
Gastrointestinal Digestion and Microbial Hydrolysis of Alkyl Gallates: Potential Sustained Release of Gallic Acid

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Animals
2.3. Oral Administration of A-GAs and Sample Preparation
2.3.1. Preparation of Samples from Gastrointestinal Tract Contents
2.3.2. Preparation of Samples from Fecal Bacteria Fermentation
2.3.3. Preparation of Samples from Lactobacillus Fermentation
2.4. HPLC-UV Analysis
2.4.1. HPLC-UV Analytical Method
2.4.2. The External Standard Curves
2.4.3. The Degree of A-GAs Hydrolysis in Fermentation Samples
2.5. Statistical Analysis
3. Results and Discussion
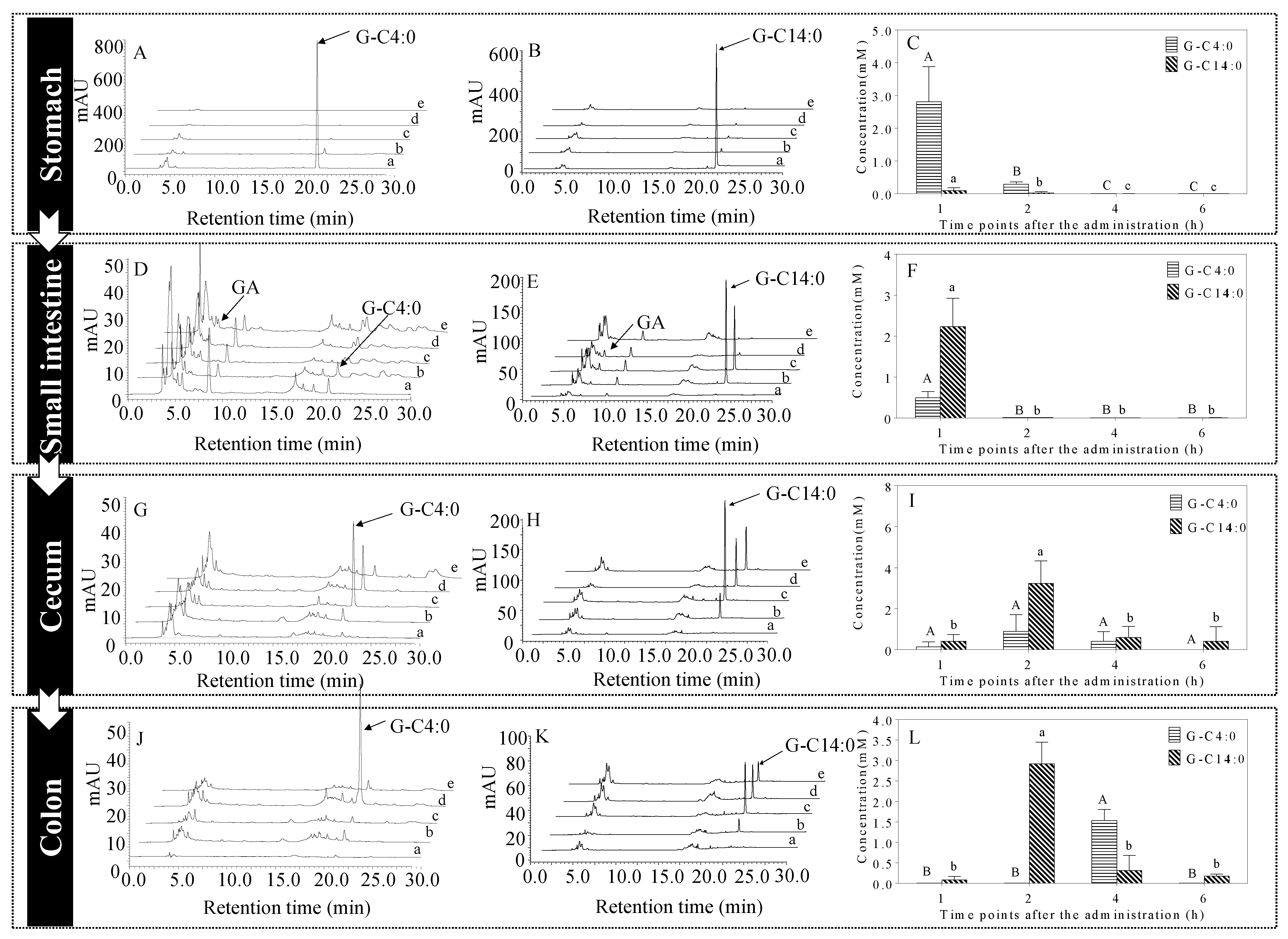
3.1. Concentrations of A-GAs in the Stomach
3.2. Concentrations of A-GAs in the Small Intestine
3.3. Hydrolysis of A-GAs in the Small Intestine
3.4. Concentrations of A-GAs in the Cecum
3.5. Concentrations of A-GAs in the Colon
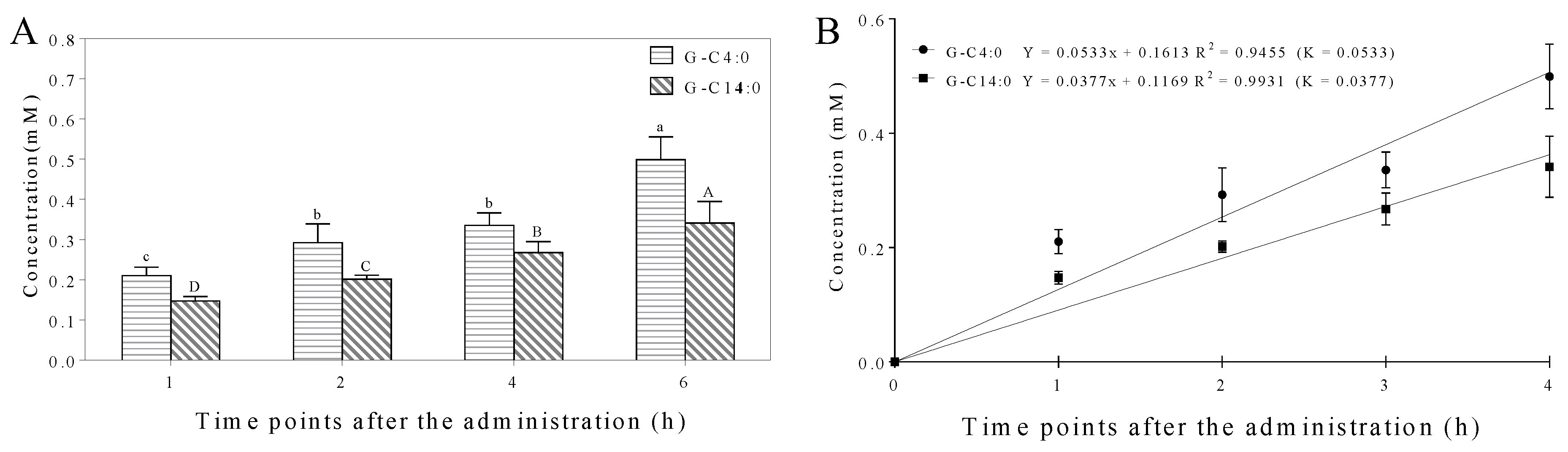
3.6. Microbial Hydrolysis Characteristic of A-GAs by Mice Fecal Microbiota
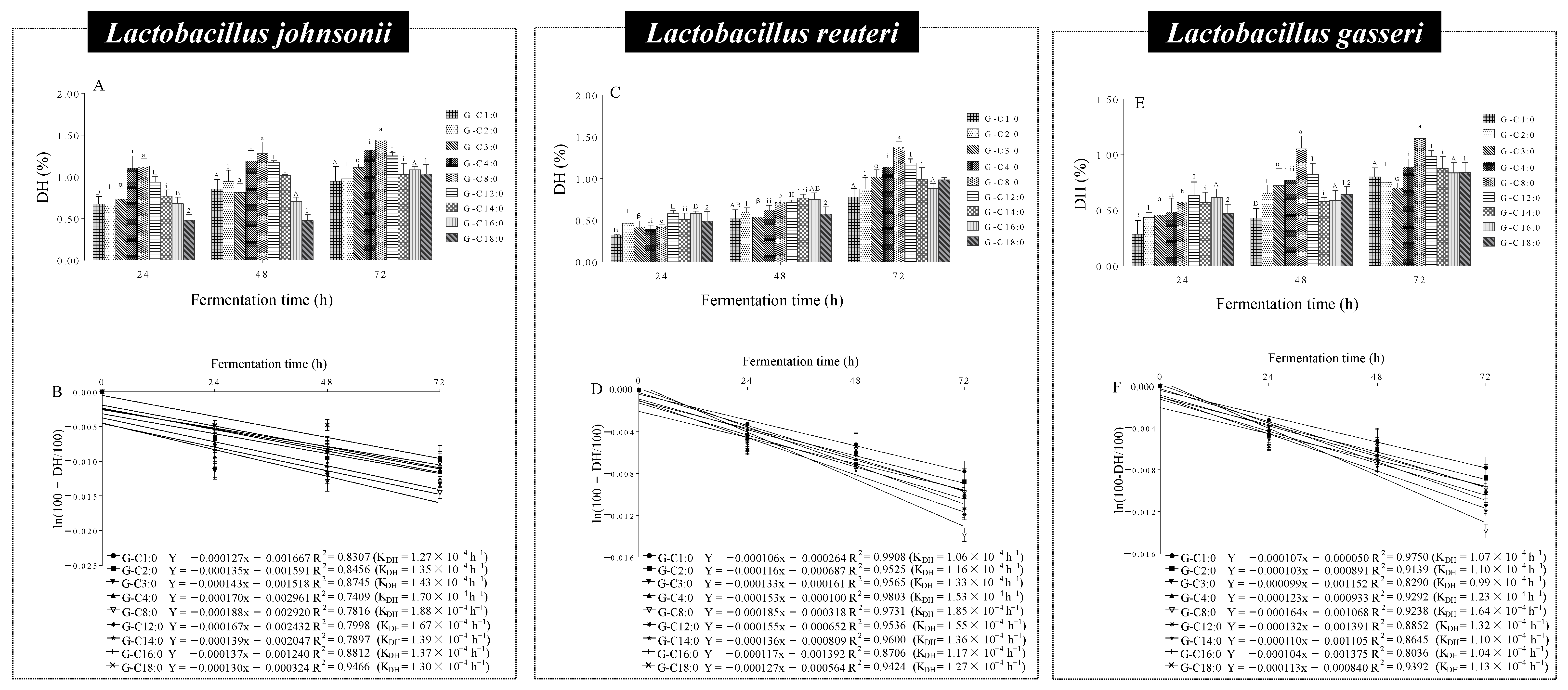
3.7. Microbial Hydrolysis Characteristics of A-GAs by Lactobacillus
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crauste, C.; Rosell, M.; Durand, T.; Vercauteren, J. Omega-3 polyunsaturated lipophenols, how and why? Biochimie 2016, 120, 62–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA Panel on Food Additives; Nutrient Sources Added to Food (ANS). Scientific opinion on the re-evaluation of propyl gallate (E 310) as a food additive. EFSA J. 2014, 12, 3642.
- GB 2760-2014; National Food Safety Standard for Uses of Food Additives. Chinese National Standard, National Health and Family Planning Commission: Beijing, China, 2014.
