
Aloe Vera-Fermented Beverage Ameliorates Obesity and Gut Dysbiosis in High-Fat-Diet Mice


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
2.2. Preparation and Composition of the Fermented Juice
2.3. Preparation of Palmitic Acid and AFB Dilution
2.4. Cell Culture and Assay
2.5. Oil Red O Staining
2.6. Animal Experimental Design
2.7. Serum Biochemical Analysis
2.8. Histopathological Examinations
2.9. Analysis of Short-Chain Fatty Acids (SCFA)
2.10. Fecal DNA Extraction and Gut Microbiota Analysis
2.11. Statistical Analysis
3. Results
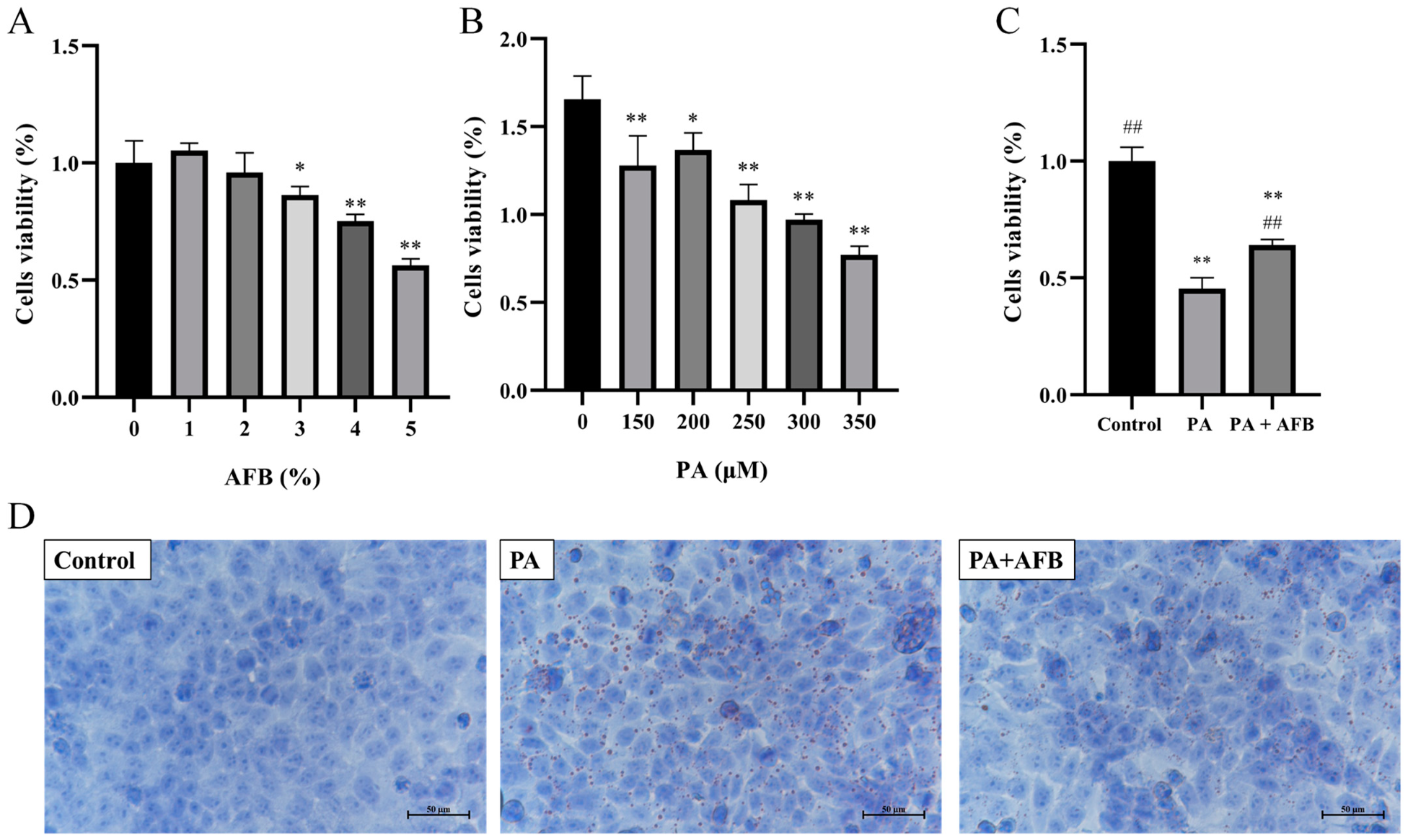
3.1. The Restorative Effects of AFB on HepG2 Cells in High-Fat Environment
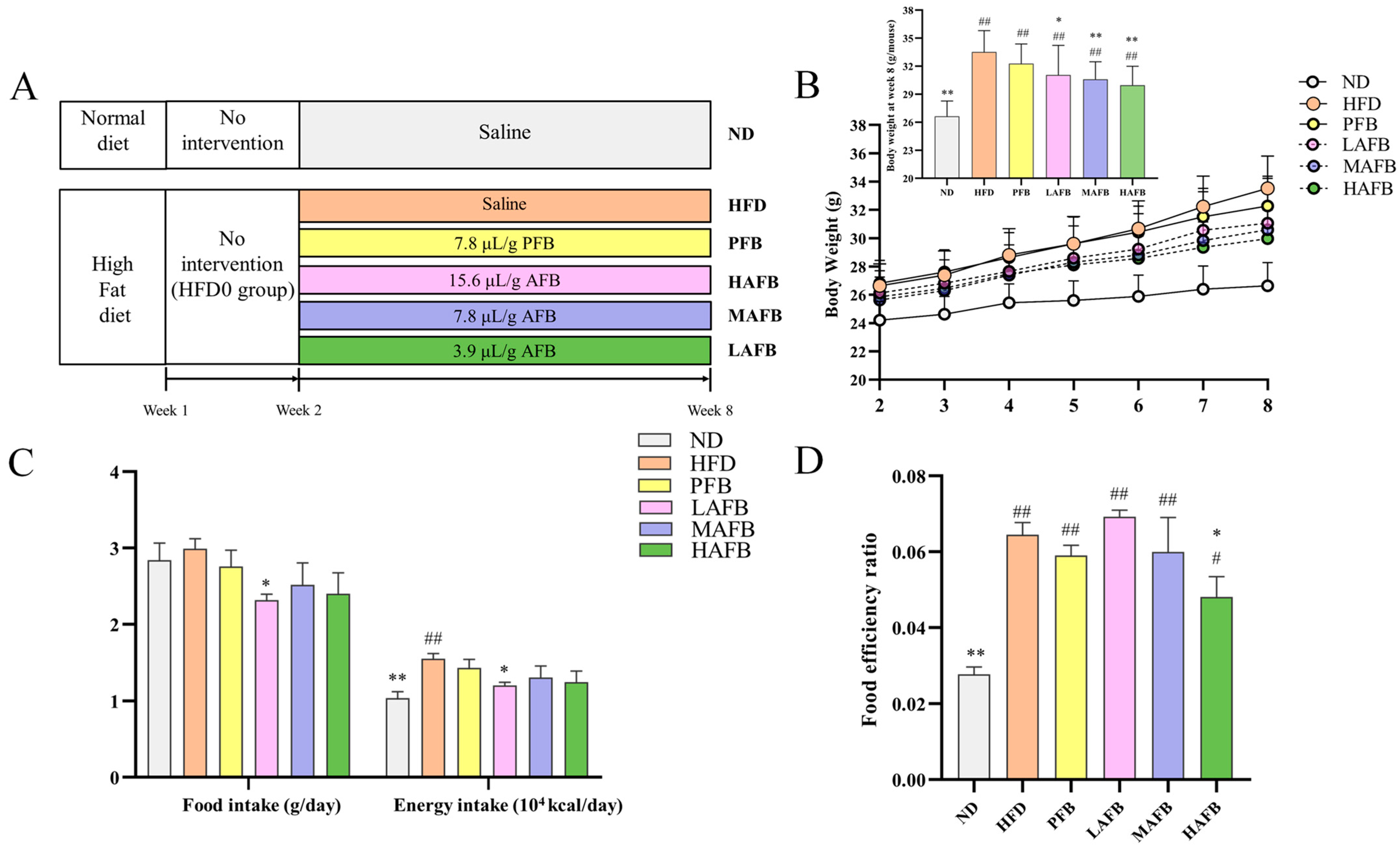
3.2. AFB Prevented HFD-Induced Obesity
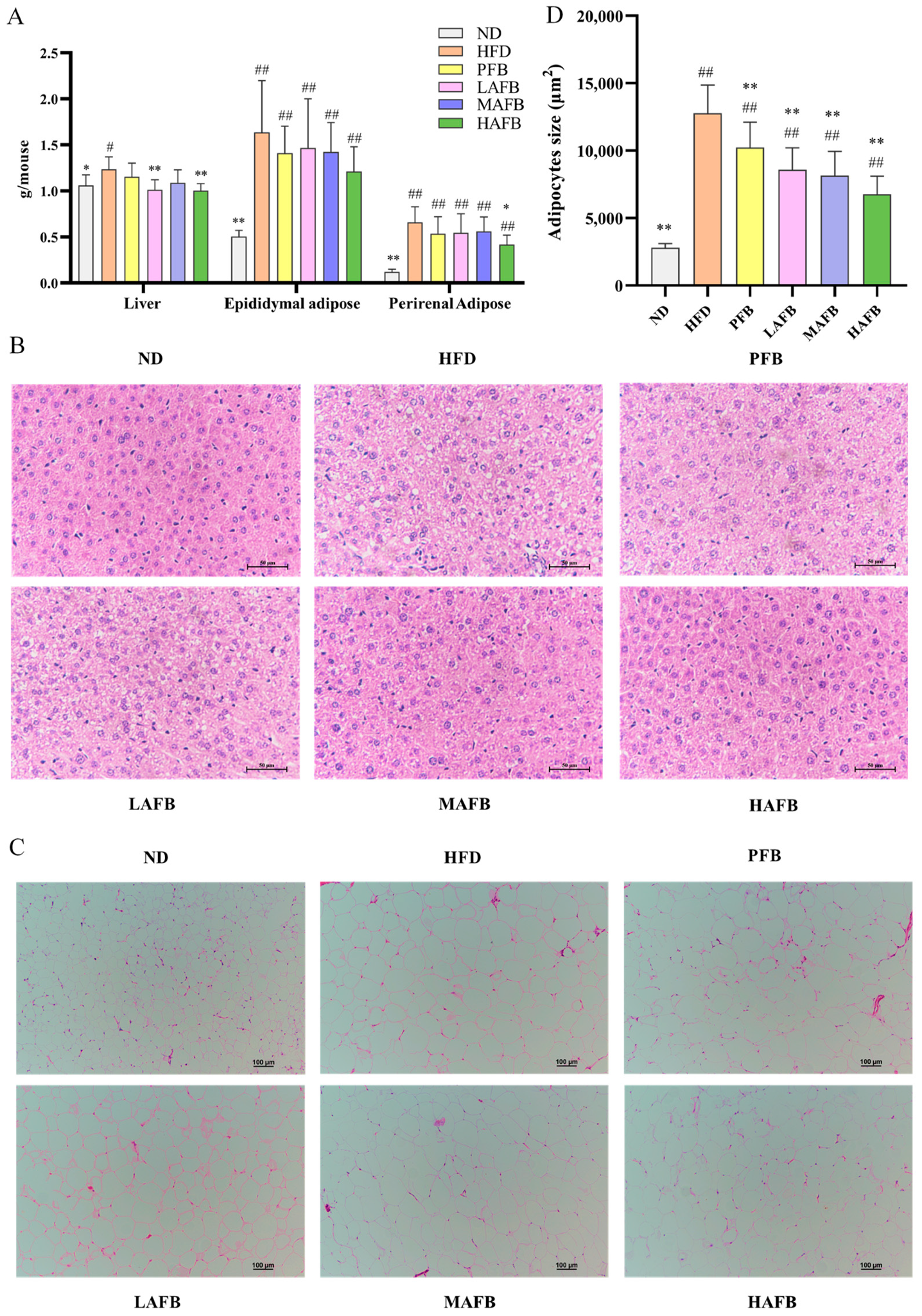
3.3. Effect of AFB on Liver and Adipose Tissue
3.4. Effect of AFB on Serum Biochemical Parameters
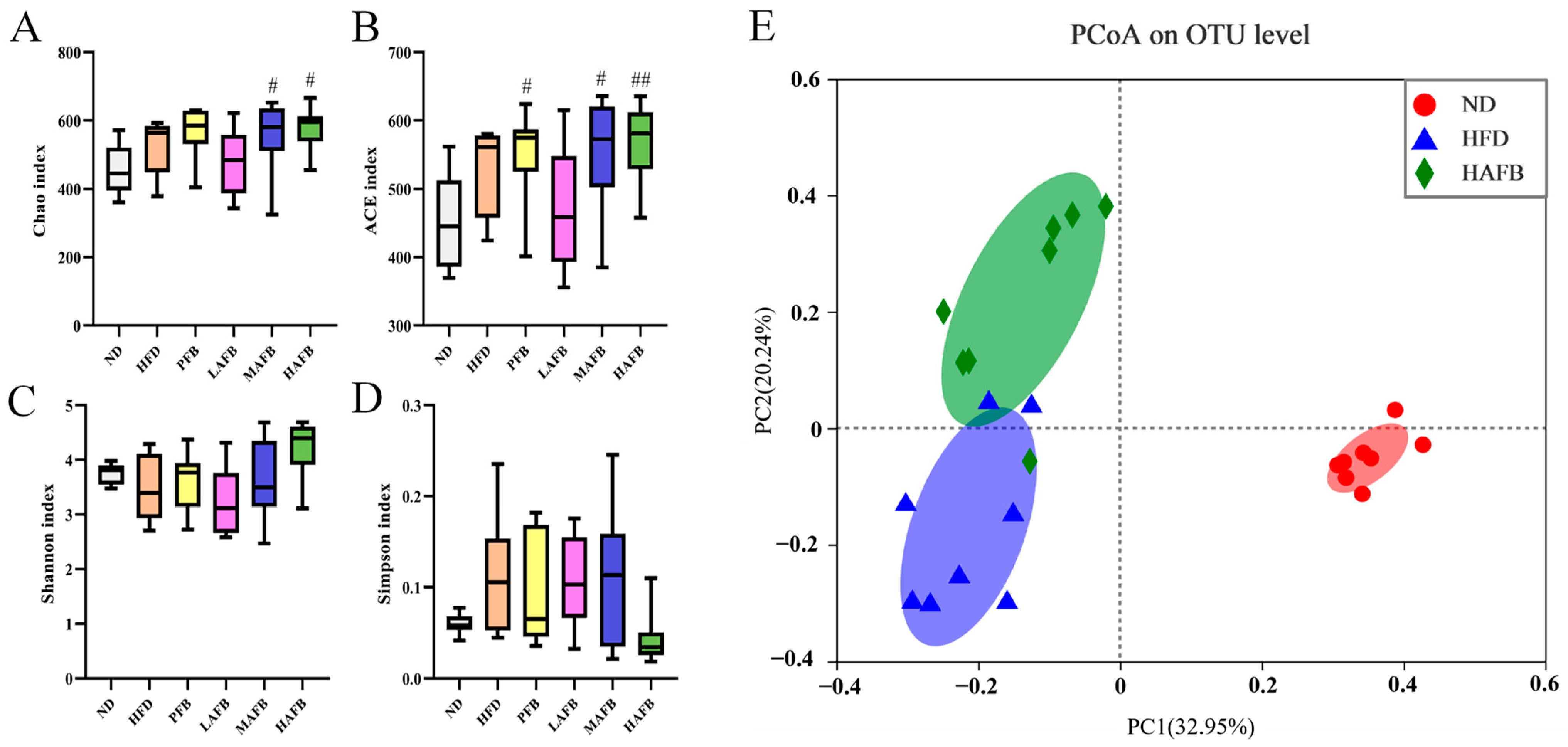
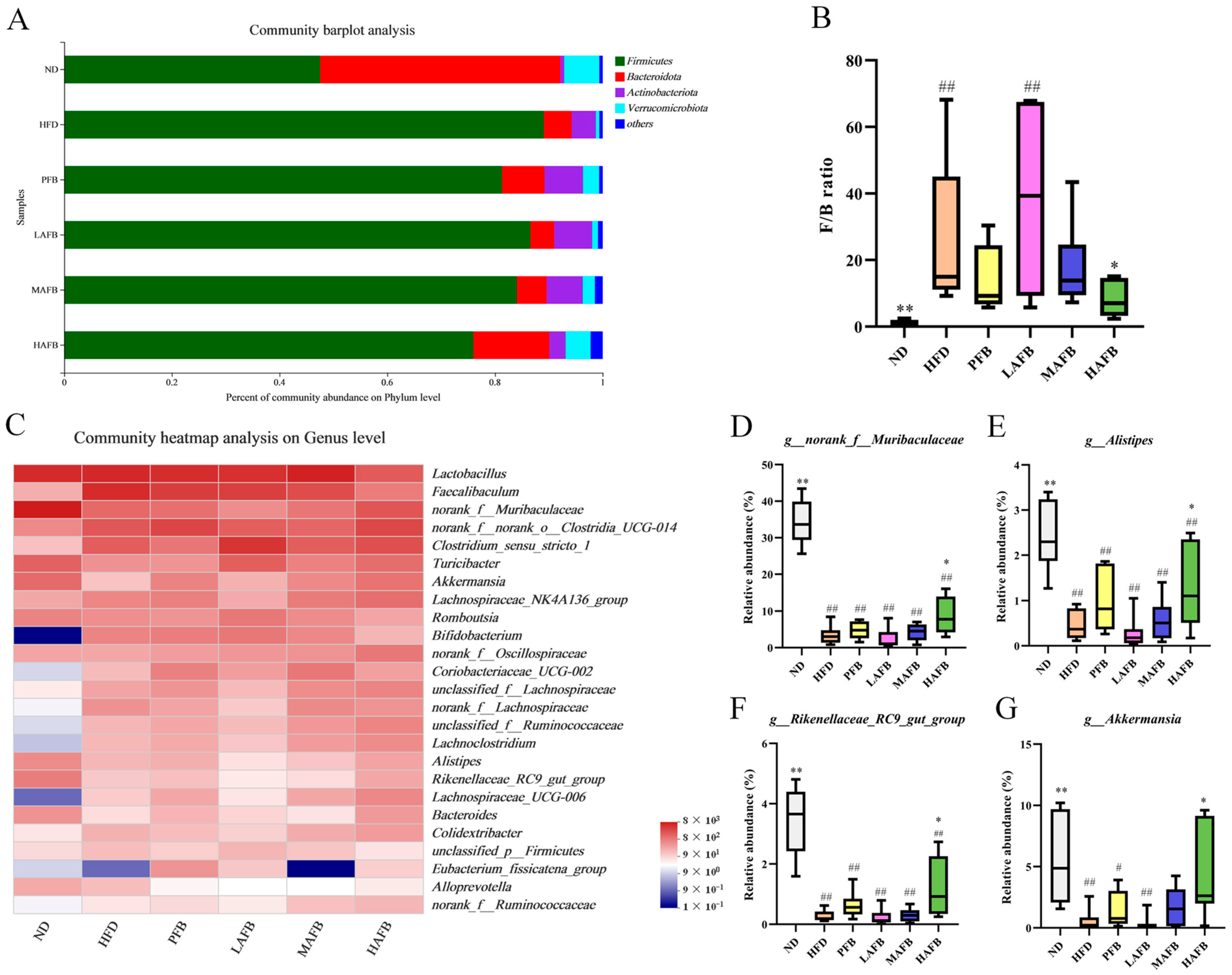
3.5. Effect of AFB Intervention on the α and β Diversities of the Gut Microbiota
3.6. Effect of HFD and AFB on the Composition of the Gut Microbiota
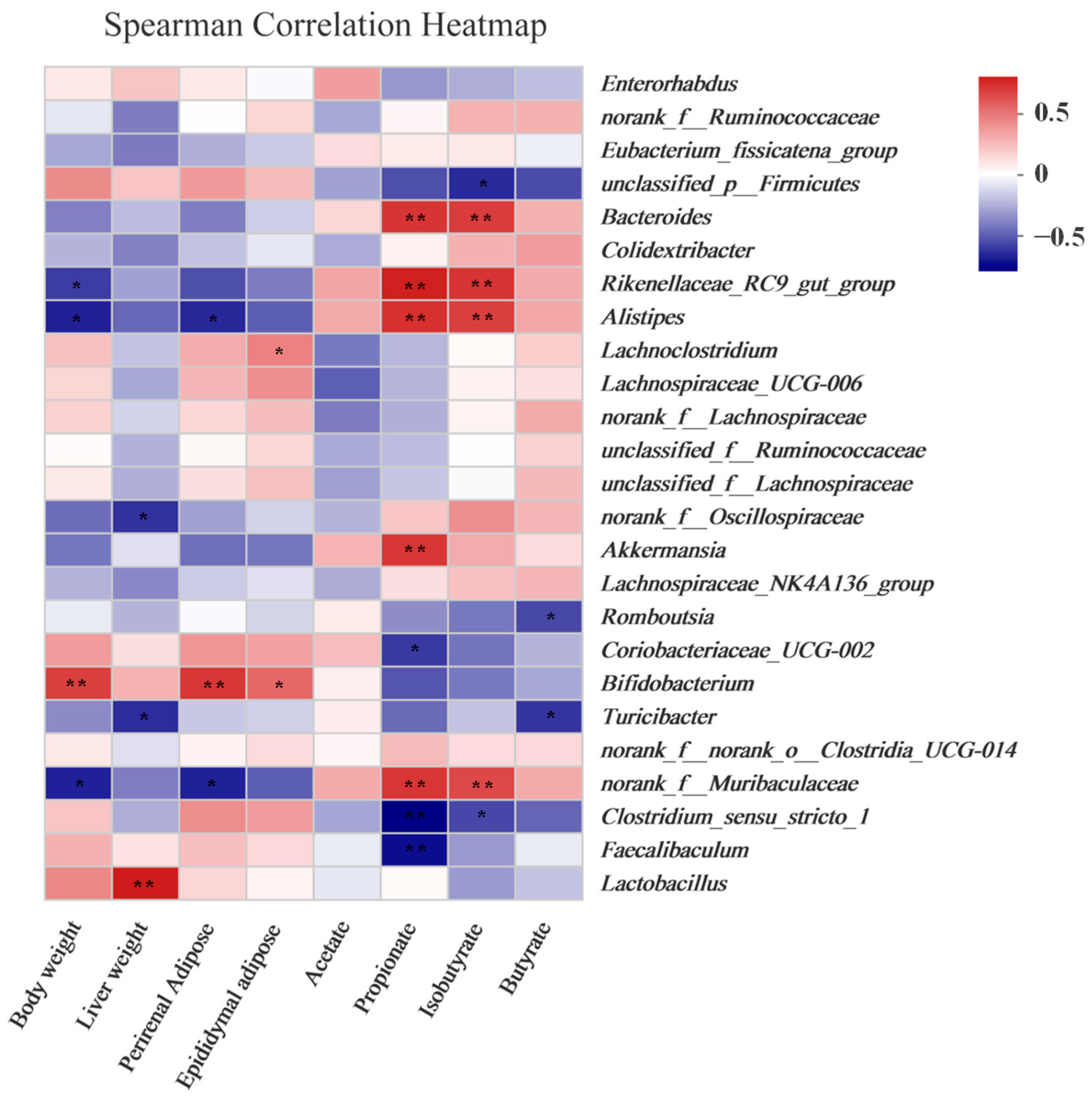
3.7. Obesity and SCFA Levels Were Correlated with the Gut Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hernandez Bautista, R.J.; Mahmoud, A.M.; Konigsberg, M.; Lopez Diaz Guerrero, N.E. Obesity: Pathophysiology, monosodium glutamate-induced model and anti-obesity medicinal plants. Biomed. Pharmacother. 2019, 111, 503–516. [Google Scholar] [CrossRef] [PubMed]
- Ward, Z.J.; Bleich, S.N.; Cradock, A.L.; Barrett, J.L.; Giles, C.M.; Flax, C.; Long, M.W.; Gortmaker, S.L. Projected U.S. State-Level Prevalence of Adult Obesity and Severe Obesity. N. Engl. J. Med. 2019, 381, 2440–2450. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Yoon, J.; Choi, M.S. Tracing the Anti-Inflammatory Mechanism/Triggers of D-Allulose: A Profile Study of Microbiome Composition and mRNA Expression in Diet-Induced Obese Mice. Mol. Nutr. Food Res. 2020, 64, e1900982. [Google Scholar] [CrossRef] [PubMed]
