


Study of Lactic Acid Bacteria Biodiversity in Fermented Cobrançosa Table Olives to Determine Their Probiotic Potential

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Bacterial Strains and Growth Conditions
2.3. Identification and Typing
2.4. Assessment of Food Safety for Human Use
2.5. Mucin Degradation Test
2.6. Hemolytic Activity Test
2.7. DNase Activity Test
2.8. Assessment of Gastrointestinal Survival
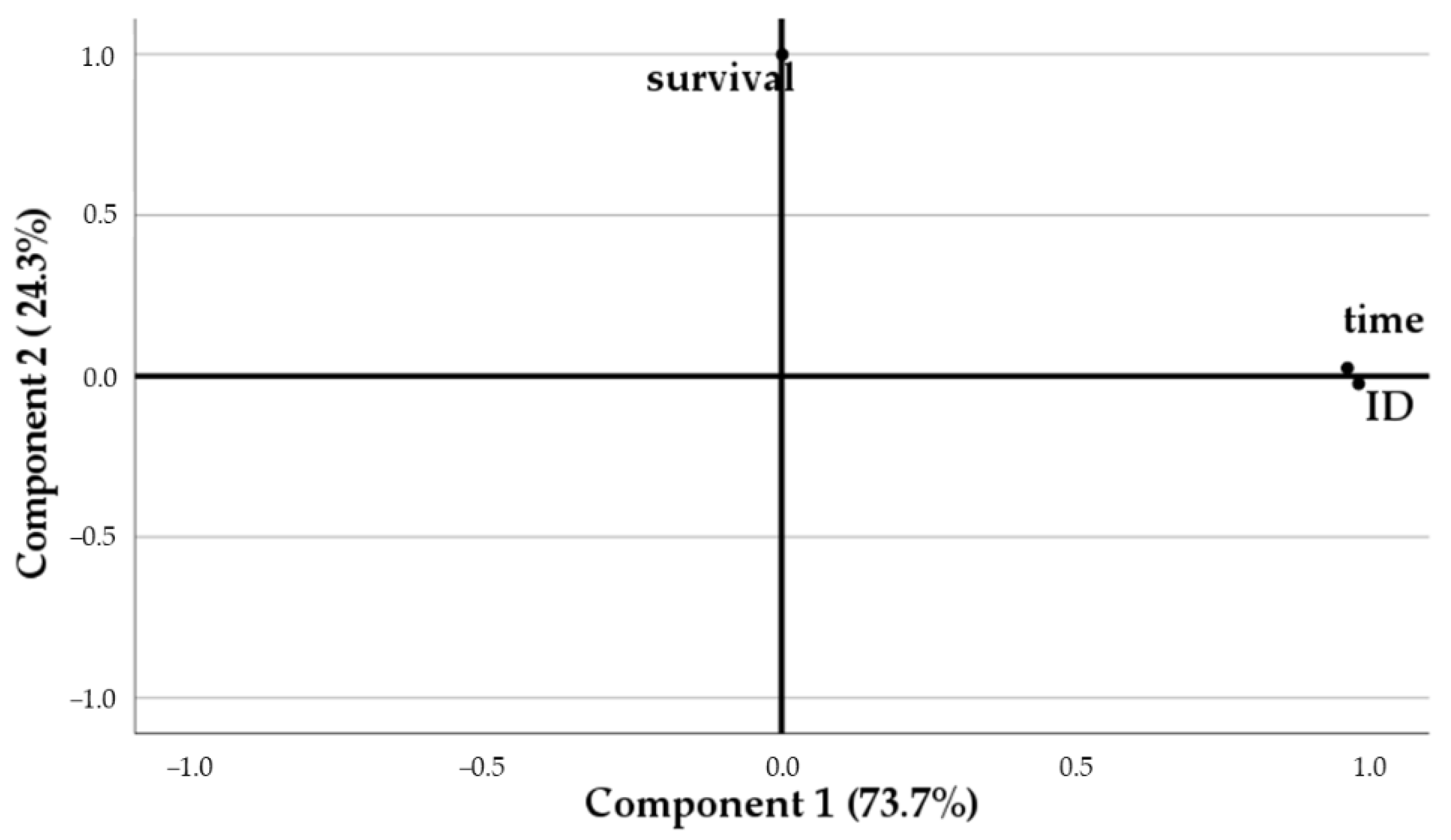
2.9. Statistical Analysis
3. Results and Discussion
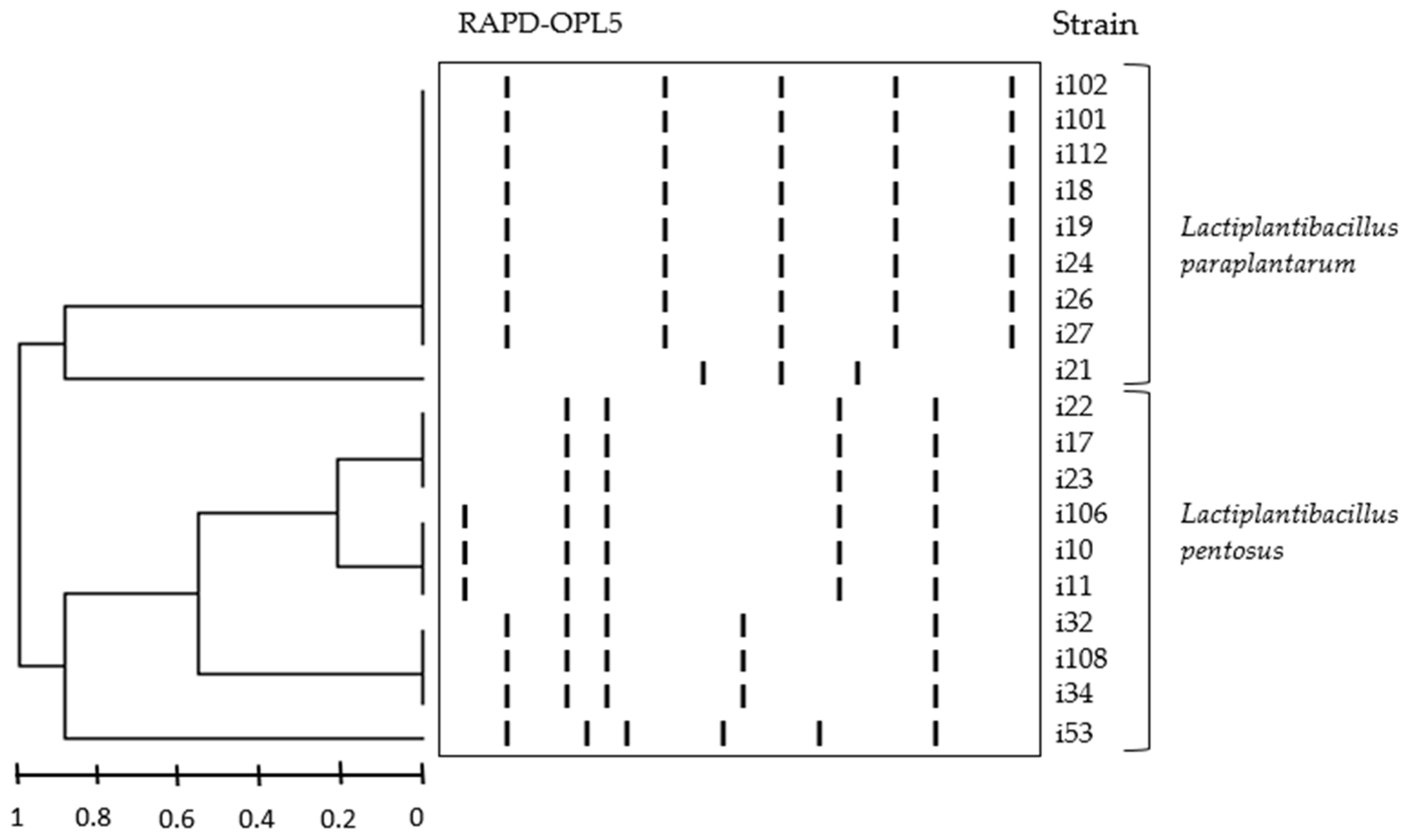
3.1. DNA-Fingerprinting and Identification
3.2. Assessment of Food Safety for Human Use
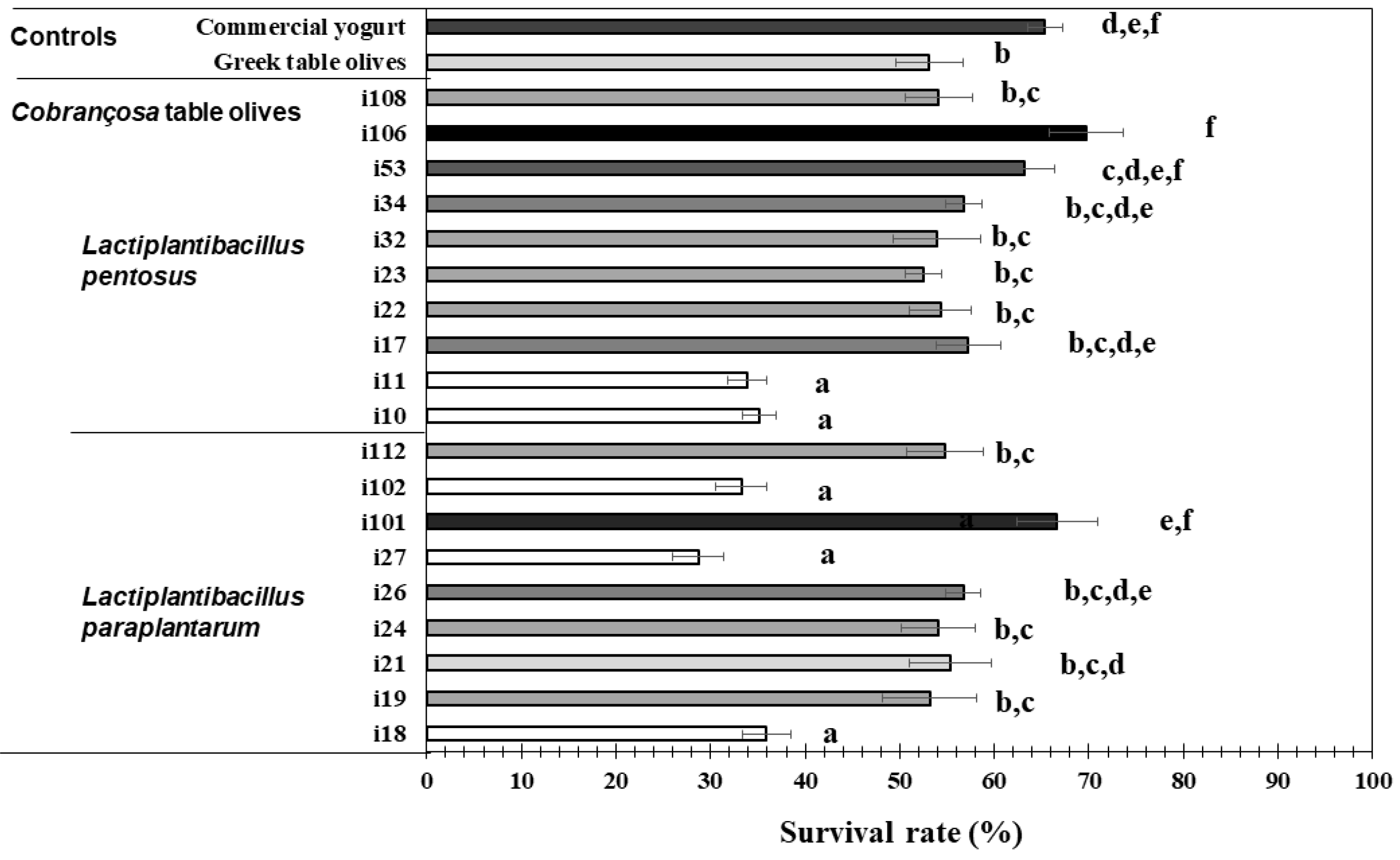
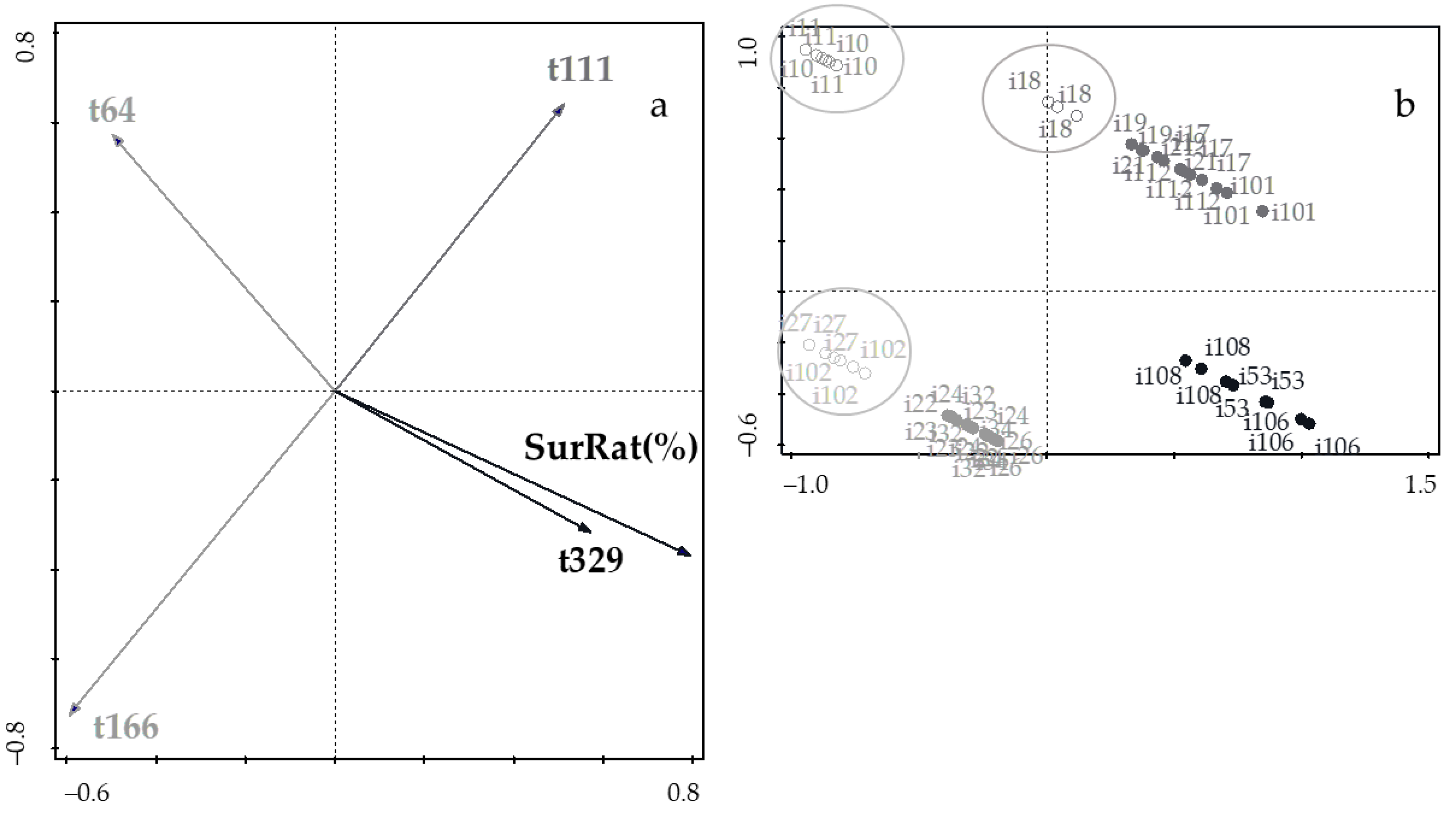
3.3. Assesment of Gastrointestinal Survival
3.4. Relevance and Pratical Implications
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bigliardi, B.; Filippelli, S. A review of the literature on innovation in the agrofood industry: Sustainability, smartness and health. Eur. J. Innov. Manag. 2022, 25, 589–611. [Google Scholar] [CrossRef]
- Koirala, S.; Anal, A.K. Probiotics-based foods and beverages as future foods and their overall safety and regulatory claims. Future Foods 2021, 3, 100013. [Google Scholar] [CrossRef]
- Albuquerque, T.G.; Costa, H.S.; Oliveira, M.B.P.P. An overview of Portuguese olive oils and table olives with Protected Designation of Origin. Eur. J. Lipid Sci. Technol. 2019, 121, 1800129. [Google Scholar] [CrossRef]