- Dodo, K.; Minato, T.; Noguchi-Yachide, T.; Suganuma, M.; Hashimoto, Y. Antiproliferative and apoptosis-inducing activities of alkyl gallate and gallamide derivatives related to (-)-epigallocatechin gallate. Bioorg. Med. Chem. 2008, 16, 7975–7982. [Google Scholar] [CrossRef] [PubMed]
- Kubo, I.; Xiao, P.; Fujita, K. Antifungal activity of octyl gallate: Structural criteria and mode of action. Bioorg. Med. Chem. Lett. 2001, 11, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, H.; Akimoto, K.; Shirasaka, N.; Shimizu, S. Inhibitory effects of alkyl gallate and its derivatives on fatty acid desaturation. BBA-Lipid Lipid Met. 1996, 1299, 34–38. [Google Scholar] [CrossRef]
- Kubo, I.; Fujita, K.I.; Nihei, K.I. Anti-salmonella activity of alkyl gallates. J. Agric. Food Chem. 2002, 50, 6692–6696. [Google Scholar] [CrossRef]
- Heijden, C.A.; Janssen, P.J.; Strik, J.J. Toxicology of gallates: A review and evaluation. Food Chem. Toxicol. 1986, 24, 1067–1070. [Google Scholar] [CrossRef]
- Eler, G.J.; Santos, I.S.; De Moraes, A.G.; Mito, M.S.; Comar, J.F.; Peralta, R.M.; Bracht, A. Kinetics of the transformation of n-propyl gallate and structural analogs in the perfused rat liver. Toxicol. Appl. Pharm. 2013, 273, 35–46. [Google Scholar] [CrossRef]
- Gao, S.H.; Zhan, Q.; Li, J.X.; Yang, Q.; Li, X.; Chen, W.S.; Sun, L.N. LC-MS/MS method for the simultaneous determination of ethyl gallate and its major metabolite in rat plasma. Biomed. Chromatogr. 2010, 24, 472–478. [Google Scholar] [CrossRef]
- Seth, H.; Axelsson, M.; Farrell, A.P. The circulation and metabolism of the gastrointestinal tract. Fish Physiol. 2010, 30, 351–393. [Google Scholar]
- Wang, X.M.; Chen, K.F.; Wang, S.J.; Wang, Q.; Hu, Y.Y.; Yin, F.W.; Liu, X.Y.; Zhou, D.Y. Distribution of tyrosol fatty acid esters in the gastrointestinal tracts of mice and their hydrolysis characteristics by gut microbiota. Food Funct. 2022, 13, 2998–3008. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Chen, K.F.; Qiu, J.; Hu, Y.Y.; Yin, F.W.; Liu, X.Y.; Zhou, D.Y. Gastrointestinal distribution of tyrosol acyl esters in orally infected mice and their hydrolysis by Lactobacillus species isolated from the feces of mice. J. Agric. Food Chem. 2022, 70, 1316–1326. [Google Scholar] [CrossRef] [PubMed]
- Carrière, F.; Bezzine, S.; Verger, R. Molecular evolution of the pancreatic lipase and two related enzymes towards different substrate selectivities. J. Mol. Catal. B Enzym. 1997, 3, 55–64. [Google Scholar] [CrossRef]
- Santamarina-Fojo, S.; Brewer, H.B. Lipoprotein lipase: Structure, function and mechanism of action. Int. J. Clin. Lab. Res. 1994, 24, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Dall’Asta, M.; Calani, L.; Tedeschi, M.; Jechiu, L.; Brighenti, F.; Del Rio, D. Identification of microbial metabolites derived from in vitro fecal fermentation of different polyphenolic food sources. Nutrition 2012, 28, 197–203. [Google Scholar] [CrossRef]
- Soybel, D. Anatomy and physiology of the stomach. Surg. Clin. N. Am. 2005, 85, 875–894. [Google Scholar] [CrossRef]
- Rogalska, E.; Ransac, S.; Verger, R. Stereoselectivity of lipases. II. Stereoselective hydrolysis of triglycerides by gastric and pancreatic lipases. J. Biol. Chem. 1990, 265, 20271–20276. [Google Scholar] [CrossRef]
- Tsutomu, N.; Motohiro, Y.; Toshiyuki, S. Pharmacogenetics of intestinal absorption. Curr. Drug Deliv. 2008, 5, 153–169. [Google Scholar]