- Ahirwar, R.; Mondal, P.R. Prevalence of obesity in India: A systematic review. Diabetes Metab. Syndr. 2021, 15, 469. [Google Scholar] [CrossRef]
- Tak, Y.J.; Lee, S.Y. Long-Term Efficacy and Safety of Anti-Obesity Treatment: Where Do We Stand? Curr. Obes. Rep. 2021, 10, 14–30. [Google Scholar] [CrossRef] [PubMed]
- Misawa, E.; Tanaka, M.; Nabeshima, K.; Nomaguchi, K.; Yamada, M.; Toida, T.; Iwatsuki, K. Administration of dried Aloe vera gel powder reduced body fat mass in diet-induced obesity (DIO) rats. J. Nutr. Sci. Vitaminol. 2012, 58, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Kuok, K.I.; Jin, Y.; Wang, R. Biomedical applications of Aloe vera. Crit. Rev. Food Sci. Nutr. 2019, 59, S244–S256. [Google Scholar] [CrossRef]
- Kim, K.; Kim, H.; Kwon, J.; Lee, S.; Kong, H.; Im, S.A.; Lee, Y.H.; Lee, Y.R.; Oh, S.T.; Jo, T.H.; et al. Hypoglycemic and hypolipidemic effects of processed Aloe vera gel in a mouse model of non-insulin-dependent diabetes mellitus. Phytomedicine 2009, 16, 856–863. [Google Scholar] [CrossRef]
- Walid, R.; Hafida, M.; Abdelhamid, E.I.; Reda, B.; Rachid, A.; Mohamed, B. Beneficial effects of Aloe vera gel on lipid profile, lipase activities and oxidant/antioxidant status in obese rats. J. Funct. Foods 2018, 48, 525–532. [Google Scholar] [CrossRef]
- Misawa, E.; Tanaka, M.; Nomaguchi, K.; Nabeshima, K.; Yamada, M.; Toida, T.; Iwatsuki, K. Oral ingestion of Aloe vera phytosterols alters hepatic gene expression profiles and ameliorates obesity-associated metabolic disorders in zucker diabetic fatty rats. J. Agric. Food Chem. 2012, 60, 2799–2806. [Google Scholar] [CrossRef]
- Tada, A.; Misawa, E.; Tanaka, M.; Saito, M.; Nabeshima, K.; Yamauchi, K.; Abe, F.; Goto, T.; Kawada, T. Investigating Anti-Obesity Effects by Oral Administration of Aloe vera Gel Extract (AVGE): Possible Involvement in Activation of Brown Adipose Tissue (BAT). J. Nutr. Sci. Vitaminol. 