

Polymeric Compounds of Lingonberry Waste: Characterization of Antioxidant and Hypolipidemic Polysaccharides and Polyphenol-Polysaccharide Conjugates from Vaccinium vitis-idaea Press Cake



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Chemicals
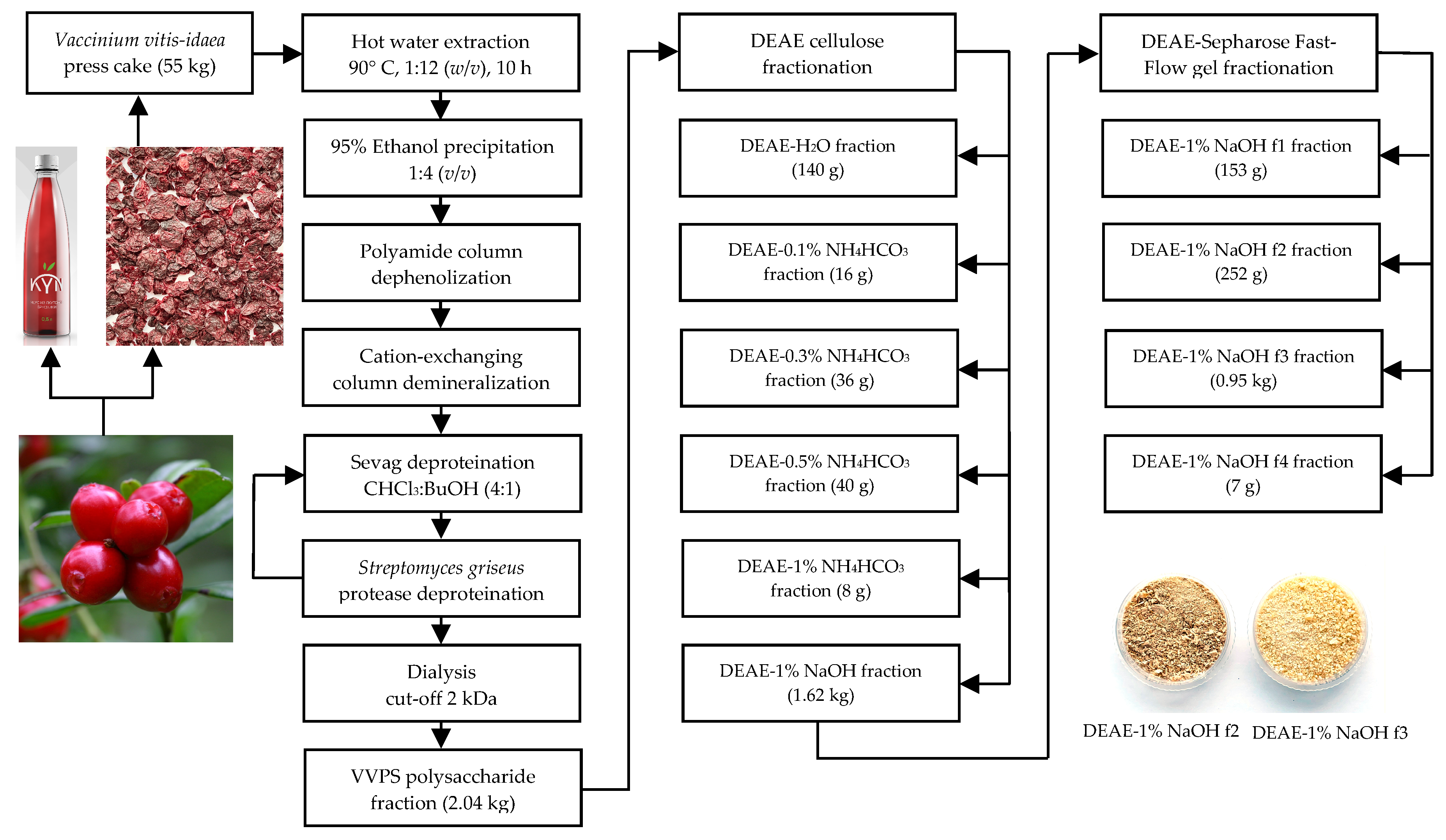
2.2. Polysaccharide Fraction of V. vitis-idaea Press Cake (VVPS) Isolation
2.3. Diethylaminoethyl–Cellulose (DEAE–Cellulose) Fractionation of VVPS
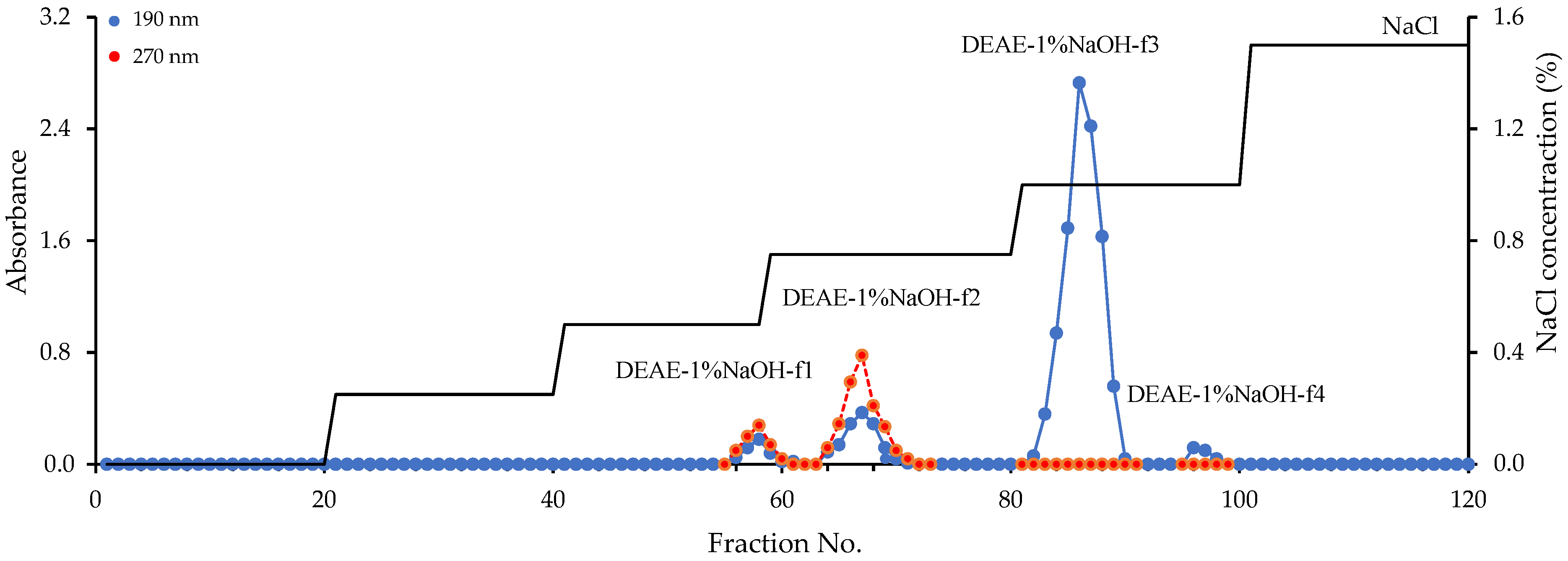
2.4. DEAE–Sepharose Fast-Flow Gel Fractionation of the DEAE–1% NaOH Fraction
2.5. Chemical Composition of VVPS and DEAE–Cellulose Fractions
2.6. Elemental Composition
2.7. Monosaccharide Composition
2.8. Ultraviolet–Visible (UV–Vis) Spectroscopy
2.9. Fourier-Transform Infrared (FTIR) Spectroscopy
2.10. Molecular Weight Determination
2.11. Linkage Analysis
2.12. Alkaline Destruction and Analysis of Degradation Products by High-Performance Liquid Chromatography with Photodiode Array Detection and Electrospray Ionization Triple Quadrupole Mass Spectrometric Detection (HPLC–PDA–ESI–tQMS)
2.13. Hydrolysis of DEAE–1% NaOH-f1 and DEAE–1% NaOH-f2 by 2% Oxalic Acid
2.14. Antioxidant Activity
2.15. Hypolipidemic Activity
2.15.1. In Vitro Assays
2.15.2. In Vivo Assays
2.16. Statistical and Multivariate Analysis
3. Results and Discussion
3.1. Yield and Chemical Composition of V. vitis-idaea Press Cake Polysaccharides (VVPS) and DEAE–Cellulose Fractions
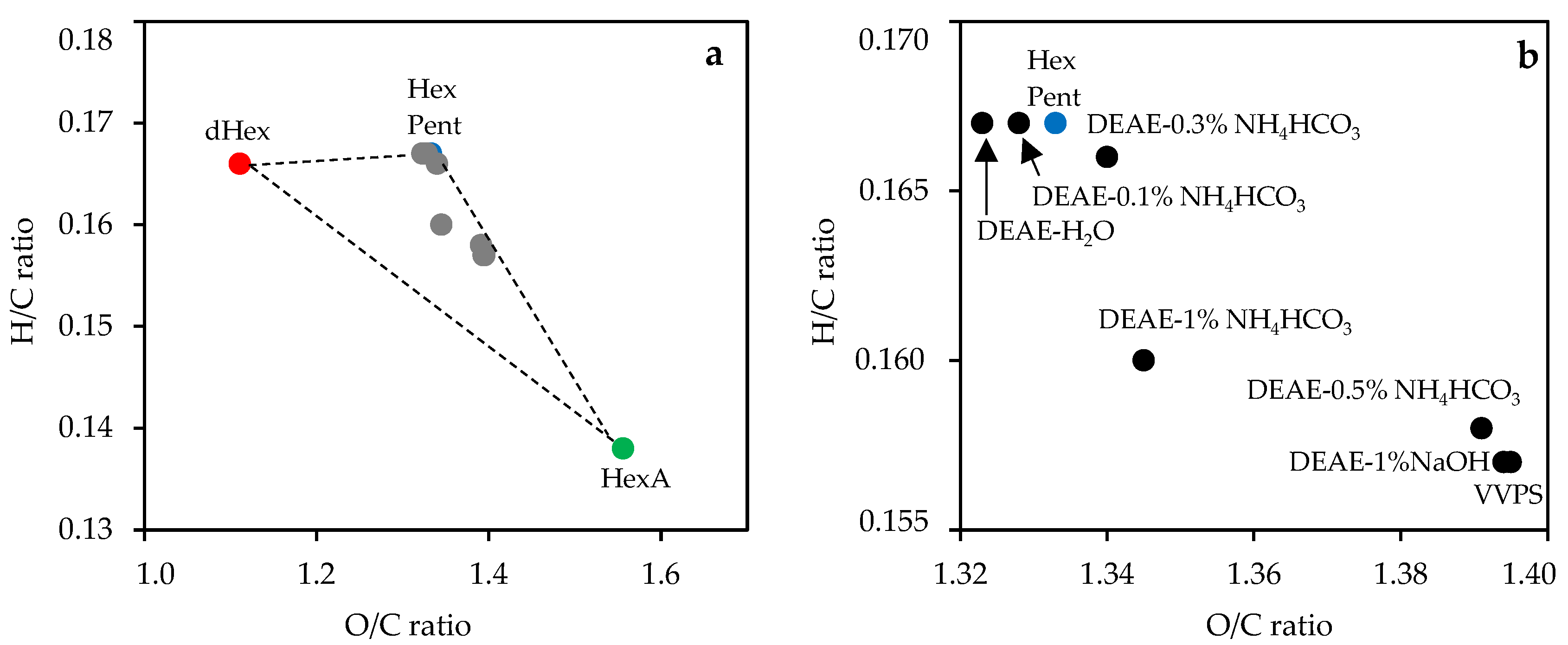
3.2. Elemental Composition
3.3. Monosaccharide Composition
3.4. Bioactivity of VVPS and DEAE–Cellulose Fractions of V. vitis-idaea Press Cake
3.4.1. Antioxidant Activity
3.4.2. In Vitro Hypolipidemic Activity
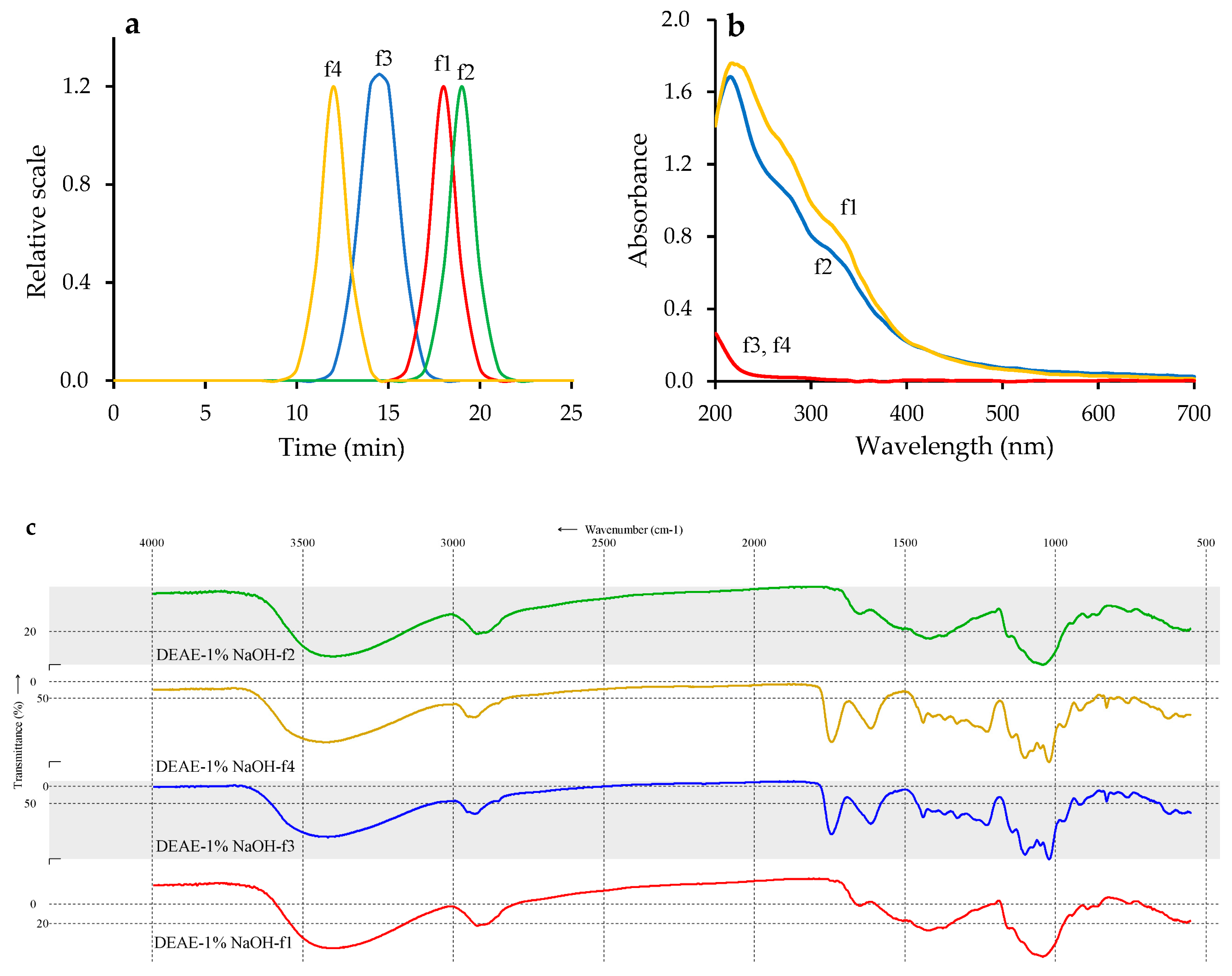
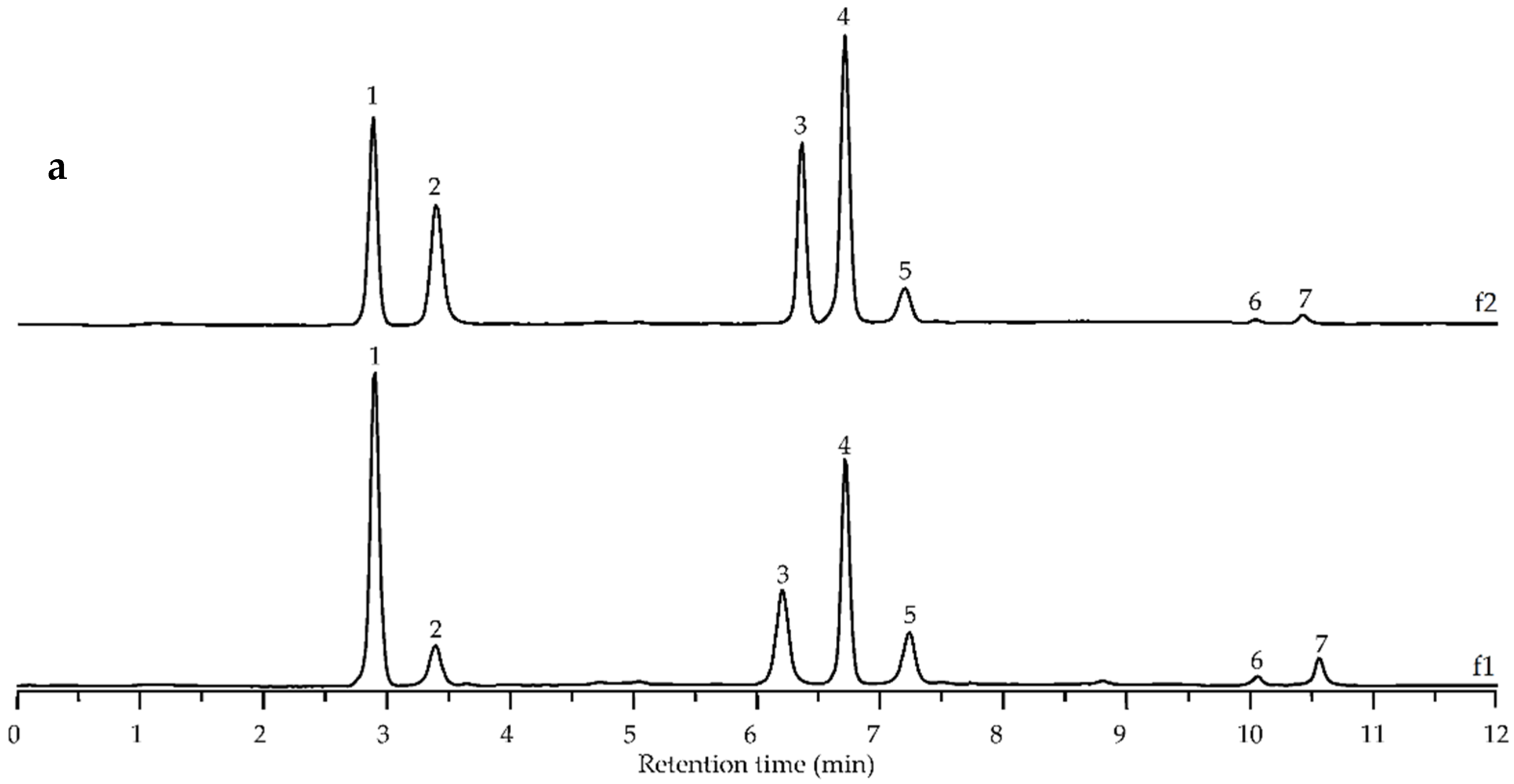
3.5. Preparative Chromatography of Fraction DEAE-1% NaOH and Characterization of Homogenous Polymers
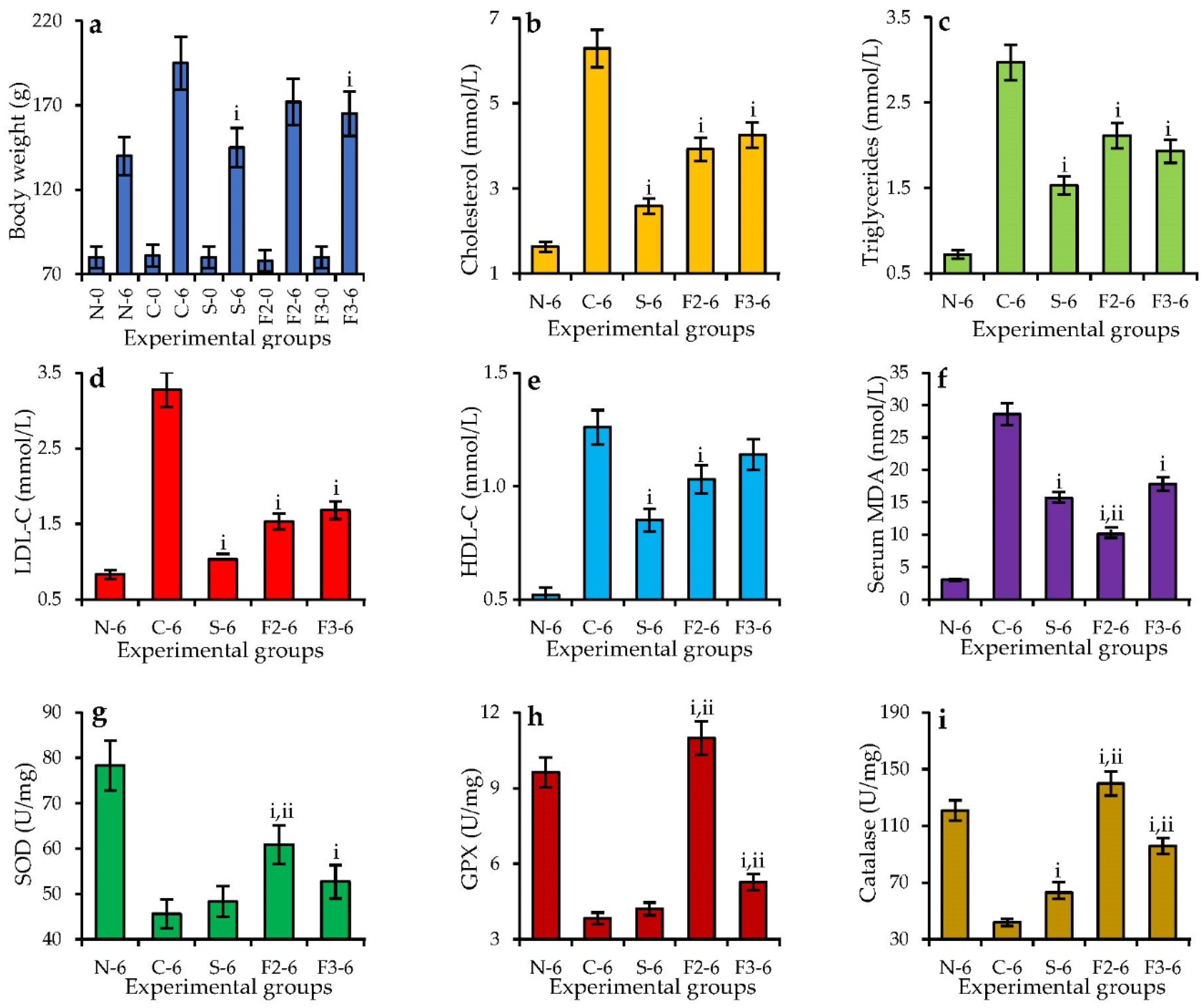
3.6. Antioxidant and Hypolipidemic Activity of Homogenic Polymers: In Vitro vs. In Vivo Assays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fierascu, R.C.; Fierascu, I.; Avramescu, S.M.; Sieniawska, E. Recovery of natural antioxidants from agro-industrial side streams through advanced extraction techniques. Molecules 2019, 24, 4212. [Google Scholar] [CrossRef] [PubMed]
- Maraveas, C. Production of sustainable and biodegradable polymers from agricultural waste. Polymers 2020, 12, 1127. [Google Scholar] [CrossRef] [PubMed]
- Córdova, A.; Henríquez, P.; Nuñez, H.; Rico-Rodriguez, F.; Guerrero, C.; Astudillo-Castro, C.; Illanes, A. Recent advances in the application of enzyme processing assisted by ultrasound in agri-foods: A review. Catalysts 2022, 12, 107. [Google Scholar] [CrossRef]
- Šelo, G.; Planinić, M.; Tišma, M.; Tomas, S.; Koceva Komlenić, D.; Bucić-Kojić, A. A comprehensive review on valorization of agro-food industrial residues by solid-state fermentation. Foods 2021, 10, 927. [Google Scholar] [CrossRef]
- Bujor, O.-C.; Tanase, C.; Popa, M.E. Phenolic antioxidants in aerial parts of wild Vaccinium species: Towards pharmaceutical and biological properties. Antioxidants 2019, 8, 649. [Google Scholar] [CrossRef]
- Malyshev, L.I. Flora of Siberia: Pyrolaceae-Lamiaceae; CRC Press: Boca Raton, FL, USA, 2006; pp. 25–28. [Google Scholar]
- Timoshok, E.E.; Skorokhodov, S.N. Assessment of berry resources of Ericaceae family in Tomsk region, their rational use and protection. Siberian Forest J. 2019, 4, 80–88. [Google Scholar] [CrossRef]
- Kowalska, K. Lingonberry (Vaccinium vitis-idaea L.) fruit as a source of bioactive compounds with health-promoting effects—A review. Int. J. Mol. Sci. 2021, 22, 5126. [Google Scholar] [CrossRef]
- Vilas-Boas, A.A.; Pintado, M.; Oliveira, A.L.S. Natural bioactive compounds from food waste: Toxicity and safety concerns. Foods 2021, 10, 1564. [Google Scholar] [CrossRef]
- Baroi, A.M.; Popitiu, M.; Fierascu, I.; Sărdărescu, I.-D.; Fierascu, R.C. Grapevine wastes: A rich source of antioxidants and other biologically active compounds. Antioxidants 2022, 11, 393. [Google Scholar] [CrossRef]
- Moccia, F.; Flores-Gallegos, A.C.; Chávez-González, M.L.; Sepúlveda, L.; Marzorati, S.; Verotta, L. Ellagic acid recovery by solid state fermentation of pomegranate wastes by Aspergillus niger and Saccharomyces cerevisiae: A comparison. Molecules 2019, 24, 3689. [Google Scholar] [CrossRef] [Green Version]
