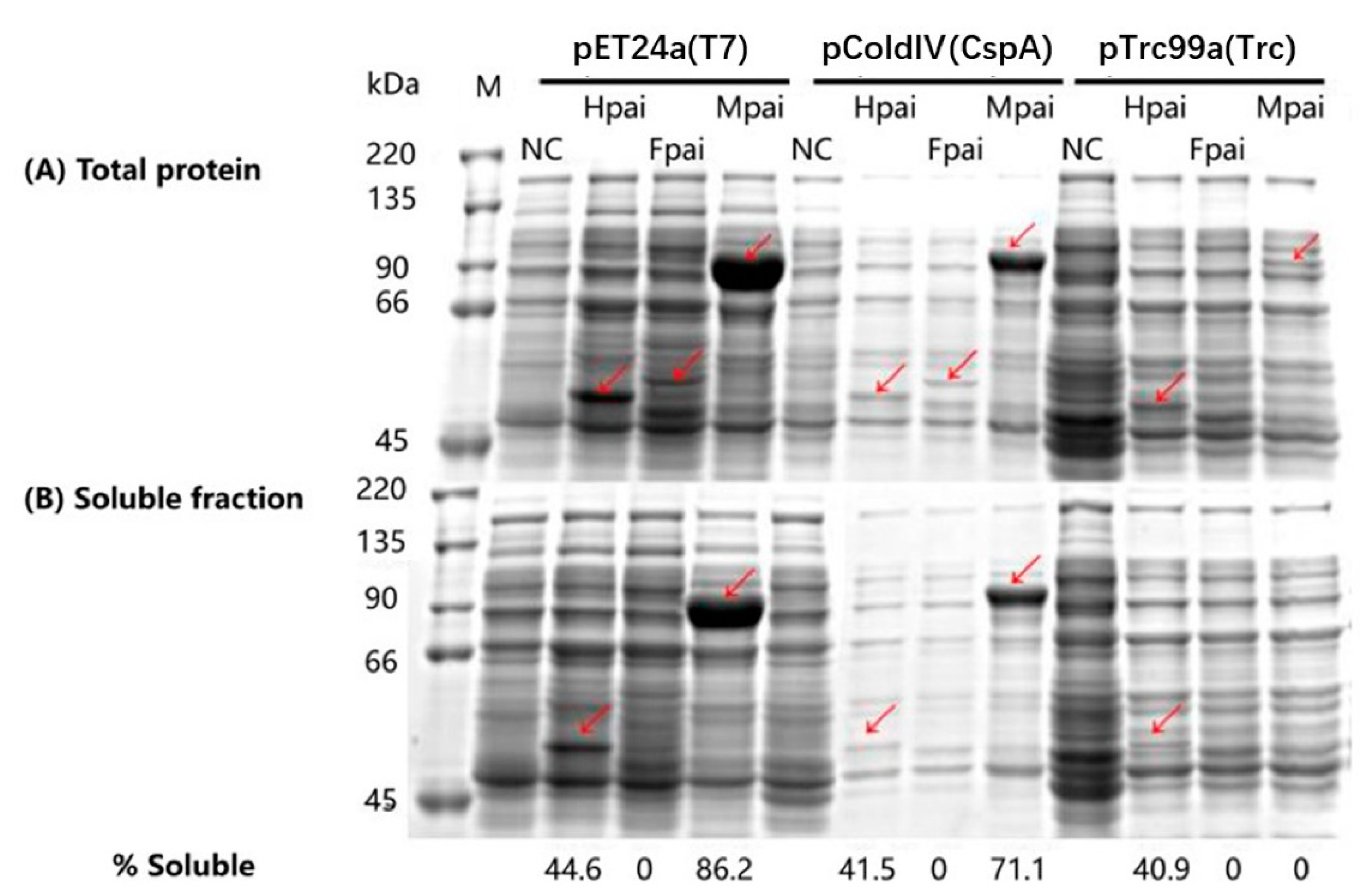
3.1. Effects of Promoters and Fusion Tags on PAI Expression and Solubility
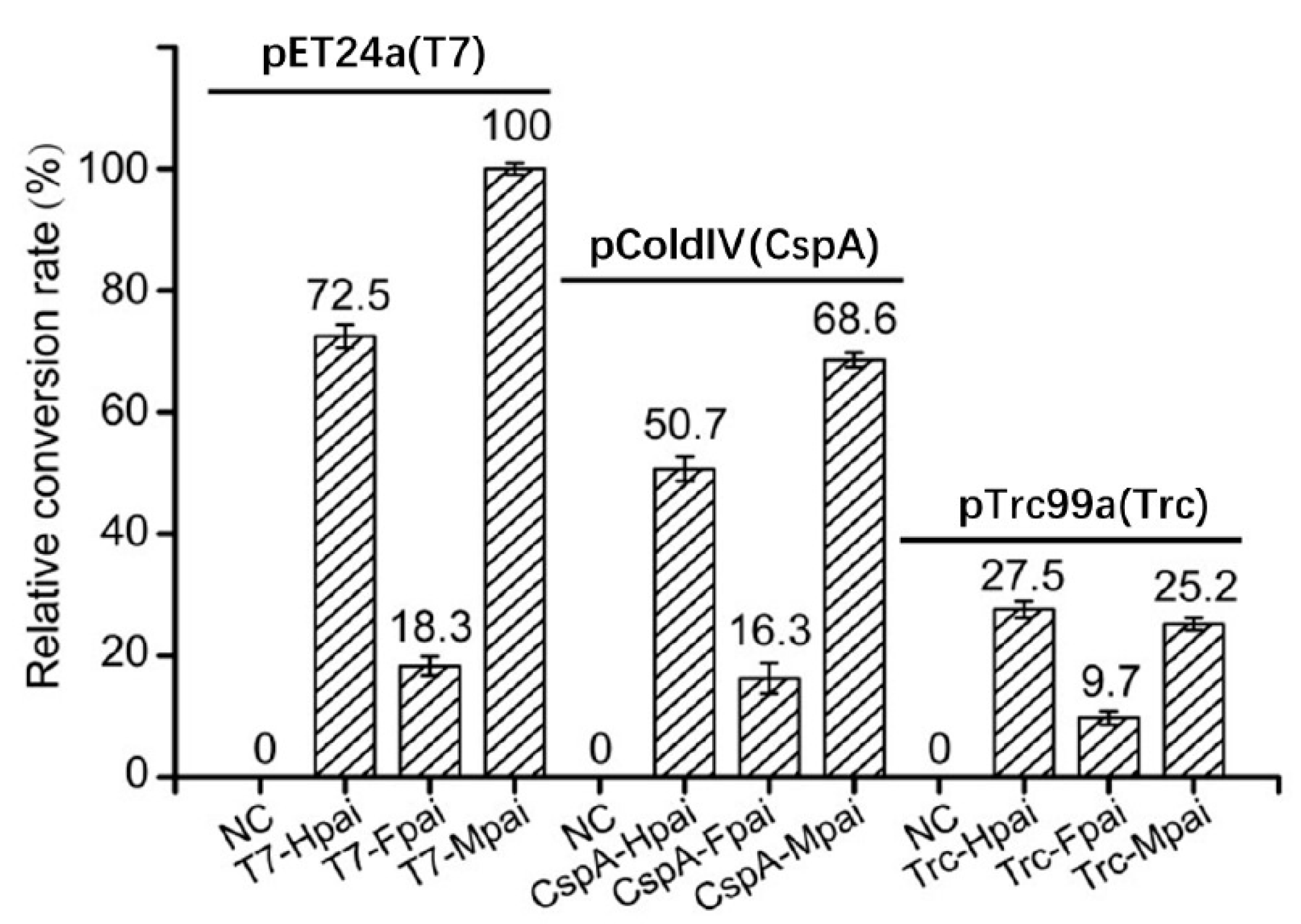
3.2. Effects of Promoters and Fusion Tags on Total Activity of Crude Enzyme Mixture
3.3. Purification and Cleavage of MBP-PAI by Factor Xa Protease
3.4. Characterization of MBP-PAI
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Den Hartigh, L.J. Conjugated Linoleic Acid Effects on Cancer, Obesity, and Atherosclerosis: A Review of Pre-Clinical and Human Trials with Current Perspectives. Nutrients 2019, 11, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.M.; McIntosh, M.K. Conjugated linoleic acid in humans: Regulation of adiposity and insulin sensitivity. J. Nutr. 2003, 133, 3041–3046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, Y.; Storkson, J.M.; Liu, W.; Albright, K.J.; Cook, M.E.; Pariza, M.W. Structure-activity relationship of conjugated linoleic acid and its cognates in inhibiting heparin-releasable lipoprotein lipase and glycerol release from fully differentiated 3T3-L1 adipocytes. J. Nutr. Biochem. 2004, 15, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Baldwin, J.; Collins, B.; Hixson, L.; Lee, K.T.; Herberg, T.; Starnes, J.; Cooney, P.; Chuang, C.C.; Hopkins, R.; et al. Low level of trans-10, cis-12 conjugated linoleic acid decreases adiposity and increases browning independent of inflammatory signaling in overweight Sv129 mice. J. Nutr. Biochem. 2015, 26, 616–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagao, K.; Inoue, N.; Wang, Y.M.; Yanagita, T. Conjugated linoleic acid enhances plasma adiponectin level and alleviates hyperinsulinemia and hypertension in Zucker diabetic fatty (fa/fa) rats. Biochem. Biophys. Res. Commun. 2003, 310, 562–566. [Google Scholar] [CrossRef]
- Mitchell, P.L.; Karakach, T.K.; Currie, D.L.; McLeod, R.S. t-10, c-12 CLA dietary supplementation inhibits atherosclerotic lesion development despite adverse cardiovascular and hepatic metabolic marker profiles. PLoS ONE 2012, 7, e52634. [Google Scholar] [CrossRef] [Green Version]