2020, 66, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Tanaka, M.; Misawa, E.; Yamada, M.; Yamauchi, K.; Iwatsuki, K. Aloe vera gel extract attenuates ethanol-induced hepatic lipid accumulation by suppressing the expression of lipogenic genes in mice. Biosci. Biotechnol. Biochem. 2012, 76, 2049–2054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pothuraju, R.; Sharma, R.K.; Chagalamarri, J.; Kavadi, P.K.; Jangra, S. Influence of milk fermented with Lactobacillus rhamnosus NCDC 17 alone and in combination with herbal ingredients on diet induced adiposity and related gene expression in C57BL/6J mice. Food Funct. 2015, 6, 3576–3584. [Google Scholar] [CrossRef] [PubMed]
- Ray, M.; Hor, P.K.; Ojha, D.; Soren, J.P.; Singh, S.N.; Mondal, K.C. Bifidobacteria and its rice fermented products on diet induced obese mice: Analysis of physical status, serum profile and gene expressions. Benef. Microbes 2018, 9, 441–452. [Google Scholar] [CrossRef]
- Park, S.; Son, H.K.; Chang, H.C.; Lee, J.J. Effects of Cabbage-Apple Juice Fermented by Lactobacillus plantarum EM on Lipid Profile Improvement and Obesity Amelioration in Rats. Nutrients 2020, 12, 1135. [Google Scholar] [CrossRef] [Green Version]
- Guan, Q.; Ding, X.W.; Zhong, L.Y.; Zhu, C.; Nie, P.; Song, L.H. Beneficial effects of Lactobacillus-fermented black barley on high fat diet-induced fatty liver in rats. Food Funct. 2021, 12, 6526–6539. [Google Scholar] [CrossRef]
- Shiferaw Terefe, N.; Augustin, M.A. Fermentation for tailoring the technological and health related functionality of food products. Crit. Rev. Food Sci. Nutr. 2020, 60, 2887–2913. [Google Scholar] [CrossRef]
- Vieira, A.D.S.; de Souza, C.B.; Padilha, M.; Zoetendal, E.G.; Smidt, H.; Saad, S.M.I.; Venema, K. Impact of a fermented soy beverage supplemented with acerola by-product on the gut microbiota from lean and obese subjects using an in vitro model of the human colon. Appl. Microbiol. Biotechnol. 2021, 105, 3771–3785. [Google Scholar] [CrossRef]