- Vu, H.T.; Scarlett, C.J.; Vuong, Q.V. Optimization of ultrasound-assisted extraction conditions for recovery of phenolic compounds and antioxidant capacity from banana (Musa cavendish) peel. J. Food Process. Preserv. 2017, 41, e13148. [Google Scholar] [CrossRef]
- Markhali, F.S.; Teixeira, J.A.; Rocha, C.M.R. Olive tree leaves—A source of valuable active compounds. Processes 2020, 8, 1177. [Google Scholar] [CrossRef]
- Boukroufa, M.; Boutekedjiret, C.; Petigny, L.; Rakotomanomana, N.; Chemat, F. Bio-refinery of orange peels waste: A new concept based on integrated green and solvent free extraction processes using ultrasound and microwave techniques to obtain essential oil, polyphenols and pectin. Ultrason. Sonochem. 2015, 24, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Ferrentino, G.; Morozova, K.; Mosibo, O.K.; Ramezani, M.; Scampicchio, M. Biorecovery of antioxidants from apple pomace by supercritical fluid extraction. J. Clean Prod. 2018, 186, 253–261. [Google Scholar] [CrossRef]
- Panzella, L.; Moccia, F.; Nasti, R.; Marzorati, S.; Verotta, L.; Napolitano, A. Bioactive phenolic compounds from agri-food wastes: An update on green and sustainable extraction methodologies. Front. Nutr. 2020, 7, 60. [Google Scholar] [CrossRef] [PubMed]
- Zokaityte, E.; Lele, V.; Starkute, V.; Zavistanaviciute, P.; Cernauskas, D.; Klupsaite, D.; Ruzauskas, M.; Alisauskaite, J.; Baltrusaitytė, A.; Dapsas, M.; et al. Antimicrobial, antioxidant, sensory properties, and emotions induced for the consumers of nutraceutical beverages developed from technological functionalised food industry by-products. Foods 2020, 9, 1620. [Google Scholar] [CrossRef] [PubMed]
- Pukalskienė, M.; Pukalskas, A.; Dienaitė, L.; Revinytė, S.; Pereira, C.V.; Matias, A.A.; Venskutonis, P.R. Recovery of bioactive compounds from strawberry (Fragaria × ananassa) pomace by conventional and pressurized liquid extraction and assessment their bioactivity in human cell cultures. Foods 2021, 10, 1780. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; Veberic, R.; Hudina, M.; Zorenc, Z.; Koron, D.; Senica, M. Fruit quality characteristics and biochemical composition of fully ripe blackberries harvested at different times. Foods 2021, 10, 1581. [Google Scholar] [CrossRef]
- Jurevičiūtė, I.; Keršienė, M.; Bašinskienė, L.; Leskauskaitė, D.; Jasutienė, I. Characterization of berry pomace powders as dietary fiber-rich food ingredients with functional properties. Foods 2022, 11, 716. [Google Scholar] [CrossRef]
- Struck, S.; Plaza, M.; Turner, C.; Rohm, H. Berry pomace–A review of processing and chemical analysis of its polyphenols. Int. J. Food Sci. Technol. 2016, 51, 1305–1318. [Google Scholar] [CrossRef]
- Souza, M.A.d.; Vilas-Boas, I.T.; Leite-da-Silva, J.M.; Abrahão, P.d.N.; Teixeira-Costa, B.E.; Veiga-Junior, V.F. Polysaccharides in agro-industrial biomass residues. Polysaccharides 2022, 3, 95–120. [Google Scholar] [CrossRef]
- Kitrytė, V.; Kavaliauskaitė, A.; Tamkutė, L.; Pukalskienė, M.; Syrpas, M.; Rimantas Venskutonis, P. Zero waste biorefining of lingonberry (Vaccinium vitis-idaea L.) pomace into functional ingredients by consecutive high pressure and enzyme assisted extractions with green solvents. Food Chem. 2020, 322, 126767. [Google Scholar] [CrossRef] [PubMed]
- Kunrade, L.; Rembergs, R.; Jēkabsons, K.; Kļaviņš, L.; Kļaviņš, M.; Muceniece, R.; Riekstiņa, U. Inhibition of NF-κB pathway in LPS-stimulated THP-1 monocytes and COX-2 activity in vitro by berry pomace extracts from five Vaccinium species. J. Berry Res. 2020, 10, 381–396. [Google Scholar] [CrossRef]
- Kahlon, T.S.; Smith, G.E. In vitro binding of bile acids by blueberries (Vaccinium spp.), plums (Prunus spp.), prunes (Prunus spp.), strawberries (Fragaria × ananassa), cherries (Malpighia punicifolia), cranberries (Vaccinium macrocarpon) and apples (Malus sylvestris). Food Chem. 2007, 100, 1182–1187. [Google Scholar] [CrossRef]
- Shamilov, A.A.; Bubenchikova, V.N.; Chernikov, M.V.; Pozdnyakov, D.I.; Garsiya, E.R. Vaccinium vitis-idaea L.: Chemical contents, pharmacological activities. Pharm. Sci. 2020, 26, 344–362. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Shamilov, A.A. Catechin-O-rhamnosides from Vaccinium vitis-idaea stems. Chem. Nat. Comp. 2022, 58, 269–273. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Shamilov, A.A. New compounds from Vaccinium vitis-idaea. Chem. Nat. Comp. 2022, 58, 240–244. [Google Scholar] [CrossRef]