- De Bellis, P.; Sisto, A.; Lavermicocca, P. Probiotic bacteria and plant-based matrices: An association with improved health-promoting features. J. Funct. Foods 2021, 87, 104821. [Google Scholar] [CrossRef]
- Bulut Albayrak, Ç.; Duran, M. Worldwide research tendencies on probiotics in food science: 1993 to 2021. Br. Food J. 2022, 124, 679–700. [Google Scholar] [CrossRef]
- Anagnostopoulos, D.A.; Tsaltas, D. Current status, recent advances, and main challenges on table olive fermentation: The present meets the future. Front. Microbiol. 2022, 12, 797295. [Google Scholar] [CrossRef]
- Perpetuini, G.; Prete, R.; Garcia-Gonzalez, N.; Khairul Alam, M.; Corsetti, A. Table olives more than a fermented food. Foods 2020, 9, 178. [Google Scholar] [CrossRef] [Green Version]
- Pires-Cabral, P.; Nunes, P.; Barros, T.; Quintas, C. Physicochemical, nutritional and microbiological characteristics of traditional table olives from Southern Portugal. Emir. J. Food Agric. 2018, 30, 611–620. [Google Scholar] [CrossRef]
- Peres, C.M.; Alves, M.; Hernandez-Mendoza, A.; Moreira, L.; Silva, S.; Bronze, M.R.; Vilas-Boas, L.; Peres, C.; Malcata, F.X. Novel isolates of lactobacilli from fermented Portuguese olive as potential probiotics. LWT Food Sci. Technol. 2014, 59, 234–246. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, T.; Ramalhosa, E.; Nunes, L.; Pereira, J.A.; Colla, E.; Pereira, E.L. Probiotic potential of indigenous yeasts isolated during the fermentation of table olives from Northeast of Portugal. Innov. Food Sci. Emerg. Technol. 2017, 44, 167–172. [Google Scholar] [CrossRef] [Green Version]
- Pereira, E.L.; Ramalhosa, E.; Borges, A.; Pereira, J.A.; Baptista, P. YEAST dynamics during the natural fermentation process of table olives (Negrinha de Freixo cv.). Food Microbiol. 2015, 46, 582–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, J.; Yang, K.; Liu, M.; Zhang, J.; Wei, X.; Fan, M. Screening for potential probiotic from spontaneously fermented non-dairy foods based on in vitro probiotic and safety properties. Ann. Microbiol. 2018, 68, 803–813. [Google Scholar] [CrossRef]
- Abouloifa, H.; Rokni, Y.; Bellaouchi, R.; Ghabbour, N.; Karboune, S.; Brasca, M.; Ben Salah, R.; Chihib, N.; Saalaoui, E.; Asehraou, A. Characterization of Probiotic Properties of Antifungal Lactobacillus Strains Isolated from Traditional Fermenting Green Olives. Probiotics Antimicrob. Proteins 2020, 12, 683–696. [Google Scholar] [CrossRef] [PubMed]
- de Melo Pereira, G.V.; de Oliveira Coelho, B.; Magalhaes Junior, A.I.; Thomaz-Soccol, V.; Soccol, C.R. How to select a probiotic? A review and update of methods and criteria. Biotechnol. Adv. 2018, 36, 2060–2076. [Google Scholar] [CrossRef] [PubMed]
