
Assessing Edible Filamentous Fungal Carriers as Cell Supports for Growth of Yeast and Cultivated Meat


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Cells and Media Components
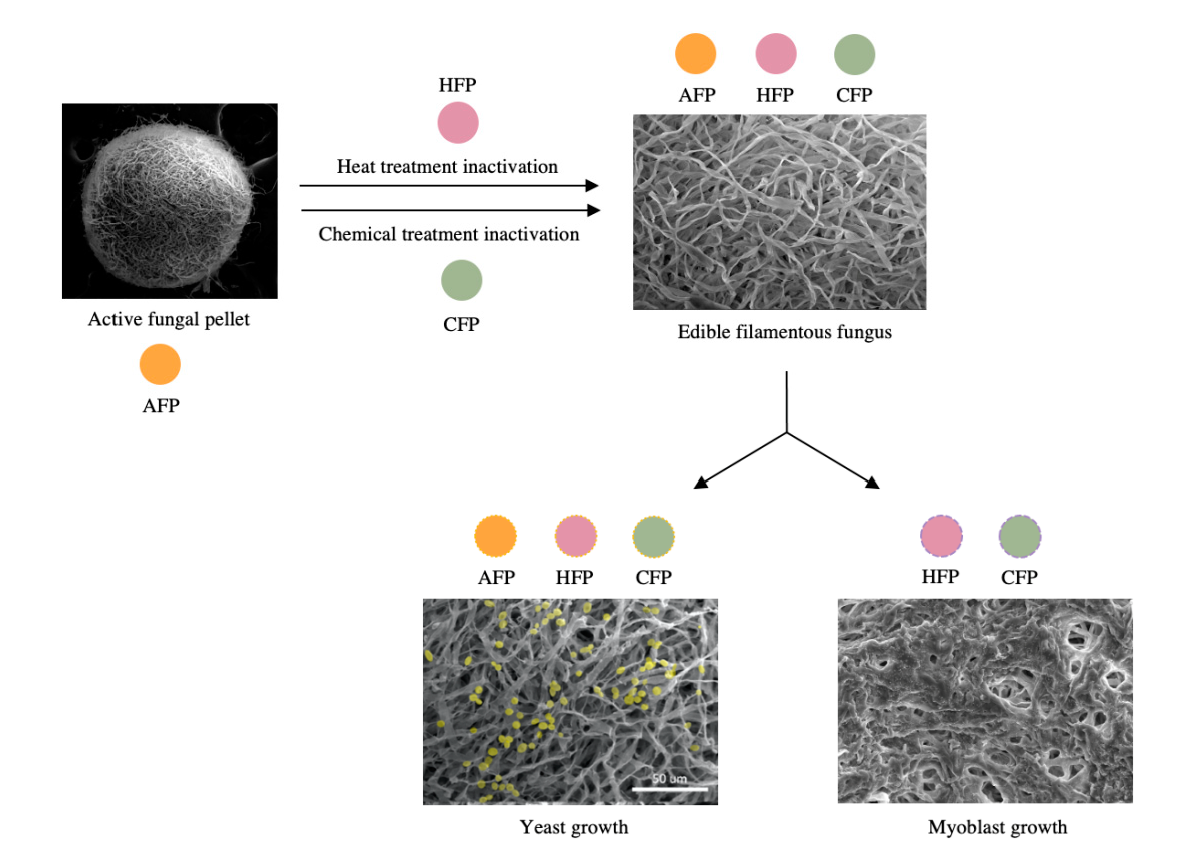
2.2. Fungal Pellet Formation and Inactivation
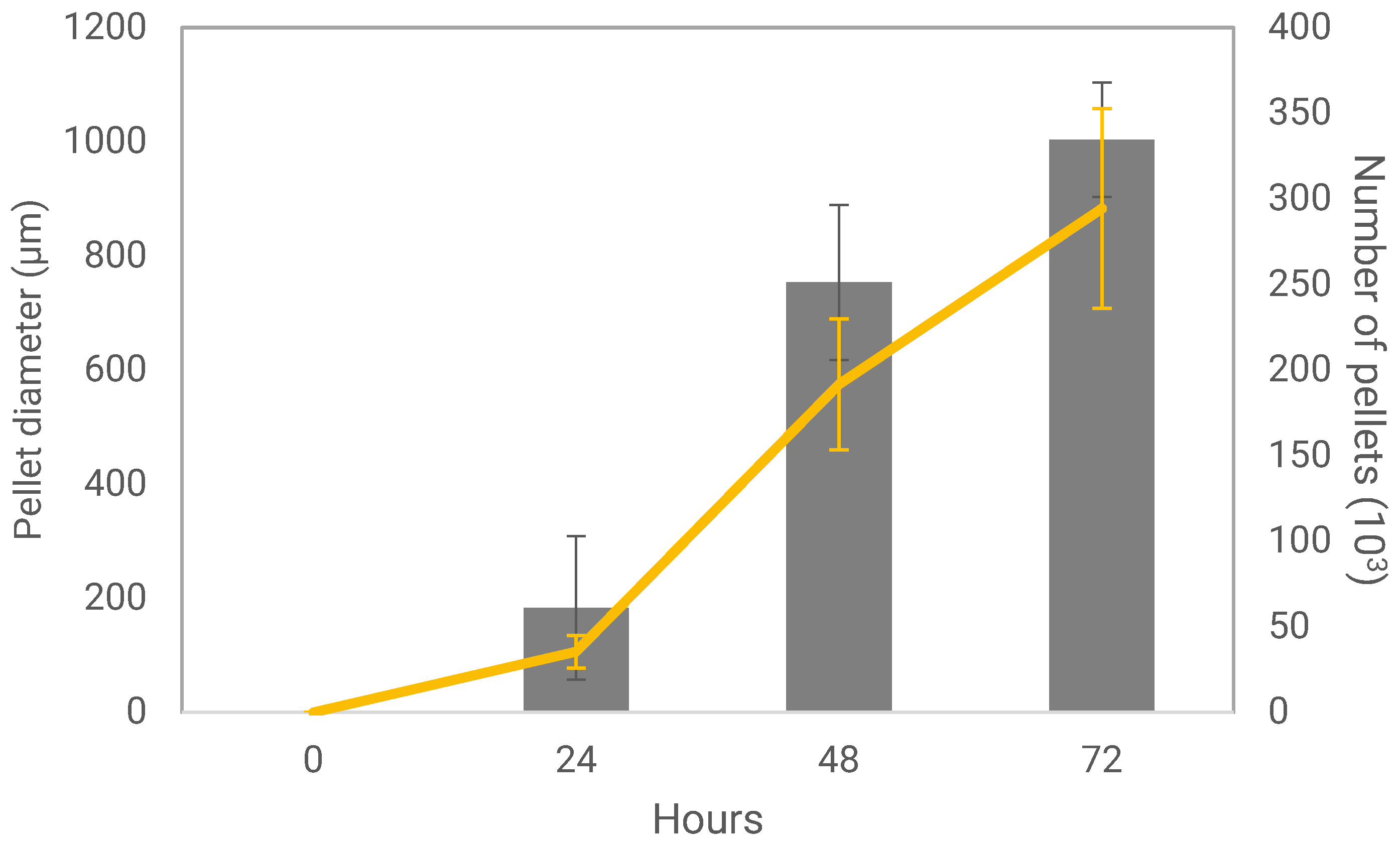
2.3. Measurement of Pellet Parameters
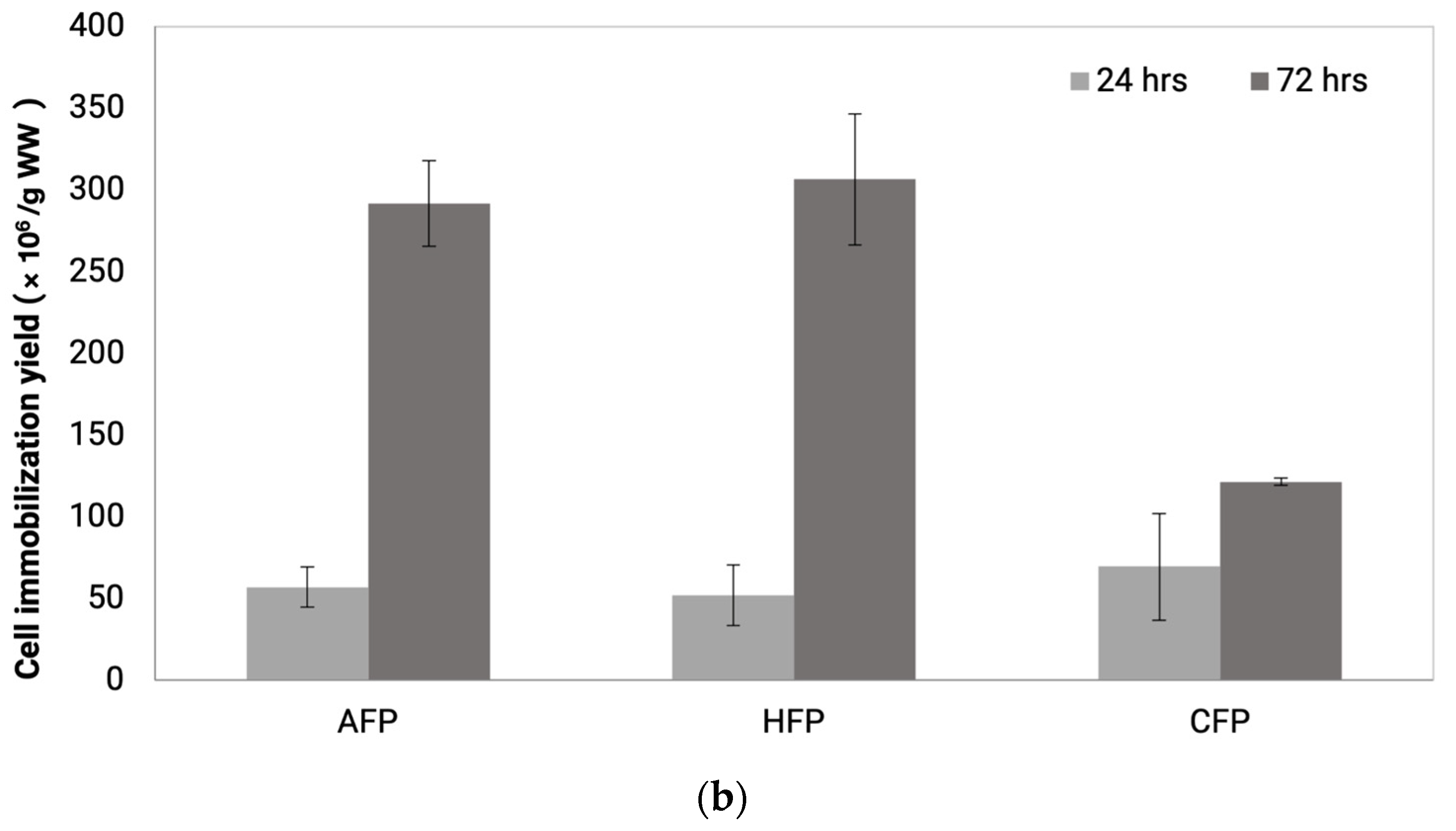
2.4. Measurement of Yeast Cell Immobilization Potential
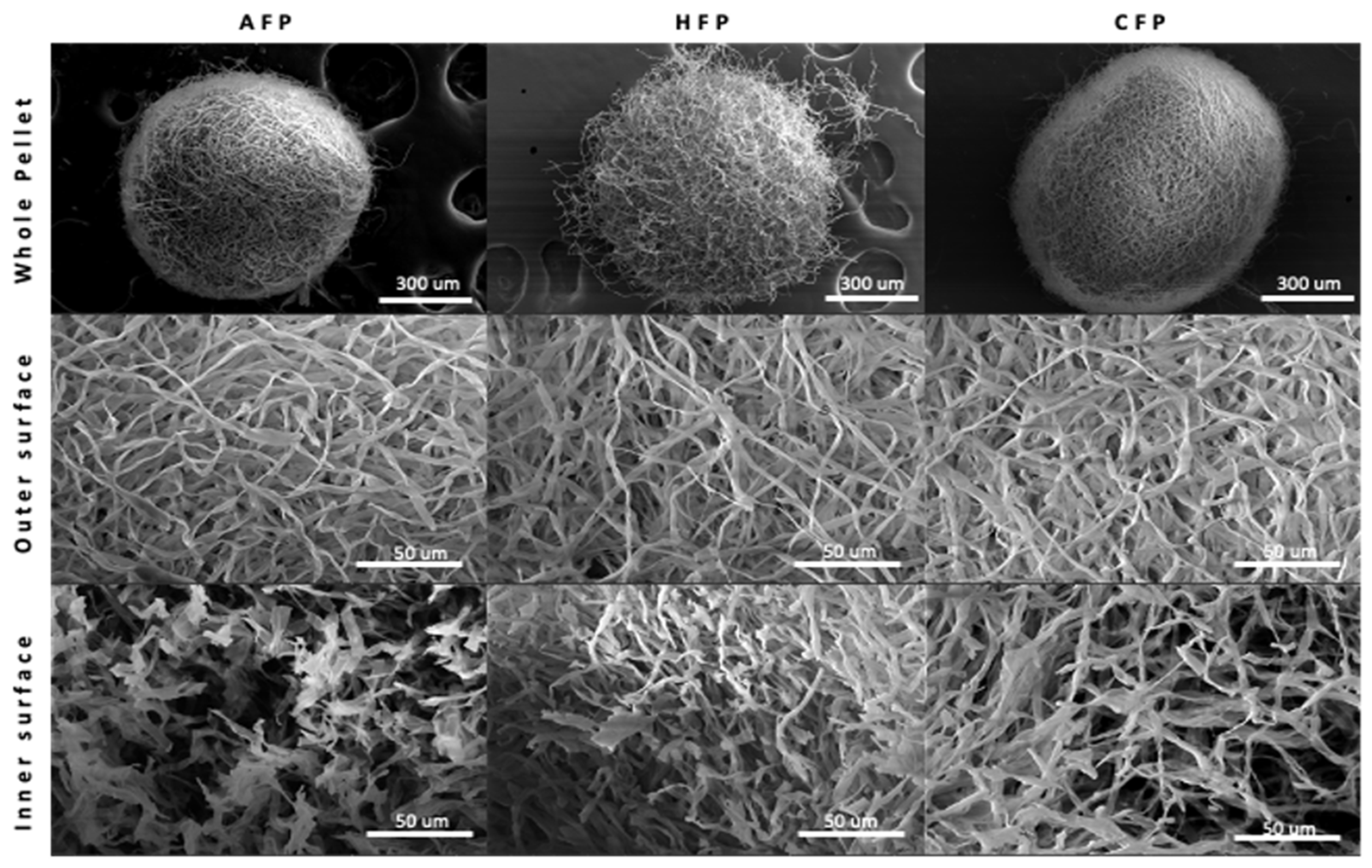
2.5. Microscopic Imaging Analysis
2.6. Measurement of Animal Cell Immobilization Potential
2.7. Statistical Analysis
3. Results
3.1. Fungal Pellet Growth and Characterization
3.2. Yeast Cell Immobilization
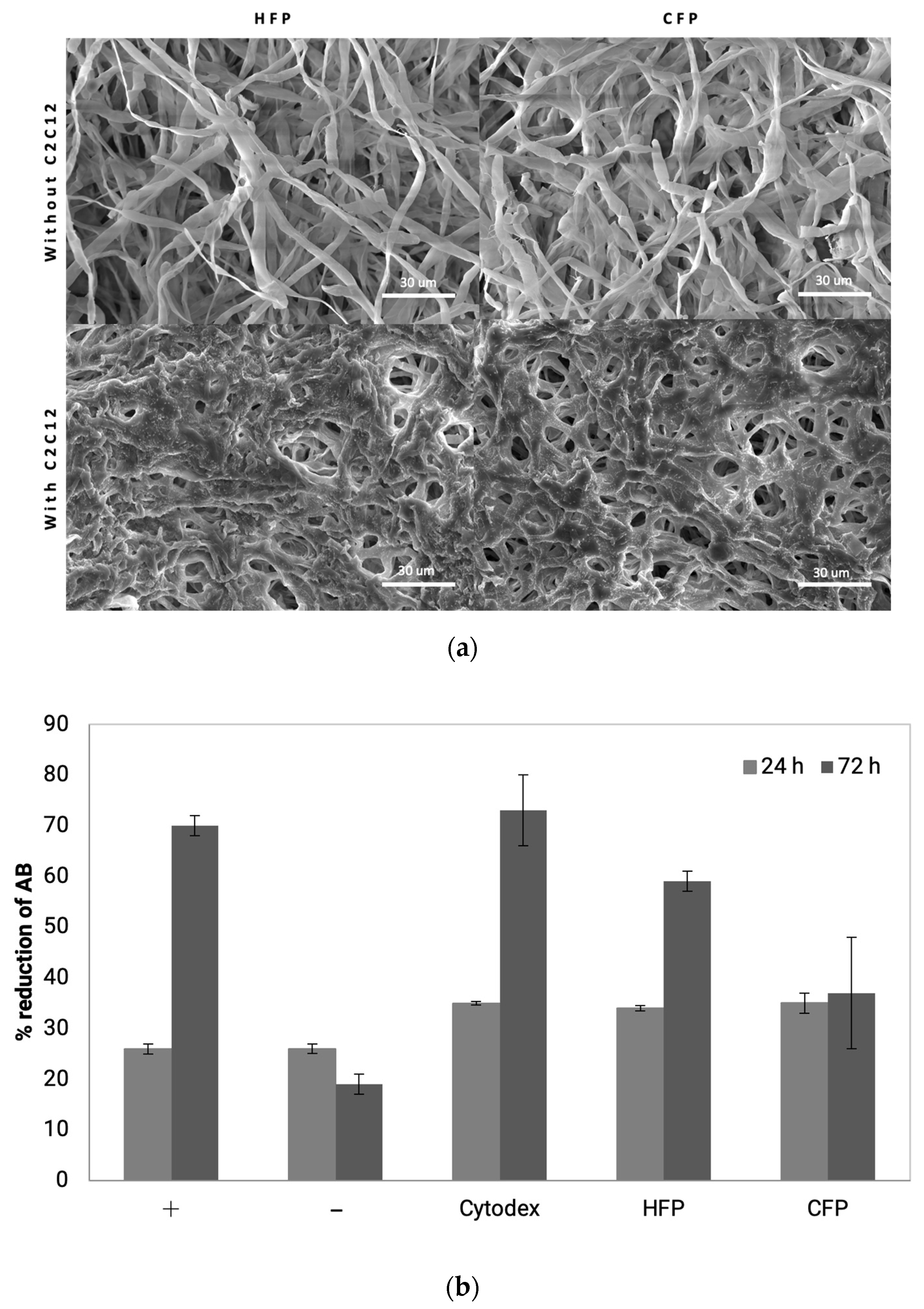
3.3. Animal Cell Immobilization
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karel, S.F.; Libicki, S.B.; Robertson, C.R. The immobilization of whole cells: Engineering principles. Chem. Eng. Sci. 1985, 40, 1321–1354. [Google Scholar] [CrossRef]
- Ravi Kumar, M.N.V. A review of chitin and chitosan applications. React. Funct. Polym. 2000, 46, 1–27. [Google Scholar] [CrossRef]