- Ross, K.A.; Godfrey, D.; Fukumoto, L. The chemical composition, antioxidant activity and α-glucosidase inhibitory activity of water-extractable polysaccharide conjugates from northern Manitoba lingonberry. Cogent Food Agricult. 2015, 1, 1109781. [Google Scholar] [CrossRef]
- Luo, Q.; Cai, Y.; Yan, J.; Sun, M.; Corke, H. Hypoglycemic and hypolipidemic effects and antioxidant activity of fruit extracts from Lycium barbarum. Life Sci. 2004, 76, 137–149. [Google Scholar] [CrossRef]
- Quesada-Morua, M.S.; Hidalgo, O.; Morera, J.; Rojas, G.; Pérez, A.M.; Vaillant, F.; Fonseca, L. Hypolipidaemic, hypoglycaemic and antioxidant effects of a tropical highland blackberry beverage consumption in healthy individuals on a high-fat, high-carbohydrate diet challenge. J. Berry Res. 2020, 10, 459–474. [Google Scholar] [CrossRef]
- Yu, C.H.; Dai, X.Y.; Chen, Q.; Zang, J.N.; Deng, L.L.; Liu, Y.H.; Ying, H.Z. Hypolipidemic and antioxidant activities of polysaccharides from Rosae Laevigatae Fructus in rats. Carbohydr. Polym. 2013, 94, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Lamport, D.T.A.; Kieliszewski, M.J.; Showalter, A.M. Salt stress upregulates periplasmic arabinogalactan proteins: Using salt stress to analyse AGP function. New Phytol. 2006, 169, 479–492. [Google Scholar] [CrossRef] [PubMed]
- Thiex, N.; Novotny, L.; Crawford, A. Determination of ash in animal Feed: AOAC Official Method 942.05 revisited. J. AOAC Int. 2012, 95, 1392–1397. [Google Scholar] [CrossRef] [PubMed]
- Olennikov, D.N.; Kashchenko, N.I.; Chirikova, N.K.; Gornostai, T.G.; Selyutina, I.Y.; Zilfikarov, I.N. Effect of low temperature cultivation on the phytochemical profile and bioactivity of Arctic plants: A case of Dracocephalum palmatum. Int. J. Molec. Sci. 2017, 18, 2579. [Google Scholar] [CrossRef]
- Needs, P.W.; Selvendran, R.R. Avoiding oxidative degradation during sodium hydroxide/methyl iodide-mediated carbohydrate methylation in dimethyl sulfoxide. Carbohydr. Res. 1993, 245, 1–10. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Agafonova, S.V.; Rokhin, A.V.; Penzina, T.A.; Borovskii, G.B. Branched glucan from the fruiting bodies of Piptoporus betulinus (Bull.:Fr) Karst. Appl. Biochem. Microbiol. 2011, 48, 65–70. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kirillina, C.S.; Chirikova, N.K. Water-soluble melanoidin pigment as a new antioxidant component of fermented willowherb leaves (Epilobium angustifolium). Antioxidants 2021, 10, 1300. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Chirikova, N.K.; Vasilieva, A.G.; Fedorov, I.A. LC-MS profile, gastrointestinal and gut microbiota stability and antioxidant activity of Rhodiola rosea herb metabolites: A comparative study with subterranean organs. Antioxidants 2020, 9, 526. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Chirikova, N.K.; Okhlopkova, Z.M.; Zulfugarov, I.S. Chemical composition and antioxidant activity of Tánara Ótó (Dracocephalum palmatum Stephan), a medicinal plant used by the North-Yakutian nomads. Molecules 2013, 18, 14105–14121. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Vasilieva, A.G.; Chirikova, N.K. Fragaria viridis fruit metabolites: Variation of LC-MS profile and antioxidant potential during ripening and storage. Pharmaceuticals 2020, 13, 262. [Google Scholar] [CrossRef]
- Kumar, S.; Kumar, D.; Jusha, M.; Saroha, K.; Singif, N.; Vashishta, B. Antioxidant and free radical scavenging potential of Citrullus colocynthis (L.) Schrad. methanolic fruit extract. Acta Pharm. 2008, 58, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Fernando, C.D.; Soysa, P. Optimized enzymatic colorimetric assay for determination of hydrogen peroxide (H2O2) scavenging activity of plant extracts. MethodsX 2015, 2, 283–291. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kashchenko, N.I.; Chirikova, N.K. A novel HPLC-assisted method for investigation of the Fe2+-chelating activity of flavonoids and plant extracts. Molecules 2014, 19, 18296–18316. [Google Scholar] [CrossRef]