- Yan, X.; Wang, F.; Weng, P.; Wu, Z. The effect of fermented Huyou juice on intestinal microbiota in a high-fat diet-induced obesity mouse model. J. Food Biochem. 2020, 44, e13480. [Google Scholar] [CrossRef]
- Cox, A.J.; West, N.P.; Cripps, A.W. Obesity, inflammation, and the gut microbiota. Lancet Diabetes Endocrinol. 2015, 3, 207–215. [Google Scholar] [CrossRef]
- Cani, P.D.; Jordan, B.F. Gut microbiota-mediated inflammation in obesity: A link with gastrointestinal cancer. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Torres-Fuentes, C.; Schellekens, H.; Dinan, T.G.; Cryan, J.F. The microbiota-gut-brain axis in obesity. Lancet Gastroenterol. Hepatol. 2017, 2, 747–756. [Google Scholar] [CrossRef]
- Zang, Y.; Fan, L.; Chen, J.; Huang, R.; Qin, H. Improvement of Lipid and Glucose Metabolism by Capsiate in Palmitic Acid-Treated HepG2 Cells via Activation of the AMPK/SIRT1 Signaling Pathway. J. Agric. Food Chem. 2018, 66, 6772–6781. [Google Scholar] [CrossRef]
- Li, Z.T.; Hu, G.A.; Zhu, L.; Zhao, Z.C.; Yun, J.; Gao, M.J.; Zhan, X.B. In vitro digestion and fecal fermentation of highly resistant starch rice and its effect on the gut microbiota. Food Chem. 2021, 361, 130095. [Google Scholar] [CrossRef] [PubMed]
- Polyzos, S.A.; Kountouras, J.; Mantzoros, C.S. Obesity and nonalcoholic fatty liver disease: From pathophysiology to therapeutics. Metabolism 2019, 92, 82–97. [Google Scholar] [CrossRef]
- Kachur, S.; Lavie, C.J.; de Schutter, A.; Milani, R.V.; Ventura, H.O. Obesity and cardiovascular diseases. Minerva Med. 2017, 108, 212–228. [Google Scholar] [CrossRef]
- Fu, Y.; Guo, J. Blood Cholesterol Monitoring With Smartphone as Miniaturized Electrochemical Analyzer for Cardiovascular Disease Prevention. IEEE Trans. Biomed. Circuits Syst. 2018, 12, 784–790. [Google Scholar] [CrossRef]