- Moreno-García, J.; García-Martínez, T.; Mauricio, J.C.; Moreno, J. Yeast immobilization systems for alcoholic wine fermentations: Actual trends and future perspectives. Front. Microbiol. 2018, 9, 241. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y. Immobilized cell fermentation for production of chemicals and fuels. In Bioprocessing for Value-Added Products from Renewable Resources; Elsevier: Amsterdam, The Netherlands, 2007; pp. 373–396. ISBN 9780444521149. [Google Scholar]
- Žnidaršič-Plazl, P. Biocatalytic process intensification via efficient biocatalyst immobilization, miniaturization, and process integration. Curr. Opin. Green Sustain. Chem. 2021, 32, 100546. [Google Scholar] [CrossRef]
- Van de Velde, F.; Lourenço, N.D.; Pinheiro, H.M.; Bakker, M. Carrageenan: A Food-Grade and Biocompatible Support for Immobilisation Techniques. Adv. Synth. Catal. 2002, 344, 815–835. [Google Scholar] [CrossRef]
- Wang, H.-T.; Yang, J.-T.; Chen, K.-I.; Wang, T.-Y.; Lu, T.-J.; Cheng, K.-C. Hydrolyzation of mogrosides: Immobilized β-glucosidase for mogrosides deglycosylation from Lo Han Kuo. Food Sci. Nutr. 2019, 7, 834–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trujillo-Ramírez, D.; Bustos-Vázquez, M.G.; Rodríguez-Durán, L.V.; Torres-de