- Bonatsou, S.; Tassou, C.C.; Panagou, E.Z.; Nychas, G.E. Table olive fermentation using starter cultures with multifunctional potential. Microorganisms 2017, 5, 30. [Google Scholar] [CrossRef] [Green Version]
- Maldonado-Barragan, A.; Caballero-Guerrero, B.; Lucena-Padros, H.; Ruiz-Barba, J.L. Induction of bacteriocin production by coculture is widespread among plantaricin-producing Lactobacillus plantarum strains with different regulatory operons. Food Microbiol. 2013, 33, 40–47. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Benitez-Cabello, A.; Calero-Delgado, B.; Rodriguez-Gomez, F.; Garrido-Fernandez, A.; Jimenez-Diaz, R.; Arroyo-Lopez, F.N. Biodiversity and multifunctional features of lactic acid bacteria isolated from table olive biofilms. Front. Microbiol. 2019, 10, 836. [Google Scholar] [CrossRef] [Green Version]
- Anagnostopoulos, D.; Bozoudi, D.; Tsaltas, D. Enterococci isolated from Cypriot green table olives as a new source of technological and probiotic properties. Fermentation 2018, 4, 48. [Google Scholar] [CrossRef] [Green Version]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carriere, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Mulet-Cabero, A.I.; Egger, L.; Portmann, R.; Menard, O.; Marze, S.; Minekus, M.; Le Feunteun, S.; Sarkar, A.; Grundy, M.M.; Carriere, F.; et al. A standardised semi-dynamic in vitro digestion method suitable for food—An international consensus. Food Funct. 