- Nagaoka, S.; Futamura, Y.; Miwa, K.; Awano, T.; Yamauchi, K.; Kanamaru, Y.; Kuwata, T. Identification of novel hypocholesterolemic peptides derived from bovine milk β-lactoglobulin. Biochem. Biophys. Res. Commun. 2001, 281, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; White, P.J. In vitro bile-acid binding and fermentation of high, medium, and low molecular weight β-glucan. J. Agricult. Food Chem. 2010, 58, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.; Yu, H.; Wang, X.; Li, K.; Li, P. Effect of the molecular weight of water-soluble chitosan on its fat-/cholesterol-binding capacities and inhibitory activities to pancreatic lipase. PeerJ 2017, 5, e3279. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Chang, S.K.C.; Zhang, Y. Comparison of α-amylase, α-glucosidase and lipase inhibitory activity of the phenolic substances in two black legumes of different genera. Food Chem. 2017, 214, 259–268. [Google Scholar] [CrossRef]
- Cheng, Y.; Tang, K.; Wu, S.; Liu, L.; Qiang, C.; Lin, X.; Liu, B. Astragalus polysaccharides lowers plasma cholesterol through mechanisms distinct from statins. PLoS ONE 2011, 6, e27437. [Google Scholar] [CrossRef]
- Deng, J.; Shi, Z.-J.; Li, X.-Z.; Liu, H.-M. Soluble polysaccharides isolation and characterization from rabbiteye blueberry (Vaccinium ashei) fruits. BioRes 2013, 8, 405–419. [Google Scholar] [CrossRef]
- Rivas-Ubach, A.; Liu, Y.; Bianchi, T.S.; Tolić, N.; Jansson, C.; Paša-Tolić, L. Moving beyond the van Krevelen diagram: A new stoichiometric approach for compound classification in organisms. Anal. Chem. 2018, 90, 6152–6160. [Google Scholar] [CrossRef]
- Jooyandeh, H.; Noshad, M.; Khamirian, R.A. Modeling of ultrasound-assisted extraction, characterization and in vitro pharmacological potential of polysaccharides from Vaccinium arctostaphylos L. Int. J. Biol. Macromol. 2018, 107, 938–948. [Google Scholar] [CrossRef] [PubMed]
- Hilz, H.; Bakx, E.J.; Schols, H.A.; Voragen, A.G.J. Cell wall polysaccharides in black currants and bilberries—characterisation in berries, juice, and press cake. Carbohydr. Polym. 2005, 59, 477–488. [Google Scholar] [CrossRef]
- Li, X.; Wang, X.; Guo, X.; Li, D.; Huo, J.; Yu, Z. Structural and biochemical characterization of a polysaccharide isolated from Vaccinium uliginosum L. Starch 2022, 74, 2100109. [Google Scholar] [CrossRef]
- Wang, J.; Hu, S.; Nie, S.; Yu, Q.; Xie, M. Reviews on mechanisms of in vitro antioxidant activity of polysaccharides. Oxidative Med. Cell. Longev. 2016, 2016, 5692852. [Google Scholar] [CrossRef] [PubMed]
- Brouns, F.; Theuwissen, E.; Adam, A.; Bell, M.; Berger, A.; Mensink, R.P. Cholesterol-lowering properties of different pectin types in mildly hyper-cholesterolemic men and women. Eur. J. Clin. Nutr. 2012, 66, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Jin, M.; Liu, H. Water-soluble polysaccharide from dried Lycium barbarum fruits: Isolation, structural features and antioxidant activity. Carbohydr. Polym. 2011, 83, 1947–1951. [Google Scholar] [CrossRef]
- Ross, K.; Siow, Y.; Brown, D.; Isaak, C.; Fukumoto, L.; Godfrey, D. Characterization of water extractable crude polysaccharides from cherry, raspberry, and ginseng berry fruits: Chemical composition and bioactivity. Int. J. Food Prop. 2015, 18, 670–689. [Google Scholar] [CrossRef]
- Wu, C.; Zhao, M.; Bu, X.; Qing, Z.; Wang, L.; Xu, Y.; Bai, J. Preparation, characterization, antioxidant and antiglycation activities of selenized polysaccharides from blackcurrant. RSC Adv. 2020, 10, 32616–32627. [Google Scholar] [CrossRef]
- Scaldaferri, F.; Pizzoferrato, M.; Ponziani, F.R.; Gasbarrini, G.; Gasbarrini, A. Use and indications of cholestyramine and bile acid sequestrants. Intern. Emerg. Med. 2013, 8, 205–210. [Google Scholar] [CrossRef]
- Kahlon, T.S.; Smith, G.E. In vitro binding of bile acids by bananas, peaches, pineapple, grapes, pears, apricots and nectarines. Food Chem. 2007, 101, 1046–1051. [Google Scholar] [CrossRef]
- Ren, D.; Chen, G. Extraction, purification and bile acid-binding capacity in vitro of polysaccharides from Okra. Food Sci. 2010, 31, 110–113. [Google Scholar] [CrossRef]
- Long, H.; Xia, X.; Liao, S.; Wu, T.; Wang, L.; Chen, Q.; Wei, S.; Gu, X.; Zhu, Z. Physicochemical characterization and antioxidant and hypolipidaemic activities of a polysaccharide from the fruit of Kadsura coccinea (Lem.) A.C.Smith. Front. Nutr. 2022, 9, 903218. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Lin, L.; Sun, B.; Zhao, M. Comparison study on polysaccharide fractions from Laminaria japonica: Structural Characterization and bile acid binding capacity. J. Agricult. Food Chem. 2017, 65, 9790–9798. [Google Scholar] [CrossRef]