los Santos, R. Universidad Autónoma de Tamaulipas Rice husk (Oryza sativa) as support in the immobilization of yeast cells. Rev. Mex. Ing. Quim. 2021, 21, 1–8. [Google Scholar] [CrossRef]
- Vasilieva, S.; Lobakova, E.; Grigoriev, T.; Selyakh, I.; Semenova, L.; Chivkunova, O.; Gotovtsev, P.; Antipova, C.; Zagoskin, Y.; Scherbakov, P.; et al. Bio-inspired materials for nutrient biocapture from wastewater: Microalgal cells immobilized on chitosan-based carriers. J. Water Process Eng. 2020, 40, 101774. [Google Scholar] [CrossRef]
- Jian, H.; Gao, L.; Guo, Z.; Yang, N.; Liu, N.; Lei, H. Immobilization of lager yeast by hydrocolloids as supporting matrix for improving fermentation performance of high gravity brewing. Ind. Crop. Prod. 2022, 187, 115340. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, J. The filamentous fungal pellet and forces driving its formation. Crit. Rev. Biotechnol. 2016, 36, 1066–1077. [Google Scholar] [CrossRef] [PubMed]
- Veiter, L.; Rajamanickam, V.; Herwig, C. The filamentous fungal pellet-relationship between morphology and productivity. Appl. Microbiol. Biotechnol. 2018, 102, 2997–3006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira, M.T.; Sanromán, A.; Feijoo, G.; Lema, J.M. Control of pellet morphology of filamentous fungi in fluidized bed bioreactors by means of a pulsing flow. Application to Aspergillus niger and Phanerochaete chrysosporium. Enzym. Microb. Technol. 1996, 19, 261–266. [Google Scholar] [CrossRef]
- Ogawa, M.; Bisson, L.F.; García-Martínez, T.; Mauricio, J.C.; Moreno-García, J. New insights on yeast and filamentous fungus adhesion in a natural co-immobilization system: Proposed advances and applications in wine industry. Appl. Microbiol. Biotechnol. 2019, 103, 4723–4731. [Google Scholar] [CrossRef] [Green Version]
- García-Martínez, T.; Peinado, R.A.; Moreno, J.; García-García, I.; Mauricio, J.C. Co-culture of Penicillium chrysogenum and Saccharomyces cerevisiae leading to the immobilization of yeast. J. Chem. Technol. Biotechnol. 2011, 86, 812–817. [Google Scholar] [CrossRef]
- Chacón-Navarrete, H.; Martín, C.; Moreno-García, J. Yeast immobilization systems for second-generation ethanol production: Actual trends and future perspectives. Biofuels Bioprod. Biorefining 2021, 15, 1549–1565. [Google Scholar] [CrossRef]
