
Influence of Endogenous Factors of Food Matrices on Avidin—Biotin Immunoassays for the Detection of Bacitracin and Colistin in Food



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Preparation of Biotinylated Analytes
2.3. Preparation of the Heterologous Coating Antigen GEL–BT(ae)
2.4. Preparation of the Heterologous Coating Antigen GEL(pi)–PMB
2.5. Competitive ELISAs Based on the Coating Complex SAV–Biotinylated Hapten and Coated Conjugates
2.6. Assessment of AVI and B7 in Food Matrices
3. Results and Discussion
3.1. Assessment of the SAV–Biotinylated Hapten Complexes as Coating Antigens
3.2. Specificity of the Developed SAV–B7–BT and SAV–B7–COL-Based ELISAs
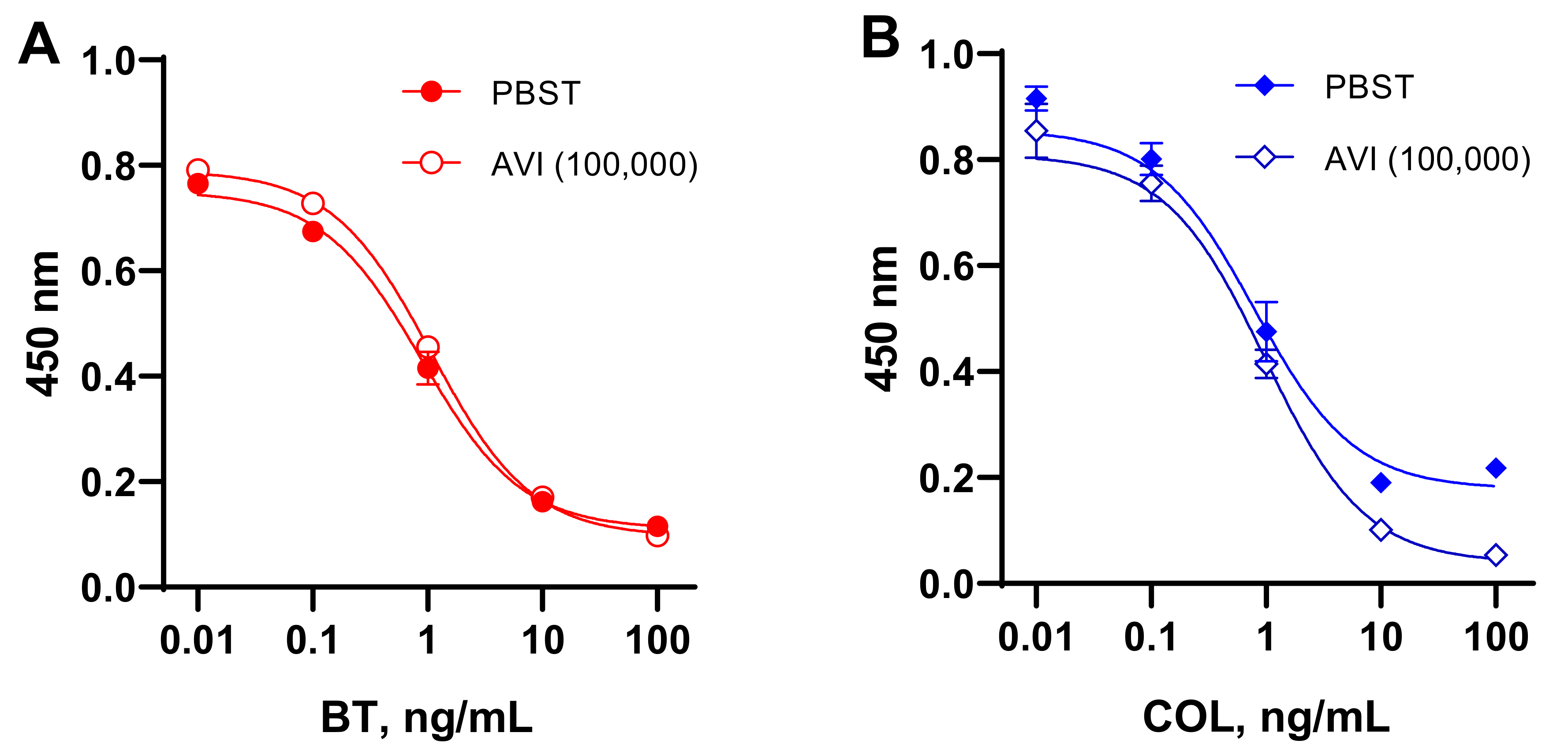
3.3. Assessment of Extraneous Avidin Influence on the Assay
3.4. Assessment of Extraneous Biotin Influence on the Assay
3.5. Influence Assessment of Avidin and Biotin Components from Different Matrices
3.6. Influence of AVI and B7 on the Quantification of BT and COL with ELISAs Based on the Coated SAV–B7–Hapten Complex
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wilchek, M.; Bayer, E.A. [54] Avidin-biotin mediated immunoassays: Overview. Methods Enzymol. 1990, 184, 467–469. [Google Scholar] [PubMed]
- El-Maghrabey, M.; Kishikawa, N.; Harada, S.; Ohyama, K.; Kuroda, N. Quinone-based antibody labeling reagent for enzyme-free chemiluminescent immunoassays. Application to avidin and biotinylated anti-rabbit IgG labeling. Biosens. Bioelectron. 2020, 160, 112215. [Google Scholar] [CrossRef]
- Mustafaoglu, N.; Alves, N.J.; Bilgicer, B. Oriented immobilization of Fab fragments by site-specific biotinylation at the conserved nucleotide binding site for enhanced antigen detection. Langmuir 2015, 31, 9728–9736. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, T.; Mawatari, K.; Kitamori, T. On-chip antibody immobilization for on-demand and rapid immunoassay on a microfluidic chip. Biomicrofluidics 2010, 4, 032207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bratthauer, G.L. The avidin–biotin complex (ABC) method and other avidin–biotin binding methods. In Immunocytochemical Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2010; pp. 257–270. [Google Scholar]
- Cho, I.-H.; Bhunia, A.; Irudayaraj, J. Rapid pathogen detection by lateral-flow immunochromatographic assay with gold nanoparticle-assisted enzyme signal amplification. Int. J. Food Microbiol. 2015, 206, 60–66. [Google Scholar] [CrossRef]
- Cohen, L.; Walt, D.R. Evaluation of antibody biotinylation approaches for enhanced sensitivity of single molecule array (Simoa) immunoassays. Bioconjugate Chem. 2018, 29, 3452–3458. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wagar, E.A.; Meng, Q.H. Comprehensive assessment of biotin interference in immunoassays. Clin. Chim. Acta 2018, 487, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Luong, J.H.; Male, K.B.; Glennon, J.D. Biotin interference in immunoassays based on biotin-strept (avidin) chemistry: An emerging threat. Biotechnol. Adv. 2019, 37, 634–641. [Google Scholar] [CrossRef]