- Gelissen, I.C.; Eastwood, M.A. Taurocholic acid adsorption during non-starch polysaccharide fermentation: An in vitro study. Brit. J. Nutr. 1995, 74, 221. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Hu, D.; Cui, Z.; Li, H.; Man, C.; Jiang, Y. Lipid-lowering effects of Inonotus obliquus polysaccharide in vivo and in vitro. Foods 2021, 10, 3085. [Google Scholar] [CrossRef]
- Bai, J.; Li, J.; Pan, R.; Zhu, Y.; Xiao, X.; Li, Y.; Li, C. Polysaccharides from Volvariella volvacea inhibit fat accumulation in C. elegans dependent on the aak-2/nhr-49-mediated pathway. J. Food Biochem. 2021, 45, e13912. [Google Scholar] [CrossRef]
- Aguilera-Angel, E.-Y.; Espinal-Ruiz, M.; Narváez-Cuenca, C.-E. Pectic polysaccharides with different structural characteristics as inhibitors of pancreatic lipase. Food Hydrocoll. 2018, 83, 229–238. [Google Scholar] [CrossRef]
- Wu, D.-T.; Zhao, Y.-X.; Guo, H.; Gan, R.-Y.; Peng, L.-X.; Zhao, G.; Zou, L. Physicochemical and biological properties of polysaccharides from Dictyophora indusiata prepared by different extraction techniques. Polymers 2021, 13, 2357. [Google Scholar] [CrossRef]
- Hong, Y.; Yin, J.; Nie, S.; Xie, M. Applications of infrared spectroscopy in polysaccharide structural analysis: Progress, challenge and perspective. Food Chem. X 2021, 12, 100168. [Google Scholar] [CrossRef]
- Ho, T.C.; Kiddane, A.T.; Sivagnanam, S.P.; Park, J.-S.; Cho, Y.-J.; Getachew, A.T.; Nguyen, T.-T.T.; Kim, G.-D.; Chun, B.-S. Green extraction of polyphenolic-polysaccharide conjugates from Pseuderanthemum palatiferum (Nees) Radlk.: Chemical profile and anticoagulant activity. Int. J. Biol. Macromol. 2020, 157, 484–493. [Google Scholar] [CrossRef]
- Kacuráková, M. FT-IR study of plant cell wall model compounds: Pectic polysaccharides and hemicelluloses. Carbohydr. Polym. 2000, 43, 195–203. [Google Scholar] [CrossRef]
- Čopíková, J.; Černá, M.; Novotná, M.; Kaasová, J.; Synytsya, A. Application of FT-IR spectroscopy in detection of food hydrocolloids in confectionery jellies and food supplements. Czech J. Food Sci. 2001, 19, 51–56. [Google Scholar] [CrossRef]
- Synytsya, A. Fourier transform Raman and infrared spectroscopy of pectins. Carbohydr. Polym. 2003, 54, 97–106. [Google Scholar] [CrossRef]
- Ridley, B.L.; O’Neill, M.A.; Mohnen, D. Pectins: Structure, biosynthesis, and oligogalacturonide-related signaling. Phytochemistry 2001, 57, 929–967. [Google Scholar] [CrossRef]
- Vismeh, R.; Lu, F.; Chundawat, S.P.S.; Humpula, J.; Azarpira, A.; Balan, V.; Dale, B.E.; Ralph, J.; Jones, A.D. Profiling of diferulates (plant cell wall cross-linkers) using ultra high performance liquid chromatography-tandem mass spectrometry. Analyst 2013, 138, 1683. [Google Scholar] [CrossRef]
- Fry, S.C. Feruloylated pectins from the primary cell wall: Their structure and possible functions. Planta 1983, 157, 111–123. [Google Scholar] [CrossRef]
- Fry, S.; Willis, S.; Paterson, A. Intraprotoplasmic and wall-localised formation of arabinoxylan-bound diferulates and larger ferulate coupling-products in maize cell-suspension cultures. Planta 2000, 211, 679–692. [Google Scholar] [CrossRef]
- Rouau, X.; Cheynier, V.; Surget, A.; Gloux, D.; Barron, C.; Meudec, E.; Criton, M. A dehydrotrimer of ferulic acid from maize bran. Phytochemistry 2003, 63, 899–903. [Google Scholar] [CrossRef]
- Chen, M.; Gitz, D.C.; McClure, J.W. Soluble sinapoyl esters are converted to wall-bound esters in phenylalanine ammonia-lyase-inhibited radish seedlings. Phytochemistry 1998, 49, 333–340. [Google Scholar] [CrossRef]
- Pedersen, T.R.; Tobert, J.A. Simvastatin: A review. Exp. Opin. Pharmacother. 2004, 5, 2583–2596. [Google Scholar] [CrossRef]
- Franzoni, F.; Quiñones-Galvan, A.; Regoli, F.; Ferrannini, E.; Galetta, F. A comparative study of the in vitro antioxidant activity of statins. Int. J. Cardiol. 2003, 90, 317–321. [Google Scholar] [CrossRef]
- Kalita, P.; Ahmed, A.B.; Sen, S.; Chakraborty, R. A comprehensive review on polysaccharides with hypolipidemic activity: Occurrence, chemistry and molecular mechanism. Int. J. Biol. Macromol. 2022, 206, 681–698. [Google Scholar] [CrossRef]