- Peinado, R.A.; Garcia Mauricio, J.C.; Vigara, J.M.; Ruíz, J.M.; Carnicer, M.M.; García, J.M.; Pérez, M.C.; Riego, M.M.; Calero, L.Z.; Cañete, M.L.; et al. Method of Obtaining Yeast Biocapsules, Biocapsules Thus Obtained and Applications of Same. Patent WO2004029240 A, 8 April 2004. [Google Scholar]
- Peinado, R.A.; Moreno, J.J.; Villalba, J.M.; Villalba, J.M.; González-Reyes, J.A.; Ortega, J.M.; Mauricio, J.C. Yeast biocapsules: A new immobilization method and their applications. Enzym. Microb. Technol. 2006, 40, 79–84. [Google Scholar] [CrossRef]
- Bodiou, V.; Moutsatsou, P.; Post, M.J. Microcarriers for upscaling cultured meat production. Front. Nutr. 2020, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Bock, A.; Sann, H.; Schulze-Horsel, J.; Genzel, Y.; Reichl, U.; Möhler, L. Growth behavior of number distributed adherent MDCK cells for optimization in microcarrier cultures. Biotechnol. Prog. 2009, 25, 1717–1731. [Google Scholar] [CrossRef]
- Rousta, N.; Hellwig, C.; Wainaina, S.; Lukitawesa, L.; Agnihotri, S.; Rousta, K.; Taherzadeh, M.J. Filamentous Fungus Aspergillus oryzae for Food: From Submerged Cultivation to Fungal Burgers and Their Sensory Evaluation-A Pilot Study. Foods 2021, 10, 2774. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, M.; Gutnick, D.; Rosenberg, E. Adherence of bacteria to hydrocarbons: A simple method for measuring cell-surface hydrophobicity. FEMS Microbiol. Lett. 1980, 9, 29–33. [Google Scholar] [CrossRef]
- Kyriakou, M.; Chatziiona, V.K.; Costa, C.N.; Kallis, M.; Koutsokeras, L.; Constantinides, G.; Koutinas, M. Biowaste-based biochar: A new strategy for fermentative bioethanol overproduction via whole-cell immobilization. Appl. Energy 2019, 242, 480–491. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, W.; Zhang, H.; Wang, Z.; Fan, C.; Zang, L. Recent achievements in enhancing anaerobic digestion with carbon- based functional materials. Bioresour. Technol. 2018, 266, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Moreno-García, J.; García-Martinez, T.; Moreno, J.; Mauricio, J.C.; Ogawa, M.; Luong, P.; Bisson, L.F. Impact of Yeast Flocculation and Biofilm Formation on Yeast-Fungus Coadhesion in a Novel Immobilization System. Am. J. Enol. Vitic. 2018, 69, 278–288. [Google Scholar] [CrossRef]
- Jeennor, S.; Anantayanon, J.; Panchanawaporn, S.; Chutrakul, C.; Laoteng, K. Morphologically engineered strain of Aspergillus oryzae as a cell chassis for production development of functional lipids. Gene 2019, 718, 144073. [Google Scholar] [CrossRef]
- Li, Z.J.; Shukla, V.; Fordyce, A.P.; Pedersen, A.G.; Wenger, K.S.; Marten, M.R. Fungal morphology and fragmentation behavior in a fed-batch Aspergillus oryzae fermentation at the production scale. Biotechnol. Bioeng. 2000, 70, 300–312. [Google Scholar] [CrossRef]
- Li, L.; Liang, T.; Liu, W.; Liu, Y.; Ma, F. A comprehensive review of the mycelial pellet: Research status, applications, and future prospects. Ind. Eng. Chem. Res. 2020, 59, 16911–16922. [Google Scholar] [CrossRef]
- Chemical Disinfectants|Disinfection & Sterilization Guidelines|Guidelines Library|Infection Control|CDC. Available online: https://www.cdc.gov/infectioncontrol/guidelines/disinfection/disinfection-methods/chemical.html (accessed on 11 May 2022).
- Steam Sterilization|Disinfection & Sterilization Guidelines|Guidelines Library|Infection Control|CDC. Available online: https://www.cdc.gov/infectioncontrol/guidelines/disinfection/sterilization/steam.html (accessed on 11 May 2022).
