

Theobroma cacao and Theobroma grandiflorum: Botany, Composition and Pharmacological Activities of Pods and Seeds


Abstract
:1. Introduction
2. Botany of Cacao and Cupuassu
2.1. Taxonomy and Origins
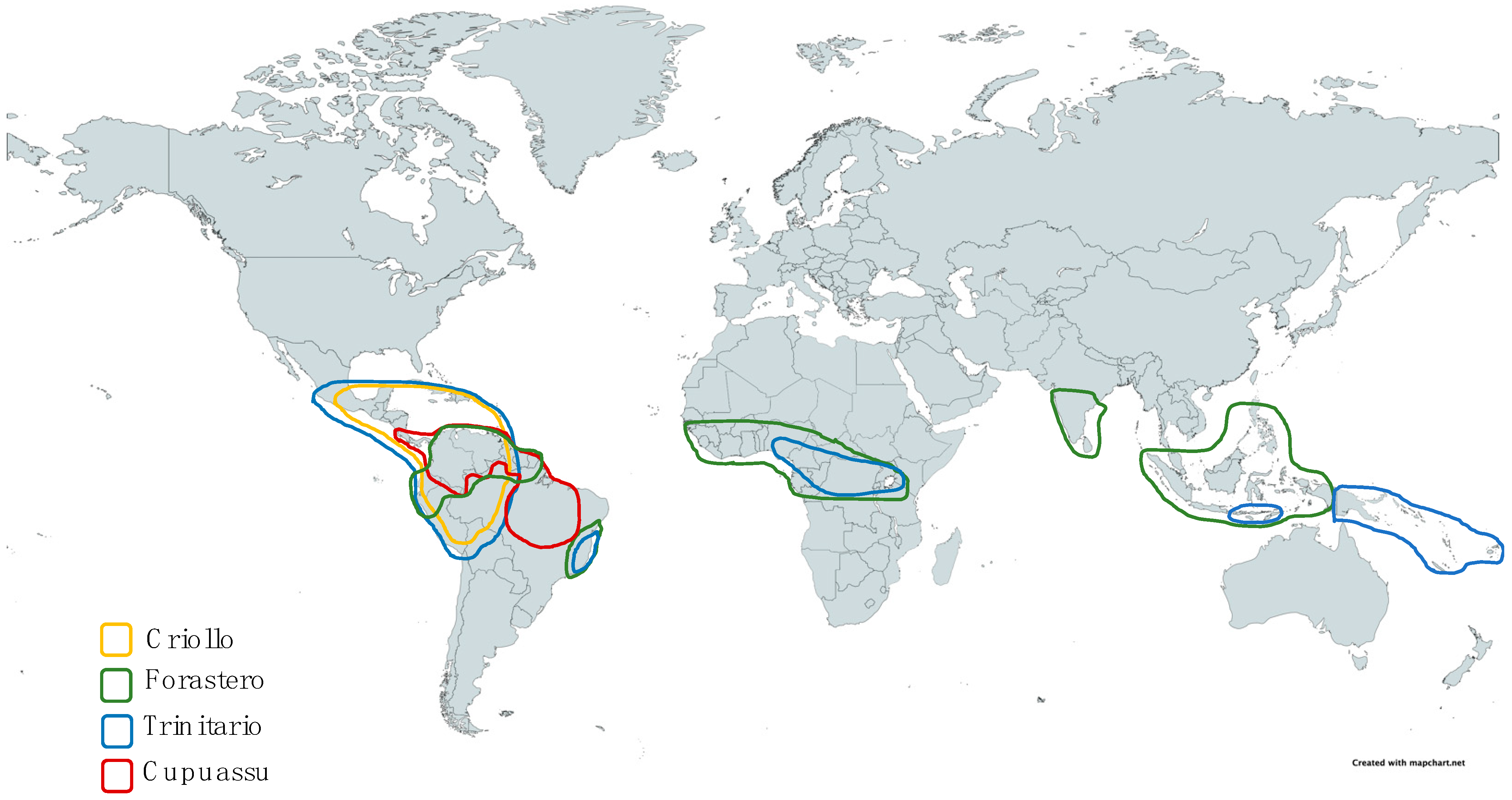
2.2. Agroecology and Geographical Distribution
2.3. Plant Morphologies
3. Chemical Composition of Beans and Pods
3.1. Macronutrients
3.1.1. Proteins
3.1.2. Lipids
3.1.3. Carbohydrates
3.2. Micronutrients
3.2.1. Polyphenols
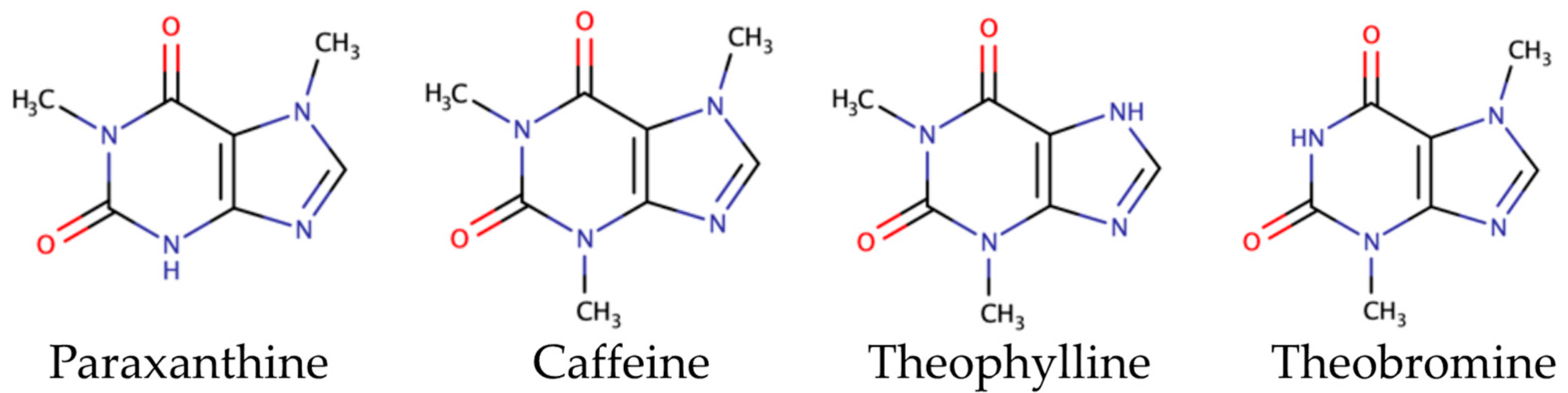
3.2.2. Methylxanthines
4. Pharmacological Activities
4.1. Antioxidant (AO) Activity
4.2. Immunomodulative (IM) Activities
4.3. Impact on Intestinal Tract
4.4. Clinical Trials of Cocoa
5. Bioaccessibility and Bioavailability
6. Perspectives
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lim, T.K. Edible Medicinal and Non Medicinal Plants: Volume 3, Fruits; Springer: Dordrecht, The Netherlands, 2012; ISBN 978-94-007-2534-8. [Google Scholar]
- Cuatrecasas, J. Cacao and Its Allies: A Taxonomic Revision of the Genus Theobroma. In Systematic Plant Studies; Contributions from the United States National Herbarium; Smithsonian Institution: Washington, DC, USA, 1964; Volume 35, ISBN 0097-1618. [Google Scholar]
- Bailey, B.A.; Meinhardt, L.W. Cacao Diseases: A History of Old Enemies and New Encounters; Springer: Berlin/Heidelberg, Germany, 2016; ISBN 978-3-319-24789-2. [Google Scholar]
- Fowler, M.S.; Coutel, F. Cocoa Beans: From Tree to Factory. In Beckett’s Industrial Chocolate Manufacture and Use; Wiley-Blackwell: Hoboken, NJ, USA, 2008; pp. 9–49. ISBN 978-1-118-92359-7. [Google Scholar]
- Toxopeus, H. Botany, Types and Populations. In Cocoa; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2001; pp. 11–37. [Google Scholar]