- Sedigheh, A.; Jamal, S.M.; Mahbubeh, S.; Somayeh, K.; Mahmoud, R.-K.; Azadeh, A. Hypoglycaemic and hypolipidemic effects of pumpkin (Cucurbita pepo L.) on alloxan-induced diabetic rats. Afr. J. Pharm. Pharmacol. 2011, 5, 2620–2626. [Google Scholar] [CrossRef]
- Song, H.; Sun, Z. Hypolipidaemic and hypoglycaemic properties of pumpkin polysaccharides. 3Biotech 2017, 7, 159. [Google Scholar] [CrossRef] [PubMed]
- Ming, M.; Guanhua, L.; Zhanhai, Y.; Guang, C.; Xuan, Z. Effect of the Lycium barbarum polysaccharides administration on blood lipid metabolism and oxidative stress of mice fed high-fat diet in vivo. Food Chem. 2009, 113, 872–877. [Google Scholar] [CrossRef]
- Ji, X.; Liu, F.; Peng, Q.; Wang, M. Purification, structural characterization, and hypolipidemic effects of a neutral polysaccharide from Ziziphus jujuba cv. Muzao. Food Chem. 2018, 245, 1124–1130. [Google Scholar] [CrossRef]
- Wang, D.; Li, C.; Fan, W.; Yi, T.; Wei, A.; Ma, Y. Hypoglycemic and hypolipidemic effects of a polysaccharide from Fructus Corni in streptozotocin-induced diabetic rats. Int. J. Biol. Macromol. 2019, 133, 420–427. [Google Scholar] [CrossRef]
- Zhao, X.H.; Qian, L.; Yina, D.; Zhou, Y. Hypolipidemic effect of the polysaccharides extracted from pumpkin by cellulase-assisted method on mice. Int. J. Biol. Macromol. 2014, 64, 137–138. [Google Scholar] [CrossRef]
- Kolsi, R.B.A.; Jardak, N.; Hadjkacem, F.; Chaaben, R.; Jribi, I.; Feki, A.E.; Rebai, T.; Jamoussi, K.; Fki, L.; Belghith, H.; et al. Anti-obesity effect and protection of liver-kidney functions by Codium fragile sulphated polysaccharide on high fat diet induced obese rats. Int. J. Biol. Macromol. 2017, 102, 119–129. [Google Scholar] [CrossRef]
- Teng, Z.; Qian, L.; Zhou, Y. Hypolipidemic activity of the polysaccharides from Enteromorpha prolifera. Int. J. Biol. Macromol. 2013, 62, 254–256. [Google Scholar] [CrossRef]
- Rjeibi, I.; Feriani, A.; Hentati, F.; Hfaiedh, N.; Michaud, P.; Pierre, G. Structural characterization of water-soluble polysaccharides from Nitraria retusa fruits and their antioxidant and hypolipidemic activities. Int. J. Biol. Macromol. 2019, 129, 422–432. [Google Scholar] [CrossRef] [PubMed]
- Nie, Q.; Hu, J.; Gao, H.; Fan, L.; Chen, H.; Nie, S. Polysaccharide from Plantago asiatica L. attenuates hyperglycemia, hyperlipidemia and affects colon microbiota in type 2 diabetic rats. Food Hydrocoll. 2019, 86, 34–42. [Google Scholar] [CrossRef]
- Kolodziejczyk-Czepas, J.; Bijak, M.; Saluk, J.; Ponczek, M.B.; Zbikowska, H.M.; Nowak, P.; Tsirigotis-Maniecka, M.; Pawlaczyk, I. Radical scavenging and antioxidant effects of Matricaria chamomilla polyphenolic–polysaccharide conjugates. Int. J. Biol. Macromol. 2015, 72, 1152–1158. [Google Scholar] [CrossRef] [PubMed]
- Campos, F.; Peixoto, A.F.; Fernandes, P.A.R.; Coimbra, M.A.; Mateus, N.; de Freitas, V.; Fernandes, I.; Fernandes, A. The antidiabetic effect of grape pomace polysaccharide-polyphenol complexes. Nutrients 2021, 13, 4495. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Liang, R.; Dai, T.; Chen, J.; Shuai, X.; Liu, C. Pectin-based adsorbents for heavy metal ions: A review. Trends Food Sci. Technol. 2019, 91, 319–329. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polysaccharide Fraction | Yield | Total Carbohydrate, % a | Uronic Acids, % a | Starch, % a | Proteins, % a | AGP, % a | Phenolics, % a | Ash, % a |
|---|---|---|---|---|---|---|---|---|
| VVPS | 3.7 b | 92.89 ± 2.78 i | 37.62 ± 0.75 vii | 2.04 ± 0.04 ix | 1.67 ± 0.05 xiii | 0.29 ± 0.01 xv | 2.96 ± 0.10 xix | 1.56 ± 0.03 xxii |
| DEAE-H2O | 7.0 c | 96.11 ± 2.89 ii | - | 25.36 ± 0.52 xi | 3.85 ± 0.12 xiv | 3.44 ± 0.21 xvii | - | trace |
| DEAE-0.1% NH4HCO3 | 0.8 c | 97.14 ± 2.99 ii | - | 9.27 ± 0.18 x | 1.86 ± 0.05 xiii | 0.86 ± 0.03 xvi | - | trace |
| DEAE-0.3% NH4HCO3 | 1.8 c | 98.06 ± 3.04 iii | 3.22 ± 0.05 iv | 0.53 ± 0.01 viii | 0.53 ± 0.01 xii | - | - | trace |
| DEAE-0.5% NH4HCO3 | 2.0 c | 98.15 ± 3.01 iii | 29.53 ± 0.57 v | - | - | - | - | trace |
| DEAE-1% NH4HCO3 | 0.4 c | 98.24 ± 3.11 iii | 34.12 ± 0.48 vi | - | - | - | 0.34 ± 0.01 xviii | 0.52 ± 0.01 xxi |
| DEAE-1% NaOH | 81.0 c | 92.37 ± 2.76 i | 39.67 ± 0.74 vii | - | 0.61 ± 0.02 xii | - | 3.54 ± 0.12 xx | 1.90 ± 0.05 xxiii |
| Polysaccharide Fraction | C | H | O | N | O/C | H/C |
|---|---|---|---|---|---|---|
| VVPS | 39.08 | 6.14 | 54.52 | 0.26 | 1.395 | 0.157 |
| DEAE-H2O | 39.92 | 6.65 | 52.81 | 0.62 | 1.323 | 0.167 |