- Wang, X.M.; Chen, K.F.; Zhang, X.M.; Hu, Y.Y.; Wang, Z.X.; Yin, F.W.; Liu, X.Y.; Zhang, J.H.; Qin, L.; Zhou, D.Y. Effect of carbon chain length on the hydrolysis and transport characteristics of alkyl gallates in rat intestine. Food Funct. 2021, 12, 10581–10588. [Google Scholar] [CrossRef]
- Yin, F.W.; Hu, X.P.; Zhou, D.Y.; Ma, X.C.; Tian, X.G.; Huo, X.K.; Rakariyatham, K.; Shahidi, F.; Zhu, B.W. Evaluation of the stability of tyrosol esters during in vitro gastrointestinal digestion. Food Funct. 2018, 9, 3610–3616. [Google Scholar] [CrossRef]
- Yin, F.W.; Hu, X.P.; Zhou, D.Y.; Ma, X.C.; Tian, X.G.; Huo, X.K.; Rakariyatham, K.; Shahidi, F.; Zhu, B.W. Hydrolysis and transport characteristics of tyrosol acyl esters in rat intestine. J. Agric. Food Chem. 2018, 66, 12521–12526. [Google Scholar] [CrossRef] [PubMed]
- Van Tilbeurgh, H.; Sarda, L.; Verger, R.; Cambillau, C. Structure of the pancreatic lipase-procolipase complex. Nature 1992, 359, 159–162. [Google Scholar] [CrossRef]
- Laguerre, M.; Bily, A.; Roller, M.; Birtić, S. Mass transport phenomena in lipid oxidation and antioxidation. Annu. Rev. Food Sci. Technol. 2017, 8, 391–411. [Google Scholar] [CrossRef]
- Giang, T.M.; Gaucel, S.; Brestaz, P.; Anton, M.; Eynier, A.; Trelea, I.C.; Le Feunteun, S. Dynamic modeling of in vitro lipid digestion: Individual fatty acid release and bio-accessibility kinetics. Food Chem. 2016, 194, 1180–1188. [Google Scholar] [CrossRef]
- Diaz, A.; Muñoz, G.; Caporal-Hernandez, K.; Vázquez-Roque, R.; Lopez-Lopez, G.; Kozina, A.; Espinosa, B.; Flores, G.; Trevino, S.; Guevara, J. Gallic acid improves recognition memory and decreases oxidative-inflammatory damage in the rat hippocampus with metabolic syndrome. Synapse 2020, 75, 1–17. [Google Scholar] [CrossRef] [PubMed]
- El-Ghareb, W.I.; Swidan, M.M.; Ibrahim, I.T.; El-Bary, A.A.; Sakr, T.M. 99mTc-Doxorubicin-loaded gallic acid-gold nanoparticles (99mTc-DOX-loaded GA-Au NPs) as a multifunctional theranostic agent. Int. J. Pharmaceut. 2020, 586, 119514. [Google Scholar] [CrossRef] [PubMed]
- Agatonovic-Kustrin, S.; Kustrin, E.; Morton, D.W. Phenolic acids contribution to antioxidant activities and comparative assessment of phenolic content in mango pulp and peel. S. Afr. J. Bot. 2018, 116, 158–163. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, Z.; Omari-Siaw, E.; Lu, S.; Zhu, Y.; Jiang, N.; Wang, M.; Yu, J.; Xu, X.; Zhang, W. Preparation and in vitro–in vivo evaluation of sustained-release matrix pellets of capsaicin to enhance the oral bioavailability. AAPS PharmSciTech 2015, 17, 339–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Li, H.; Gao, X.; Zhao, Y.; Yu, Z. Pharmacokinetics and absolute bioavailability of gallic acid in rat plasma. J. Shenyang Pharm. Univ. 2008, 25, 944–947. [Google Scholar]
- Hao, Y.W.; Lee, Y.K. Microflora of the gastrointestinal tract: A review. Methods Mol. Biol. 2004, 268, 491–502. [Google Scholar] [PubMed]
- Bailey, M.T.; Coe, C.L. Intestinal microbial patterns of the common marmoset and rhesus macaque. Comp. Biochem. Phys. A 2002, 133, 379–388. [Google Scholar] [CrossRef]
- Fan, M.; Peng, C.; Peng, Y.; Zhang, M.; Li, X. Analysis of metabolites of anthraquinones by human fecal bacteria using UPLC-Q-TOF-HRMS/MS. Chromatographia 2016, 79, 1593–1604. [Google Scholar] [CrossRef]