- Saltiel, A.R.; Olefsky, J.M. Inflammatory mechanisms linking obesity and metabolic disease. J. Clin. Investig. 2017, 127, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Fabbrini, E.; Sullivan, S.; Klein, S. Obesity and nonalcoholic fatty liver disease: Biochemical, metabolic, and clinical implications. Hepatology 2010, 51, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Nomaguchi, K.; Tanaka, M.; Misawa, E.; Yamada, M.; Toida, T.; Iwatsuki, K.; Goto, T.; Kawada, T. Aloe vera phytosterols act as ligands for PPAR and improve the expression levels of PPAR target genes in the livers of mice with diet-induced obesity. Obes. Res. Clin. Pract. 2011, 5, e190–e201. [Google Scholar] [CrossRef]
- Shakib, Z.; Shahraki, N.; Razavi, B.M.; Hosseinzadeh, H. Aloe vera as an herbal medicine in the treatment of metabolic syndrome: A review. Phytother. Res. 2019, 33, 2649–2660. [Google Scholar] [CrossRef] [PubMed]
- Pothuraju, R.; Sharma, R.K.; Onteru, S.K.; Singh, S.; Hussain, S.A. Hypoglycemic and Hypolipidemic Effects of Aloe vera Extract Preparations: A Review. Phytother. Res. 2016, 30, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Abdullah; Deng, L.; Zhao, M.; Tang, J.; Liu, T.; Zhong, H.; Feng, F. Probiotic-fermented blueberry juice prevents obesity and hyperglycemia in high fat diet-fed mice in association with modulating the gut microbiota. Food Funct. 2020, 11, 9192–9207. [Google Scholar] [CrossRef] [PubMed]
- Chai, Z.; Yan, Y.; Zan, S.; Meng, X.; Zhang, F. Probiotic-fermented blueberry pomace alleviates obesity and hyperlipidemia in high-fat diet C57BL/6J mice. Food Res. Int. 2022, 157, 111396. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.C.; Lin, X.H.; Xue, B.; Luo, J.M.; Gao, L.J.; Wang, Y.; Ou, S.Y.; Peng, X.C. Impact of polyphenols combined with high-fat diet on rats’ gut microbiota. J. Funct. Foods 2016, 26, 763–771. [Google Scholar] [CrossRef]
- Sandoval, V.; Sanz-Lamora, H.; Arias, G.; Marrero, P.F.; Haro, D.; Relat, J. Metabolic Impact of Flavonoids Consumption in Obesity: From Central to Peripheral. Nutrients 2020, 12, 2393. [Google Scholar] [CrossRef]