- McGowan, M.M.; Eisenberg, B.L.; Lewis, L.D.; Froehlich, H.M.; Wells, W.A.; Eastman, A.; Kuemmerle, N.B.; Rosenkrantz, K.M.; Barth, R.J., Jr.; Schwartz, G.N.; et al. A proof of principle clinical trial to determine whether conjugated linoleic acid modulates the lipogenic pathway in human breast cancer tissue. Breast Cancer Res. Treat. 2013, 138, 175–183. [Google Scholar] [CrossRef]
- Liavonchanka, A.; Hornung, E.; Feussner, I.; Rudolph, M.G. Structure and mechanism of the Propionibacterium acnes polyunsaturated fatty acid isomerase. Proc. Natl. Acad. Sci. USA 2006, 103, 2576–2581. [Google Scholar] [CrossRef] [Green Version]
- Hornung, E.; Krueger, C.; Pernstich, C.; Gipmans, M.; Porzel, A.; Feussner, I. Production of (10E, 12Z)-conjugated linoleic acid in yeast and tobacco seeds. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2005, 1738, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.D.; Grund, A.D.; Schneider, K.J.; Langley, K.M.; Wassink, S.L.; Peng, S.S.; Rosson, R.A. Linoleic acid isomerase from Propionibacterium acnes: Purification, characterization, molecular cloning, and heterologous expression. Appl. Biochem. Biotechnol. 2007, 143, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Farmani, J.; Roohvand, F.; Safari, M.; Aghasadeghi, M.R.; Razavi, S.H.; Motevalli, F. Cell surface display of Propionibacterium acnes linoleic acid isomerase by Pichia pastoris. Rom. Biotech. Lett. 2013, 18, 8307–8316. [Google Scholar]
- Zhang, B.X.; Rong, C.C.; Chen, H.Q.; Song, Y.D.; Zhang, H.; Chen, W. De novo synthesis of trans-10, cis-12 conjugated linoleic acid in oleaginous yeast Yarrowia Lipolytica. Microb. Cell Factories 2012, 11, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, D.H.; Chen, H.Q.; Hao, G.F.; Yang, B.; Zhang, B.X.; Zhang, H.; Chen, W.; Chen, Y.Q. Production of conjugated linoleic acid by heterologous expression of linoleic acid isomerase in oleaginous fungus Mortierella alpina. Biotechnol. Lett. 2015, 37, 1983–1992. [Google Scholar] [CrossRef] [PubMed]
- Baneyx, F.; Mujacic, M. Recombinant protein folding and misfolding in Escherichia coli. Nat. Biotechnol. 2004, 22, 1399–1408. [Google Scholar] [CrossRef] [PubMed]