- Zempleni, J.; Wijeratne, S.S.; Hassan, Y.I. Biotin. Biofactors 2009, 35, 36–46. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Beier, R.C.; Luo, P.; Zhai, P.; Wu, N.; Lin, G.; Wang, X.; Xu, G. Analysis of pirlimycin residues in beef muscle, milk, and honey by a biotin–streptavidin-amplified enzyme-linked immunosorbent assay. J. Agric. Food Chem. 2016, 64, 364–370. [Google Scholar] [CrossRef]
- Sun, Z.; Lv, J.; Liu, X.; Tang, Z.; Wang, X.; Xu, Y.; Hammock, B.D. Development of a nanobody-aviTag fusion protein and its application in a streptavidin–biotin-amplified enzyme-linked immunosorbent assay for ochratoxin A in cereal. Anal. Chem. 2018, 90, 10628–10634. [Google Scholar] [CrossRef]
- Lu, M.; Liang, M.; Pan, J.; Zhong, Y.; Zhang, C.; Cui, X.; Wang, T.; Yan, J.; Ding, J.; Zhao, S. Development of a Highly Sensitive Biotin-Streptavidin Amplified Enzyme-Linked Immunosorbent Assay for Determination of Progesterone in Milk Samples. Food Anal. Methods 2021. [CrossRef]
- Zempleni, J.; Mock, D. Biotin biochemistry and human requirements. J. Nutr. Biochem. 1999, 10, 128–138. [Google Scholar] [CrossRef]
- Staggs, C.; Sealey, W.; McCabe, B.; Teague, A.; Mock, D. Determination of the biotin content of select foods using accurate and sensitive HPLC/avidin binding. J. Food Compos. Anal. 2004, 17, 767–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, K.J.; Strominger, J.L. Mechanism of action of bacitracin: Complexation with metal ion and C55-isoprenyl pyrophosphate. Proc. Natl. Acad. Sci. USA 1971, 68, 3223–3227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warren, H.S.; Kania, S.A.; Siber, G. Binding and neutralization of bacterial lipopolysaccharide by colistin nonapeptide. Antimicrob. Agents Chemother. 1985, 28, 107–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, W.; Wang, L.; Usongo, V.; Zhao, X. Colistin induces S. aureus susceptibility to bacitracin. Front. Microbiol. 2018, 9, 2805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regulation, E.C. EU Council Regulation (2010) N 37/2010 of 22 December 2009 on pharmacologically active substances and their classification regardingmaximum residue limits in foodstuffs of animal origin. Off. J. Eur. Union 2010, L15, 1–72. [Google Scholar]
- Hygienic Requirements in Respect of the Safety and Nutritional Value of Foodstuffs, SanPiN 2.3.2.1078-01, as Amended. Chapter 1. Requirements for the Safety and Nutrition of Foods of the Unified Sanitary-Epidemiological and Hygiene Requirements of the Commission of the Customs Union of Russia, Belarus and Kazakhstan. 2019. Available online: http://docs.cntd.ru/document/902249109 (accessed on 10 December 2021).