- Kawano, Y.; Cohen, D.E. Mechanisms of hepatic triglyceride accumulation in non-alcoholic fatty liver disease. J. Gastroenterol. 2013, 48, 434–441. [Google Scholar] [CrossRef] [Green Version]
- Al-Khayri, J.M.; Sahana, G.R.; Nagella, P.; Joseph, B.V.; Alessa, F.M.; Al-Mssallem, M.Q. Flavonoids as Potential Anti-Inflammatory Molecules: A Review. Molecules 2022, 27, 2901. [Google Scholar] [CrossRef]
- Ma, S.; Zhang, H.; Xu, J. Characterization, Antioxidant and Anti-Inflammation Capacities of Fermented Flammulina velutipes Polyphenols. Molecules 2021, 26, 6205. [Google Scholar] [CrossRef]
- Guo, X.; Cao, X.; Fang, X.; Guo, A.; Li, E. Inhibitory effects of fermented Ougan (Citrus reticulata cv. Suavissima) juice on high-fat diet-induced obesity associated with white adipose tissue browning and gut microbiota modulation in mice. Food Funct. 2021, 12, 9300–9314. [Google Scholar] [CrossRef]
- Kallus, S.J.; Brandt, L.J. The intestinal microbiota and obesity. J. Clin. Gastroenterol. 2012, 46, 16–24. [Google Scholar] [CrossRef]
- Zhu, L.Y.; Gao, M.L.; Li, H.Y.; Deng, Z.Y.; Zhang, B.; Fan, Y.W. Effects of soluble dietary fiber from sweet potato dregs on the structures of intestinal flora in mice. Food Biosci. 2021, 40, 100880. [Google Scholar] [CrossRef]
- Harada, N.; Minami, Y.; Hanada, K.; Hanaoka, R.; Kobayashi, Y.; Izawa, T.; Sato, T.; Kato, S.; Inui, H.; Yamaji, R. Relationship between gut environment, feces-to-food ratio, and androgen deficiency-induced metabolic disorders. Gut Microbes 2020, 12, 1817719. [Google Scholar] [CrossRef]
- Yuan, G.; Tan, M.; Chen, X. Punicic acid ameliorates obesity and liver steatosis by regulating gut microbiota composition in mice. Food Funct. 2021, 12, 7897–7908. [Google Scholar] [CrossRef]
- Ye, J.; Zhao, Y.; Chen, X.; Zhou, H.; Yang, Y.; Zhang, X.; Huang, Y.; Zhang, N.; Lui, E.M.K.; Xiao, M. Pu-erh tea ameliorates obesity and modulates gut microbiota in high fat diet fed mice. Food Res. Int. 2021, 144, 110360. [Google Scholar] [CrossRef]