2020, 11, 1702–1720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bautista-Gallego, J.; Arroyo-López, F.N.; Rantsiou, K.; Jiménez-Díaz, R.; Garrido-Fernández, A.; Cocolin, L. Screening of lactic acid bacteria isolated from fermented table olives with probiotic potential. Food Res. Int. 2013, 50, 135–142. [Google Scholar] [CrossRef]
- Piepho, H.P. Letters in mean comparisons: What they do and don’t mean. J. Agron. 2018, 110, 431–434. [Google Scholar] [CrossRef] [Green Version]
- Portilha-Cunha, M.F.; Macedo, A.C.; Malcata, F.X. A review on adventitious lactic acid bacteria from table olives. Foods 2020, 9, 948. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.B.; Martinez, R.C.R.; Pereira, E.P.R.; Balthazar, C.F.; Cruz, A.G.; Ranadheera, C.S.; Sant’Ana, A.S. The resistance of Bacillus, Bifidobacterium, and Lactobacillus strains with claimed probiotic properties in different food matrices exposed to simulated gastrointestinal tract conditions. Food Res. Int. 2019, 125, 108542. [Google Scholar] [CrossRef]
- Botta, C.; Langerholc, T.; Cencic, A.; Cocolin, L. In vitro selection and characterization of new probiotic candidates from table olive microbiota. PLoS ONE 2014, 9, e94457. [Google Scholar] [CrossRef]
- Montoro, B.P.; Benomar, N.; Lavilla Lerma, L.; Castillo Gutierrez, S.; Galvez, A.; Abriouel, H. Fermented Alorena table olives as a source of potential probiotic Lactobacillus pentosus strains. Front. Microbiol. 2016, 7, 1583. [Google Scholar] [CrossRef] [Green Version]