- Gaugain, M.; Raynaud, A.; Bourcier, S.; Verdon, E.; Hurtaud-Pessel, D. Development of a liquid chromatography-tandem mass spectrometry method to determine colistin, bacitracin and virginiamycin M1 at cross-contamination levels in animal feed. Food Addit. Contam. Part A 2021, 38, 1481–1494. [Google Scholar] [CrossRef]
- Gaudin, V.; Hédou, C.; Rault, A.; Verdon, E.; Soumet, C. Evaluation of three ELISA kits for the screening of colistin residue in porcine and poultry muscle according to the European guideline for the validation of screening methods. Food Addit. Contam. Part A 2020, 37, 1651–1666. [Google Scholar] [CrossRef]
- Li, Y.; Jin, G.; Liu, L.; Kuang, H.; Xiao, J.; Xu, C. A portable fluorescent microsphere-based lateral flow immunosensor for the simultaneous detection of colistin and bacitracin in milk. Analyst 2020, 145, 7884–7892. [Google Scholar] [CrossRef] [PubMed]
- Byzova, N.A.; Serchenya, T.S.; Vashkevich, I.I.; Zherdev, A.V.; Sviridov, O.V.; Dzantiev, B.B. Lateral flow immunoassay for rapid qualitative and quantitative control of the veterinary drug bacitracin in milk. Microchem. J. 2020, 156, 104884. [Google Scholar] [CrossRef]
- Galvidis, I.A.; Eremin, S.A.; Burkin, M.A. Development of indirect competitive enzyme-linked immunoassay of colistin for milk and egg analysis. Food Agric. Immunol. 2020, 31, 424–434. [Google Scholar] [CrossRef] [Green Version]
- Kononenko, G.; Burkin, A. Methods of sanitary surveillance for livestock production. II. Enzyme immunoassay (EIA) of bacitracin. Agric. Biol. 2010; 6, 88–93. [Google Scholar]
- Steinbuch, M.; Audran, R. The isolation of IgG from mammalian sera with the aid of caprylic acid. Arch. Biochem. Biophys. 1969, 134, 279–284. [Google Scholar] [CrossRef]
- Galvidis, I.A.; Burkin, K.M.; Eremin, S.A.; Burkin, M.A. Group-specific detection of 2-deoxystreptamine aminoglycosides in honey based on antibodies against ribostamycin. Anal. Methods 2019, 11, 4620–4628. [Google Scholar] [CrossRef]
- Burkin, M.; Galvidis, I. Simultaneous and differential determination of drugs and metabolites using the same antibody: Difloxacin and sarafloxacin case. Anal. Methods 2016, 8, 5843–5850. [Google Scholar] [CrossRef]
- Burkin, M.; Galvidis, I. Immunochemical detection of apramycin as a contaminant in tissues of edible animals. Food Control 2013, 34, 408–413. [Google Scholar] [CrossRef]
- Burkin, M.; Galvidis, I. Development and application of indirect competitive enzyme immunoassay for detection of neomycin in milk. Appl. Biochem. Microbiol. 2011, 47, 321–326. [Google Scholar] [CrossRef]
- Burkin, M.A.; Galvidis, I.A.; Surovoy, Y.A.; Plyushchenko, I.V.; Rodin, I.A.; Tsarenko, S.V. Development of ELISA formats for polymyxin B monitoring in serum of critically ill patients. J. Pharm. Biomed. Anal. 2021, 204, 114275. [Google Scholar] [CrossRef]
- Green, N.M. Avidin. In Advances in Protein Chemistry; Elsevier: Amsterdam, The Netherlands, 1975; Volume 29, pp. 85–133. [Google Scholar]
- Taskinen, B.; Zmurko, J.; Ojanen, M.; Kukkurainen, S.; Parthiban, M.; Määttä, J.A.; Leppiniemi, J.; Jänis, J.; Parikka, M.; Turpeinen, H. Zebavidin-An avidin-like protein from zebrafish. PLoS ONE 2013, 8, e77207. [Google Scholar] [CrossRef]