- Jing, N.; Liu, X.; Jin, M.; Yang, X.; Hu, X.; Li, C.; Zhao, K. Fubrick tea attenuates high-fat diet induced fat deposition and metabolic disorder by regulating gut microbiota and caffeine metabolism. Food Funct. 2020, 11, 6971–6986. [Google Scholar] [CrossRef]
- Song, H.; Shen, X.; Chu, Q.; Zheng, X. Vaccinium bracteatum Thunb. fruit extract reduces high-fat diet-induced obesity with modulation of the gut microbiota in obese mice. J. Food Biochem. 2021, 45, e13808. [Google Scholar] [CrossRef]
- Lan, Y.; Sun, Q.; Ma, Z.; Peng, J.; Zhang, M.; Wang, C.; Zhang, X.; Yan, X.; Chang, L.; Hou, X.; et al. Seabuckthorn polysaccharide ameliorates high-fat diet-induced obesity by gut microbiota-SCFAs-liver axis. Food Funct. 2022, 13, 2925–2937. [Google Scholar] [CrossRef]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef] [Green Version]
- Sahuri-Arisoylu, M.; Brody, L.P.; Parkinson, J.R.; Parkes, H.; Navaratnam, N.; Miller, A.D.; Thomas, E.L.; Frost, G.; Bell, J.D. Reprogramming of hepatic fat accumulation and ‘browning’ of adipose tissue by the short-chain fatty acid acetate. Int. J. Obes. 2016, 40, 955–963. [Google Scholar] [CrossRef]
- Canfora, E.E.; Meex, R.C.R.; Venema, K.; Blaak, E.E. Gut microbial metabolites in obesity, NAFLD and T2DM. Nat. Rev. Endocrinol 2019, 15, 261–273. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ND | HFD | PFB | LAFB | MAFB | HAFB | |
|---|---|---|---|---|---|---|
| TC (mmol/L) | 2.54 ± 0.18 ** | 5.08 ± 0.07 ## | 4.71 ± 0.54 ## | 4.83 ± 0.23 ## | 4.61 ± 0.39 ## | 4.30 ± 0.34 ##,** |
| TG (mmol/L) | 0.92 ± 0.08 | 1.00 ± 0.31 | 0.96 ± 0.11 | 0.86 ± 0.15 | 0.77 ± 0.19 | 0.74 ± 0.07 |
| HDL-C (mmol/L) | 2.10 ± 0.18 ** | 3.96 ± 0.15 ## | 3.59 ± 0.41 ## | 3.76 ± 0.12 ## | 3.67 ± 0.10 ## | 3.58 ± 0.26 ## |
| LDL-C (mmol/L) | 0.30 ± 0.06 ** | 0.79 ± 0.30 ## | 0.87 ± 0.19 ## | 0.78 ± 0.11 ## | 0.75 ± 0.03 ## | 0.59 ± 0.10 # |
| ALT (U/L) | 497.25 ± 5.38 | 514.30 ± 21.31 | 473.22 ± 21.23 ** | 478.82 ± 16.57 * | 472.78 ± 4.98 ** | 477.06 ± 20.16 ** |