| DEAE-0.1% NH4HCO3 | 39.96 | 6.67 | 53.08 | 0.29 | 1.328 | 0.167 |
| DEAE-0.3% NH4HCO3 | 39.87 | 6.62 | 53.43 | 0.08 | 1.340 | 0.166 |
| DEAE-0.5% NH4HCO3 | 39.22 | 6.21 | 54.57 | - | 1.391 | 0.158 |
| DEAE-1% NH4HCO3 | 39.92 | 6.40 | 53.68 | - | 1.345 | 0.160 |
| DEAE-1% NaOH | 38.94 | 6.10 | 54.29 | 0.10 | 1.394 | 0.157 |
| Hexose (C6H12O6), pentose (C5H10O5) | 40.00 | 6.67 | 53.33 | - | 1.333 | 0.167 |
| Desoxyhexose (C6H12O5) | 43.90 | 7.31 | 48.79 | - | 1.111 | 0.166 |
| Hexuronic acid (C6H10O7) | 37.11 | 5.15 | 57.74 | - | 1.556 | 0.138 |
| Polysaccharide Fraction, Vaccinium Species | Ara | Gal | Glc | Fuc | Man | Rib | Rha | Xyl | GalA | GlcA |
|---|---|---|---|---|---|---|---|---|---|---|
| VVPS | 22.4 | 12.6 | 23.2 | 0.4 | 0.9 | traces | 4.6 | traces | 35.4 | 0.6 |
| V. arctostaphylos PS [52] | 39.2 | 21.0 | 6.3 | - | - | - | 1.5 | 13.7 | 18.3 * | |
| V. ashei PS [50] | 29.7 | 10.9 | 15.5 | - | 1.2 | - | 1.6 | 3.6 | 37.3 | - |
| V. myrtillus PS [53] | 4.0 | 4.0 | 41.0 | - | 3.0 | - | 1.0 | 33.0 | 14.0 | 1.0 |
| V. uliginosum PS [54] | 36.4 | 24.5 | 25.7 | - | 3.1 | - | - | - | 10.3 | - |
| V. vitis-idaea ssp. minus PS [29] | 26.8 | 8.6 | 14.6 | 0.3 | 0.8 | - | 1.3 | 8.5 | 39.1 * | |
| DEAE-H2O | 29.3 | 22.5 | 45.5 | - | 2.6 | - | - | - | - | - |
| DEAE-0.1% NH4HCO3 | 25.5 | 27.8 | 41.4 | 0.5 | 4.8 | - | - | - | - | - |
| DEAE-0.3% NH4HCO3 | 31.2 | 33.0 | 28.3 | - | 4.7 | - | - | - | 2.8 | - |
| DEAE-0.5% NH4HCO3 | 22.0 | 29.4 | 11.5 | - | 2.9 | 0.1 | 4.4 | 0.3 | 27.7 | 1.7 |
| DEAE-1% NH4HCO3 | 24.0 | 19.2 | 5.0 | - | 3.6 | - | 17.6 | - | 30.6 | - |
| DEAE-1% NaOH | 26.2 | 13.9 | 0.9 | - | 0.7 | - | 3.6 | - | 54.8 | - |
| Polysaccharide Fraction | DPPH a | ABTS+ a | O2•− a | OH• a | Cl b | NO a | H2O2 c | FeCA d |
|---|---|---|---|---|---|---|---|---|
| VVPS | 35.69 ± 0.73 v | 22.59 ± 0.45 x | 144.17 ± 4.32 xv | 32.60 ± 0.96 xix | 27.56 ± 0.55 xx | 92.75 ± 2.84 xxvii | 0.36 ± 0.01 xxxii | 4.71 ± 0.14 xxxviii |
| DEAE-H2O | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. |
| DEAE-0.1% NH4HCO3 | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. |
| DEAE-0.3% NH4HCO3 | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. |
| DEAE-0.5% NH4HCO3 | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. | 0.25 ± 0.00 xxxi | 0.31 ± 0.01 xxxvii |
| DEAE-1% NH4HCO3 | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. | 0.18 ± 0.00 xxxi | 0.24 ± 0.00 xxxvi |
| DEAE-1% NaOH | 24.18 ± 0.48 iv | 15.25 ± 0.31 ix | 108.26 ± 2.07 xiii | 22.86 ± 0.67 xviii | 41.67 ± 0.83 xxi | 72.11 ± 2.16 xxvi | 0.27 ± 0.01 | 6.83 ± 0.20 xxxxi |
| DEAE-1% NaOH-f1 | 12.73 ± 0.25 iii | 7.62 ± 0.15 viii | 84.75 ± 1.76 xii | 14.06 ± 0.42 xvii | 126.79 ± 2.53 xxii | 53.86 ± 1.61 xxv | 0.10 ± 0.00 xxx | 8.26 ± 0.34 xxxxii |
| DEAE-1% NaOH-f2 | 10.82 ± 0.21 ii | 6.83 ± 0.12 vii | 70.29 ± 1.70 xi | 11.73 ± 0.39 xvi | 157.11 ± 3.14 xxiii | 41.09 ± 1.20 xxiv | 0.08 ± 0.00 xxx | 8.72 ± 0.35 xxxxii |
| DEAE-1% NaOH-f3 | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. | 6.14 ± 0.25 xxxx |
| DEAE-1% NaOH-f4 | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. | 6.09 ± 0.24 xxxx |
| Trolox e | 8.94 ± 0.18 i | 3.25 ± 0.06 vi | 122.36 ± 2.44 xiv | 15.23 ± 0.36 xvii | 1000 | 125.11 ± 3.75 xxviii | 0.59 ± 0.02 xxxiii | 0.15 ± 0.00 xxxv |
| Pectin from citrus peel e | i.a. | i.a. | i.a. | i.a. | i.a. | 265.82 ± 10.63 xxix | 2.77 ± 0.11 xxxiv | 5.26 ± 0.15 xxxix |
| Starch e | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. |
| Arabinogalactan e | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. | i.a. | 0.22 ± 0.01 xxxvi |
| Polysaccharide Fraction | Bile Acids Binding, μmole/100 g | Fat Binding, g/100 g | Cholesterol Binding, mg/g | Pancreatic Lipase Inhibition, IC50, mg/mL |
|---|---|---|---|---|
| VVPS | 5.73 ± 0.22 viii | 200.02 ± 7.24 xvi | 57.02 ± 2.56 xxiii | 6.24 ± 0.18 xxx |
| DEAE-H2O | 0.10 ± 0.00 ii | 103.75 ± 3.70 xii | 15.37 ± 0.63 xx | i.a. |
| DEAE-0.1% NH4HCO3 | 0.14 ± 0.00 ii | 127.80 ± 4.49 xiii | 18.62 ± 0.80 xxi | i.a. |
| DEAE-0.3% NH4HCO3 | 0.27 ± 0.01 iii | 131.29 ± 4.63 xiii | 19.83 ± 0.93 xxi | i.a. |
| DEAE-0.5% NH4HCO3 | 0.92 ± 0.04 v | 139.16 ± 4.90 xiii | 37.10 ± 1.69 xxii | i.a. |
| DEAE-1% NH4HCO3 | 3.62 ± 0.16 vii | 231.02 ± 8.14 xvii | 59.22 ± 2.50 xxiii | i.a. |
| DEAE-1% NaOH | 6.04 ± 0.29 viii | 252.37 ± 8.97 xvii | 68.37 ± 3.02 xxiv | 5.33 ± 0.15 xix |