- Määttä, J.A.; Helppolainen, S.H.; Hytönen, V.P.; Johnson, M.S.; Kulomaa, M.S.; Airenne, T.T.; Nordlund, H.R. Structural and functional characteristics of xenavidin, the first frog avidin from Xenopus tropicalis. BMC Struct. Biol. 2009, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaiet, L.; Wolf, F.J. The properties of streptavidin, a biotin-binding protein produced by Streptomycetes. Arch. Biochem. Biophys. 1964, 106, 1–5. [Google Scholar] [CrossRef]
- Helppolainen, S.H.; Nurminen, K.P.; Määttä, J.A.; Halling, K.K.; Slotte, J.P.; Huhtala, T.; Liimatainen, T.; Ylä-Herttuala, S.; Airenne, K.J.; Närvänen, A. Rhizavidin from Rhizobium etli: The first natural dimer in the avidin protein family. Biochem. J. 2007, 405, 397–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takakura, Y.; Tsunashima, M.; Suzuki, J.; Usami, S.; Kakuta, Y.; Okino, N.; Ito, M.; Yamamoto, T. Tamavidins–novel avidin-like biotin-binding proteins from the Tamogitake mushroom. FEBS J. 2009, 276, 1383–1397. [Google Scholar] [CrossRef] [PubMed]
- Takakura, Y.; Sofuku, K.; Tsunashima, M.; Kuwata, S. Lentiavidins: Novel avidin-like proteins with low isoelectric points from shiitake mushroom (Lentinula edodes). J. Biosci. Bioeng. 2016, 121, 420–423. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Foodstuffs | MRLs (µg/kg) in EU [19] | MRLs (µg/kg) in EAEU [20] | ||
|---|---|---|---|---|
| BT | COL | BT | COL | |
| Milk | 100 | 50 | NE ** | 50 |
| Eggs | NE | 300 | <20 | 300 |
| Meat, Fish, Poultry | 150 * | 150 | <20 | 150 |
| Liver | 150 * | 150 | <20 | 150 |
| Kidney | 150 * | 200 | <20 | 200 |
| Analyte | Competitive ELISA Based on Coated Complexes | |||
|---|---|---|---|---|
| SAV–B7–BT | SAV–B7–COL | |||
| IC50, ng/mL | CR,% | IC50, ng/mL | CR,% | |
| Bacitracin A | 0.57 | 100 | >10,000 | <0.01 |
| Zn-bacitracin | 0.85 | 67 | >10,000 | <0.01 |
| Colistin | >10,000 | <0.01 | 0.34 | 100 |
| Colistin MS | >10,000 | <0.01 | 9.2 | 3.7 |
| Polymixin B | >10,000 | <0.01 | 1.5 | 22.4 |
| Actinomycin D | >10,000 | <0.01 | >10,000 | <0.01 |
| Vancomicin | >10,000 | <0.01 | >10,000 | <0.01 |
| Virginiamycin M1 | >10,000 | <0.01 | >10,000 | <0.01 |
| Virginiamycin S1 | >10,000 | <0.01 | >10,000 | <0.01 |
| Tetracyclin | >10,000 | <0.01 | >10,000 | <0.01 |
| Erythromycin | >10,000 | <0.01 | >10,000 | <0.01 |
| Tylosin | >10,000 | <0.01 | >10,000 | <0.01 |
| Lasalocid | >10,000 | <0.01 | >10,000 | <0.01 |
| Salinomycin | >10,000 | <0.01 | >10,000 | <0.01 |
| Lincomycin | >10,000 | <0.01 | >10,000 | <0.01 |
| Neomycin | >10,000 | <0.01 | >10,000 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burkin, M.A.; Galvidis, I.A.; Eremin, S.A. Influence of Endogenous Factors of Food Matrices on Avidin—Biotin Immunoassays for the Detection of Bacitracin and Colistin in Food. Foods 2022, 11, 219. https://doi.org/10.3390/foods11020219
Burkin MA, Galvidis IA, Eremin SA. Influence of Endogenous Factors of Food Matrices on Avidin—Biotin Immunoassays for the Detection of Bacitracin and Colistin in Food. Foods. 2022; 11(2):219. https://doi.org/10.3390/foods11020219
Chicago/Turabian StyleBurkin, Maksim A., Inna A. Galvidis, and Sergei A. Eremin. 2022. "Influence of Endogenous Factors of Food Matrices on Avidin—Biotin Immunoassays for the Detection of Bacitracin and Colistin in Food" Foods 11, no. 2: 219. https://doi.org/10.3390/foods11020219
APA StyleBurkin, M. A., Galvidis, I. A., & Eremin, S. A. (2022). Influence of Endogenous Factors of Food Matrices on Avidin—Biotin Immunoassays for the Detection of Bacitracin and Colistin in Food. Foods, 11(2), 219. https://doi.org/10.3390/foods11020219