- López-Menchero, J.R.; Ogawa, M.; Mauricio, J.C.; Moreno, J.; Moreno-García, J. Effect of calcium alginate coating on the cell retention and fermentation of a fungus-yeast immobilization system. LWT 2021, 144, 111250. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Klis, F.M. Flocculation, adhesion and biofilm formation in yeasts. Mol. Microbiol. 2006, 60, 5–15. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Derdelinckx, G.; Verachtert, H.; Delvaux, F.R. Yeast flocculation: What brewers should know. Appl. Microbiol. Biotechnol. 2003, 61, 197–205. [Google Scholar] [CrossRef]
- Garcia-Rubio, R.; de Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The fungal cell wall: Candida, cryptococcus, and aspergillus species. Front. Microbiol. 2019, 10, 2993. [Google Scholar] [CrossRef]
- Ogawa, M.; García-Martínez, T.; Bisson, L.; Mauricio, J.C.; Moreno, J.; Moreno-García, J. Mapping the intracellular metabolome of yeast biocapsules—Spherical structures of yeast attached to fungal pellets. New Biotechnol. 2020, 58, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Bardouille, C.; Lehmann, J.; Heimann, P.; Jockusch, H. Growth and differentiation of permanent and secondary mouse myogenic cell lines on microcarriers. Appl. Microbiol. Biotechnol. 2001, 55, 556–562. [Google Scholar] [CrossRef] [PubMed]
- McKee, C.; Chaudhry, G.R. Advances and challenges in stem cell culture. Colloids Surf. B Biointerfaces 2017, 159, 62–77. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.J.; Jeong, J.; Kong, H. The interplay between cell adhesion cues and curvature of cell adherent alginate microgels in multipotent stem cell culture. Tissue Eng. Part A 2011, 17, 2687–2694. [Google Scholar] [CrossRef]
- Cha, S.H.; Lee, H.J.; Koh, W.-G. Study of myoblast differentiation using multi-dimensional scaffolds consisting of nano and micropatterns. Biomater. Res. 2017, 21, 1. [Google Scholar] [CrossRef] [Green Version]
- Choquet, D.; Felsenfeld, D.P.; Sheetz, M.P. Extracellular matrix rigidity causes strengthening of integrin-cytoskeleton linkages. Cell 1997, 88, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Cukierman, E.; Pankov, R.; Stevens, D.R.; Yamada, K.M. Taking cell-matrix adhesions to the third dimension. Science 2001, 294, 1708–1712. [Google Scholar] [CrossRef]
- Boonen, K.J.M.; Rosaria-Chak, K.Y.; Baaijens, F.P.T.; van der Schaft, D.W.J.; Post, M.J. Essential environmental cues from the satellite cell niche: Optimizing proliferation and differentiation. Am. J. Physiol. Cell Physiol. 2009, 296, C1338–C1345. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AFP | HFP | CFP | |
|---|---|---|---|
| Viability (%) | 100 ± 0 b | 0 ± 0 a | 17 ± 2 a |
| Diameter (mm) | 0.90 ± 0.10 a | 0.97 ± 0.21 a | 1.03 ± 0.25 a |
| Wet weight (mg/fungal pellet) | 0.37 ± 0.05 a | 0.27 ± 0.06 ab | 0.21 ± 0.04 a |
| Dry mass (mg/fungal pellet) | 0.04 ± 0.01 b | 0.03 ± 0.01 b | 0.01 ± 0.01 a |
| Volume (mm3/fungal pellet) | 0.017 ± 0.005 a | 0.014 ± 0.002 a | 0.017 ± 0.001 a |
| Charge or zeta potential (mV) | −4.3 ± 0.6 a | −2.5 ± 0.3 b | −2.4 ± 0.2 b |
| Carbon % on pellet surface | 61 ± 10 a | 53 ± 9 a | 50 ± 8 a |
| HPBI 1 | 4.7 ± 0.1 c | 2.1 ± 0.5 b | 0.7 ± 0.1 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogawa, M.; Moreno García, J.; Nitin, N.; Baar, K.; Block, D.E. Assessing Edible Filamentous Fungal Carriers as Cell Supports for Growth of Yeast and Cultivated Meat. Foods 2022, 11, 3142. https://doi.org/10.3390/foods11193142
Ogawa M, Moreno García J, Nitin N, Baar K, Block DE. Assessing Edible Filamentous Fungal Carriers as Cell Supports for Growth of Yeast and Cultivated Meat. Foods. 2022; 11(19):3142. https://doi.org/10.3390/foods11193142
Chicago/Turabian StyleOgawa, Minami, Jaime Moreno García, Nitin Nitin, Keith Baar, and David E. Block. 2022. "Assessing Edible Filamentous Fungal Carriers as Cell Supports for Growth of Yeast and Cultivated Meat" Foods 11, no. 19: 3142. https://doi.org/10.3390/foods11193142
APA StyleOgawa, M., Moreno García, J., Nitin, N., Baar, K., & Block, D. E. (2022). Assessing Edible Filamentous Fungal Carriers as Cell Supports for Growth of Yeast and Cultivated Meat. Foods, 11(19), 3142. https://doi.org/10.3390/foods11193142