- López de las Hazas, M.; Piñol, C.; Macià, A.; Romero, M.; Pedret, A.; Solà, R.; Rubió, L.; Motilva, M. Differential absorption and metabolism of hydroxytyrosol and its precursors oleuropein and secoiridoids. J. Funct. Foods. 2016, 22, 52–63. [Google Scholar] [CrossRef] [Green Version]
- Cordle, R.A.; Lowe, M.E. The hydrophobic surface of colipase influences lipase activity at an oil-water interface. J. Lipid Res. 1998, 39, 1759–1767. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, Y.; Kobayashi, I.; Neves, M.A.; Ichikawa, S.; Uemura, K.; Nakajima, M. Formulation of W/O/W emulsions loaded with short-chain fatty acid and their stability improvement by layer-by-layer deposition using dietary fibers. LWT-Food Sci. Technol. 2017, 76, 344–350. [Google Scholar] [CrossRef]
- Niu, Q.; Li, P.; Hao, S.; Kim, S.; Du, T.; Hua, J.; Huang, R. Characteristics of gut microbiota in sows and their relationship with apparent nutrient digestibility. Int. J. Mol. Sci. 2019, 20, 870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guglielmetti, S.; Noni, I.D.; Caracciolo, F.; Molinari, F.; Parini, C.; Mora, D. Bacterial cinnamoyl esterase activity screening for the production of a novel functional food product. Appl. Environ. Microb. 2008, 74, 1284–1288. [Google Scholar] [CrossRef] [Green Version]
- Simons, J.W.F.A.; Adams, H.; Cox, R.C.; Dekker, N.; Götz, F.; Slotboom, A.J.; Verheij, H.M. The lipase from Staphylococcus aureus. Expression in Escherichia coli, large-scale purifieation and comparison of substrate specificity to Staphylococcus hyicus lipase. Nature 2010, 242, 760–769. [Google Scholar]
- Aguilar-Zarate, P.; Cruz, M.A.; Montanez, J.; Rodriguez-Herrera, R.; Wong-Paz, J.E.; Belmares, R.E.; Aguilar, C.N. Gallic acid production under anaerobic submerged fermentation by two bacilli strains. Microb. Cell Fact. 2015, 14, 209. [Google Scholar] [CrossRef] [Green Version]
- Koushik, M.; Prosun, T.; Balaram, M.; Kumar, S.A. Antibacterial activity of long-chain fatty alcohols against mycobacteria. FEMS Microbiol. Lett. 2013, 338, 177–183. [Google Scholar]
- Esteban-Torres, M.; Santamaría, L.; Cabrera-Rubio, R.; Plaza-Vinuesa, L.; Crispie, F.; Rivas, B.D.L.; Cotter, P.; Muñoz, R. A diverse range of human gut bacteria have the potential to metabolize the dietary component gallic acid. Appl. Environ. Microbiol. 2018, 84, e01558-18. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Wang, Q.; Hu, Y.; Yin, F.; Liu, X.; Zhou, D. Gastrointestinal Digestion and Microbial Hydrolysis of Alkyl Gallates: Potential Sustained Release of Gallic Acid. Foods 2022, 11, 3936. https://doi.org/10.3390/foods11233936
Wang X, Wang Q, Hu Y, Yin F, Liu X, Zhou D. Gastrointestinal Digestion and Microbial Hydrolysis of Alkyl Gallates: Potential Sustained Release of Gallic Acid. Foods. 2022; 11(23):3936. https://doi.org/10.3390/foods11233936
Chicago/Turabian StyleWang, Xinmiao, Qian Wang, Yuanyuan Hu, Fawen Yin, Xiaoyang Liu, and Dayong Zhou. 2022. "Gastrointestinal Digestion and Microbial Hydrolysis of Alkyl Gallates: Potential Sustained Release of Gallic Acid" Foods 11, no. 23: 3936. https://doi.org/10.3390/foods11233936
APA StyleWang, X., Wang, Q., Hu, Y., Yin, F., Liu, X., & Zhou, D. (2022). Gastrointestinal Digestion and Microbial Hydrolysis of Alkyl Gallates: Potential Sustained Release of Gallic Acid. Foods, 11(23), 3936. https://doi.org/10.3390/foods11233936