- Arroyo-Lopez, F.N.; Blanquet-Diot, S.; Denis, S.; Thevenot, J.; Chalancon, S.; Alric, M.; Rodriguez-Gomez, F.; Romero-Gil, V.; Jimenez-Diaz, R.; Garrido-Fernandez, A. Survival of pathogenic and lactobacilli species of fermented olives during simulated human digestion. Front. Microbiol. 2014, 5, 540. [Google Scholar] [CrossRef]
- Guantario, B.; Zinno, P.; Schifano, E.; Roselli, M.; Perozzi, G.; Palleschi, C.; Uccelletti, D.; Devirgiliis, C. In vitro and in vivo selection of potentially probiotic Lactobacilli from Nocellara del Belice table olives. Front. Microbiol. 2018, 9, e595. [Google Scholar] [CrossRef] [Green Version]
- Tulini, F.L.; Winkelstroter, L.K.; De Martinis, E.C. Identification and evaluation of the probiotic potential of Lactobacillus paraplantarum FT259, a bacteriocinogenic strain isolated from Brazilian semi-hard artisanal cheese. Annaerobe 2013, 22, 57–63. [Google Scholar] [CrossRef]
- Kalhoro, M.S.; Visessanguan, W.; Nguyen, L.T.; Anal, A.K. Probiotic potential of Lactobacillus paraplantarum BT-11 isolated from raw buffalo (Bubalus bubalis) milk and characterization of bacteriocin-like inhibitory substance produced. J. Food Process. Preserv. 2019, 43, e14015. [Google Scholar] [CrossRef]
- IOOC. IOC News: Table Olives—Provisional 2019/20 Campaign and Estimate for 2020/21. Available online: https://www.internationaloliveoil.org/ (accessed on 25 May 2022).