- Esposito, D.; Chatterjee, D.K. Enhancement of soluble protein expression through the use of fusion tags. Curr. Opin. Biotechnol. 2006, 17, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Lu, X.; Zong, H.; Zhuge, B.; Shen, W. Bioproduction of trans-10,cis-12-Conjugated Linoleic Acid by a Highly Soluble and Conveniently Extracted Linoleic Acid Isomerase and an Extracellularly Expressed Lipase from Recombinant Escherichia coli Strains. J. Microbiol. Biotechnol. 2018, 28, 739–747. [Google Scholar] [CrossRef] [PubMed]
- Raran-Kurussi, S.; Waugh, D.S. The Ability to Enhance the Solubility of Its Fusion Partners Is an Intrinsic Property of Maltose-Binding Protein but Their Folding Is Either Spontaneous or Chaperone-Mediated. PLoS ONE 2012, 7, e49589. [Google Scholar] [CrossRef] [Green Version]
- Nagai, K.; Thøgersen, H. Generation of β-globin by sequence-specific proteolysis of a hybrid protein produced in Escherichia coli. Nature 1984, 309, 810–812. [Google Scholar] [CrossRef] [PubMed]
- Nagai, K.; Thgersen, H.C. Synthesis and sequence-specific proteolysis of hybrid proteins produced in Escherichia coli. Methods Enzymol. 1987, 153, 461–481. [Google Scholar] [PubMed]
- Biasutti, M.A.; Abuin, E.B.; Silber, J.J.; Correa, N.M.; Lissi, E.A. Kinetics of reactions catalyzed by enzymes in solutions of surfactants. Adv. Colloid Interface Sci. 2008, 136, 1–24. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Application Fragment |
|---|---|---|
| F1 | 5′-CGCCATATGCATCATCATCATCATCACATGTCCATCTCGAAGGATTCACG-3′ | Fragment NdeI-His-PAI- XhoI for pET24a(+) |
| R1 | 5′-CCGCTCGAGTTACACGAAGAACCGCGTCAC-3′ | |
| F2 | 5′-CGAGCTCCATCATCATCATCATCACATGTCCATCTCGAAGGATTCACG-3′ | Fragment SacI-His-PAI- HindIII for pCold IV and pTrc99a |
| R2 | 5′-CCCAAGCTTTTACACGAAGAACCGCGTCAC-3′ | |
| F3 | 5′-CTTTCAAATCAAGCTTATGAAAATCGAAGAAGGTAAAC-3′ | Fragment MBP-Factor Xa for pET24a(+) |
| R3 | 5′-ACAATTCCCCTCTAGTCACACGAAGAACCGCGTC-3′ | |
| F4 | 5′-CAAAATTATTTCTAGATGCCTAGTGTTCAAGAGG-3′ | Fragment Fh8-Factor Xa for pET24a(+) |
| R4 | 5′-CAAGGATGGCAAGCTCCTACCTTCGATCTATGATG-3′ | |