| DEAE-1% NaOH-f1 | 7.83 ± 0.39 ix | 183.70 ± 6.40 xv | 72.11 ± 3.24 xxv | 4.27 ± 0.12 xxviii |
| DEAE-1% NaOH-f2 | 8.26 ± 0.44 ix | 173.11 ± 6.04 xv | 73.92 ± 3.36 xxv | 3.86 ± 0.10 xxvii |
| DEAE-1% NaOH-f3 | 1.85 ± 0.09 vi | 308.75 ± 10.83 xviii | 59.27 ± 2.65 xxiii | i.a. |
| DEAE-1% NaOH-f4 | 1.90 ± 0.10 vi | 315.61 ± 10.88 xviii | 60.08 ± 2.72 xxiii | i.a. |
| Cholestyramine * | 10.29 ± 0.40 x | i.a. | 93.11 ± 4.15 xxvi | 14.02 ± 0.42 xxxi |
| Microcrystalline cellulose * | 0.07 ± 0.00 ii | 92.63 ± 3.12 xi | 10.33 ± 0.45 xix | i.a. |
| Pectin from citrus peel * | 0.78 ± 0.03 iv | 186.85 ± 6.51 xv | 57.82 ± 2.69 xxiii | i.a. |
| Starch * | 0.02 ± 0.00 i | 97.67 ± 3.43 xi | 9.63 ± 0.44 xix | i.a. |
| Arabinogalactan * | 0.12 ± 0.00 ii | 156.14 ± 5.46 xiv | 21.16 ± 0.90 xxi | i.a. |
| Parameter | DEAE-1% NaOH-f1 | DEAE-1% NaOH-f1-d | DEAE-1% NaOH-f2 | DEAE-1% NaOH-f2-d | DEAE-1% NaOH-f3 | DEAE-1% NaOH-f4 |
|---|---|---|---|---|---|---|
| Yield | 9.5 a | 26.1 b | 15.6 a | 29.3 c | 58.4 a | 4.2 a |
| Mw, kDa d | 157.6 (±1.4%) | 35.5 (±1.0%) | 108.2 (±1.9%) | 25.4 (±1.4%) | 258.3 (±2.2%) | 318.4 (±2.7%) |
| Mw/Mn d | 1.56 (±2.9%) | 1.64 (±3.7%) | 1.48 (±2.1%) | 1.42 (±2.6%) | 1.71 (±3.7%) | 1.62 (±2.4%) |
| Monosaccharide composition, mol% | ||||||
| Ara | 65.1 | - | 58.6 | - | 15.5 | 16.8 |
| Gal | 33.1 | 99.9 | 38.7 | 99.9 | 7.1 | 8.9 |
| Glc | 0.6 | - | 1.7 | - | 0.2 | 0.1 |
| Man | 1.2 | - | 1.0 | - | 2.7 | 2.1 |
| Rha | - | - | - | - | 6.7 | 4.1 |
| GalA | - | - | - | - | 67.8 | 68.0 |
| Linkage analysis, molar ratio | ||||||
| Terminal Ara | 12.6 | - | 12.4 | - | 3.7 | 3.2 |
| 1,5-Ara | 38.4 | - | 31.6 | - | 10.3 | 10.9 |
| 1,3,5-Ara | 14.1 | - | 15.0 | - | 1.2 | 2.8 |
| Terminal Gal | 7.1 | 1.0 | 10.2 | 1.4 | 8.2 | 7.9 |
| 1,3-Gal | 10.5 | 98.9 | 9.1 | 98.5 | 4.2 | 5.7 |
| 1,3,6-Gal | 15.7 | - | 19.4 | - | 2.6 | 3.0 |
| 1,4-Gal | - | - | - | - | 60.9 | 61.3 |
| 1,2-Rha | - | - | - | - | 4.6 | 3.6 |
| 1,2,4-Rha | - | - | - | - | 2.0 | 0.6 |
| Terminal Man | 1.0 | - | 0.4 | - | 2.0 | 0.9 |
| Terminal Glc | 0.6 | - | 1.9 | - | 0.3 | 0.1 |
| Phenols, % e | 10.61 (±0.32) | - | 14.52 (±0.44) | - | <0.1 | <0.1 |
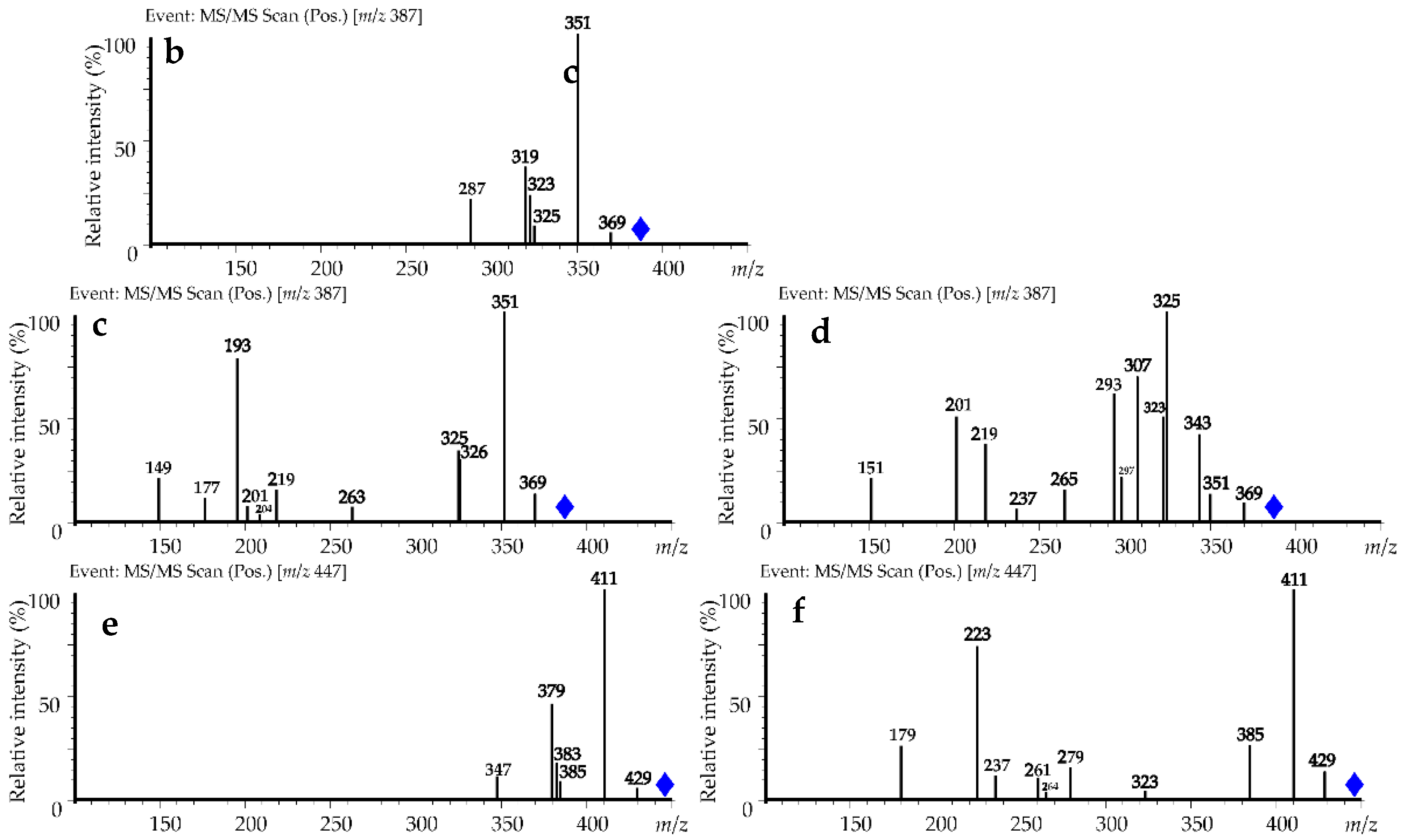
| Compound (No. Figure 5) | ESI-MS, [M + H]+, m/z | ESI-MS/MS, m/z | Content after Alkaline Destruction, % a | |
|---|---|---|---|---|
| DEAE-1% NaOH-f1 | DEAE-1% NaOH-f2 | |||
| Ferulic acid (1) | 195 | 181 | 38.4 ± 1.1 | 29.5 ± 0.8 |
| Sinapic acid (2) | 225 | 211, 197 | 9.6 ± 0.2 | 15.8 ± 0.4 |
| Diferulic acid (3) | 387 | 369, 351, 325, 323, 319, 287 | 10.5 ± 0.1 | 12.7 ± 0.2 |
| Diferulic acid (4) | 387 | 369, 351, 326, 325, 263, 219, 204, 201, 193, 177, 149 | 28.6 ± 0.4 | 35.9 ± 0.7 |
| Diferulic acid (5) | 387 | 369, 351, 343, 325, 323, 307, 297, 293, 265, 237, 219, 201, 151 | 8.2 ± 0.2 | 3.3 ± 0.1 |
| Disinapic acid (6) | 447 | 429, 385, 383, 411, 379, 347 | 1.4 ± 0.0 | 0.8 ± 0.0 |
| Disinapic acid (7) | 447 | 429, 411, 401, 323, 279, 264, 261, 237, 223, 205, 179, 177 | 3.3 ± 0.1 | 2.0 ± 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olennikov, D.N.; Chemposov, V.V.; Chirikova, N.K. Polymeric Compounds of Lingonberry Waste: Characterization of Antioxidant and Hypolipidemic Polysaccharides and Polyphenol-Polysaccharide Conjugates from Vaccinium vitis-idaea Press Cake. Foods 2022, 11, 2801. https://doi.org/10.3390/foods11182801
Olennikov DN, Chemposov VV, Chirikova NK. Polymeric Compounds of Lingonberry Waste: Characterization of Antioxidant and Hypolipidemic Polysaccharides and Polyphenol-Polysaccharide Conjugates from Vaccinium vitis-idaea Press Cake. Foods. 2022; 11(18):2801. https://doi.org/10.3390/foods11182801
Chicago/Turabian StyleOlennikov, Daniil N., Vladimir V. Chemposov, and Nadezhda K. Chirikova. 2022. "Polymeric Compounds of Lingonberry Waste: Characterization of Antioxidant and Hypolipidemic Polysaccharides and Polyphenol-Polysaccharide Conjugates from Vaccinium vitis-idaea Press Cake" Foods 11, no. 18: 2801. https://doi.org/10.3390/foods11182801
APA StyleOlennikov, D. N., Chemposov, V. V., & Chirikova, N. K. (2022). Polymeric Compounds of Lingonberry Waste: Characterization of Antioxidant and Hypolipidemic Polysaccharides and Polyphenol-Polysaccharide Conjugates from Vaccinium vitis-idaea Press Cake. Foods, 11(18), 2801. https://doi.org/10.3390/foods11182801