- IOOC. IOC Figures: Production of Table Olives. Available online: https://www.internationaloliveoil.org/ (accessed on 25 May 2022).
- Berkes, E.; Liao, Y.H.; Neef, D.; Grandalski, M.; Monsul, N. Potentiated in vitro probiotic activities of Lactobacillus fermentum LfQi6 biofilm biomass versus planktonic culture. Probiotics Antimicrob. Proteins 2020, 12, 1097–1114. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oral | Gastric | Intestinal | |||
|---|---|---|---|---|---|
| Duration (min) | |||||
| 2 * | 120 | 120 | |||
| Fixed Volumes (mL) | |||||
| Salivary fluid (SSF) (5/4) | 3.2 | Liquid food | 8.0 | Liquid food | 16.0 |
| CaCl2 0.3 M | 0.020 | Gastric juice (SGF) (5/4) | 6.4 | Duodenal juice (SIF) (5/4) | 6.8 |
| Salivary amylase solution | 0.4 | Pepsin solution | 0.4 | Pancreatin | 4.0 |
| CaCl2 0.3 M | 0.004 | Bile | 2.0 | ||
| CaCl2 0.3 M | 0.032 | ||||
| Variable volumes (mL) | |||||
| HCl 1 M to pH 3.0 | 0.740 | Acid/base 1 M to pH 7.0 | 0.350 | ||
| Water | 0.380 | Water | 0.456 | Water | 2.818 |
| Total | 8.0 | Total | 16.0 | Total | 32.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coimbra-Gomes, J.; Reis, P.J.M.; Tavares, T.G.; Malcata, F.X.; Macedo, A.C. Study of Lactic Acid Bacteria Biodiversity in Fermented Cobrançosa Table Olives to Determine Their Probiotic Potential. Foods 2022, 11, 3050. https://doi.org/10.3390/foods11193050
Coimbra-Gomes J, Reis PJM, Tavares TG, Malcata FX, Macedo AC. Study of Lactic Acid Bacteria Biodiversity in Fermented Cobrançosa Table Olives to Determine Their Probiotic Potential. Foods. 2022; 11(19):3050. https://doi.org/10.3390/foods11193050
Chicago/Turabian StyleCoimbra-Gomes, Joana, Patrícia J. M. Reis, Tânia G. Tavares, Francisco Xavier Malcata, and Angela C. Macedo. 2022. "Study of Lactic Acid Bacteria Biodiversity in Fermented Cobrançosa Table Olives to Determine Their Probiotic Potential" Foods 11, no. 19: 3050. https://doi.org/10.3390/foods11193050
APA StyleCoimbra-Gomes, J., Reis, P. J. M., Tavares, T. G., Malcata, F. X., & Macedo, A. C. (2022). Study of Lactic Acid Bacteria Biodiversity in Fermented Cobrançosa Table Olives to Determine Their Probiotic Potential. Foods, 11(19), 3050. https://doi.org/10.3390/foods11193050