| F5 | 5′-CCTCGAGGGATCCGAATTCAATGAAAATCGAAGAAGGTAAACTGGTAATC-3′ | Fragment MBP-Factor Xa-PAI for pCold IV |
| R5 | 5′-TTTAAGCAGAGATTACCTATTCACACGAAGAACCGCGTC-3′ | |
| F6 | 5′-GCTGGTACCCGGGGATCCATGAAAATCGAAGAAGGTAAACTGGTAATC-3′ | Fragment MBP-Factor Xa-PAI for pTrc99a |
| R6 | 5′-CTCATCCGCCAAAACAGCCTCACACGAAGAACCGCGTC-3′ | |
| F7 | 5′-CCTCGAGGGATCCGAATTCATGCCTAGTGTTCAAGAGGTTG-3′ | Fragment Fh8-Factor Xa-PAI for pCold IV |
| R7 | 5′-TTTAATCATATATTACCTATTCACACGAAGAACCGCGTC-3′ | |
| F8 | 5′-GCTCGGTACCCGGGGATCCTATGCCTAGTGTTCAAGAGGTTG-3′ | Fragment Fh8-Factor Xa-PAI for pTrc99a |
| R8 | 5′-CTCATCCGCCAAAACAGAGCCATCACACGAAGAACCGCGTC-3′ |
| Expression Vectors | pET24a(+) | pColdIV | pTrc99a |
|---|---|---|---|
| OD600 | 0.6 | 0.6 | 0.6 |
| Final concentration of IPTG | 0.1 mM | 0.1 mM | 0.1 mM |
| Induction temperature | 20 °C | 15 °C | 37 °C |
| Induction time | 20 h | 24 h | 8 h |
| Step | Protein (mg) | Total Activity (nmol/min) | Specific Activity (nmol/min/mg) | Yield (%) | Purification (Fold) |
| Crude extract | 34.8 | 914 | 26.3 | 100 | 1 |
| Maltose-affinity chromatography | 0.461 | 498 | 1084 | 54.5 | 41 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, B.; Zhu, T.; Huang, X. Enhanced Soluble Expression of Linoleic Acid Isomerase by Coordinated Regulation of Promoter and Fusion Tag in Escherichia coli. Foods 2022, 11, 1515. https://doi.org/10.3390/foods11101515
Zhang B, Zhu T, Huang X. Enhanced Soluble Expression of Linoleic Acid Isomerase by Coordinated Regulation of Promoter and Fusion Tag in Escherichia coli. Foods. 2022; 11(10):1515. https://doi.org/10.3390/foods11101515
Chicago/Turabian StyleZhang, Baixi, Tong Zhu, and Xintian Huang. 2022. "Enhanced Soluble Expression of Linoleic Acid Isomerase by Coordinated Regulation of Promoter and Fusion Tag in Escherichia coli" Foods 11, no. 10: 1515. https://doi.org/10.3390/foods11101515
APA StyleZhang, B., Zhu, T., & Huang, X. (2022). Enhanced Soluble Expression of Linoleic Acid Isomerase by Coordinated Regulation of Promoter and Fusion Tag in Escherichia coli. Foods, 11(10), 1515. https://doi.org/10.3390/foods11101515