| AST (U/L) | 103.98 ± 10.92 | 107.76 ± 23.19 | 101.30 ± 14.46 | 91.56 ± 12.15 | 106.66 ± 18.54 | 90.98 ± 17.52 |
| TNF-α (pg/mL) | 17.94 ± 9.19 | 24.26 ± 10.37 | 21.02 ± 9.05 | 20.11 ± 13.95 | 17.09 ± 7.42 | 15.53 ± 7.51 |
| IL-10 (pg/mL) | 52.81 ± 34.82 ** | 215.01 ± 12.55 ## | 75.78 ± 17.98 ** | 66.7 ± 42.56 ** | 54.94 ± 38.16 ** | 40.6 ± 40.07 ** |
| ND | HFD | PFB | LAFB | MAFB | HAFB | |
|---|---|---|---|---|---|---|
| Acetate (μg/g) | 1076.92 ± 381.12 | 510.78 ± 129.98 | 656.79 ± 248.34 | 661.93 ± 416.96 | 660.58 ± 474.07 | 618.82 ± 113.93 |
| Propionate (μg/g) | 687.00 ± 266.87 ** | 292.89 ± 101.46 ## | 184.74 ± 89.85 ## | 105.7 ± 45.75 ## | 223.83 ± 102.67 ## | 255.66 ± 108.68 ## |
| Butyrate (μg/g) | 185.29 ± 25.62 | 171.71 ± 51.1 | 149.61 ± 68.19 | 182.42 ± 107.22 | 207.29 ± 130.05 | 331.67 ± 252.53 |
| Isobutyrate (μg/g) | 164.98 ± 45.65 | 146.31 ± 40.23 | 84.98 ± 1.49 | 81.93 ± 51.63 | 124.35 ± 73.32 | 215.71 ± 93.39 |
| Total SCFA (μg/g) | 1588.55 ± 794.69 | 1171.43 ± 169.26 | 1306.05 ± 575.37 | 976.21 ± 606.54 | 1097.47 ± 732.45 | 1453.19 ± 926.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, S.; Dang, Y.; Xu, H.; Li, A.; Zhou, X.; Gao, X.; Li, Z. Aloe Vera-Fermented Beverage Ameliorates Obesity and Gut Dysbiosis in High-Fat-Diet Mice. Foods 2022, 11, 3728. https://doi.org/10.3390/foods11223728
Fu S, Dang Y, Xu H, Li A, Zhou X, Gao X, Li Z. Aloe Vera-Fermented Beverage Ameliorates Obesity and Gut Dysbiosis in High-Fat-Diet Mice. Foods. 2022; 11(22):3728. https://doi.org/10.3390/foods11223728
Chicago/Turabian StyleFu, Shijie, Yanting Dang, Huilin Xu, Aimin Li, Xiaoman Zhou, Xiaodong Gao, and Zijie Li. 2022. "Aloe Vera-Fermented Beverage Ameliorates Obesity and Gut Dysbiosis in High-Fat-Diet Mice" Foods 11, no. 22: 3728. https://doi.org/10.3390/foods11223728
APA StyleFu, S., Dang, Y., Xu, H., Li, A., Zhou, X., Gao, X., & Li, Z. (2022). Aloe Vera-Fermented Beverage Ameliorates Obesity and Gut Dysbiosis in High-Fat-Diet Mice. Foods, 11(22), 3728. https://doi.org/10.3390/foods11223728