- Martín, M.A.; Goya, L.; Ramos, S. Preventive Effects of Cocoa and Cocoa Antioxidants in Colon Cancer. Diseases 2016, 4, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potì, F.; Santi, D.; Spaggiari, G.; Zimetti, F.; Zanotti, I. Polyphenol Health Effects on Cardiovascular and Neurodegenerative Disorders: A Review and Meta-Analysis. Int. J. Mol. Sci. 2019, 20, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jean-Marie, E.; Bereau, D.; Robinson, J.-C. Benefits of Polyphenols and Methylxanthines from Cocoa Beans on Dietary Metabolic Disorders. Foods 2021, 10, 2049. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Protiva, P.; Cui, B.; Ma, C.; Baggett, S.; Hequet, V.; Mori, S.; Weinstein, I.B.; Kennelly, E.J. New Bioactive Polyphenols from Theobroma grandiflorum (“cupuaçu”). J. Nat. Prod. 2003, 66, 1501–1504. [Google Scholar] [CrossRef]
- Wood, G.A.R. History and Development. In Cocoa; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2001; pp. 1–10. ISBN 978-0-470-69898-3. [Google Scholar]
- Motamayor, J.C.; Lachenaud, P.; da Silva e Mota, J.W.; Loor, R.; Kuhn, D.N.; Brown, J.S.; Schnell, R.J. Geographic and Genetic Population Differentiation of the Amazonian Chocolate Tree (Theobroma cacao L). PLoS ONE 2008, 3, e3311. [Google Scholar] [CrossRef] [Green Version]
- Clement, C.; Cristo-Araújo, M.; d’Eeckenbrugge, G.; Pereira, A.; Picanço-Rodrigues, D. Origin and Domestication of Native Amazonian Crops. Diversity 2010, 2, 72–106. [Google Scholar] [CrossRef] [Green Version]
- Morris, D. Cacao: How to Grow and How to Cure It; Government Printing Establishment: Washington, DC, USA, 1882. [Google Scholar]
- Hart, J.H. “Cacao”: A Treatise on the Cultivation and Curing of “Cacao” (Theobroma cacao), Botany and Nomenclature of the Same and Hints on the Selection and Management of Estates; U.S. Government Printing Office: Washington, DC, USA, 1892.
- Motamayor, J.C.; Risterucci, A.M.; Heath, M.; Lanaud, C. Cacao Domestication II: Progenitor Germplasm of the Trinitario Cacao Cultivar. Heredity 2003, 91, 322–330. [Google Scholar] [CrossRef] [Green Version]
- Motilal, L.A.; Zhang, D.; Umaharan, P.; Mischke, S.; Mooleedhar, V.; Meinhardt, L.W. The Relic Criollo Cacao in Belize—Genetic Diversity and Relationship with Trinitario and Other Cacao Clones Held in the International Cocoa Genebank, Trinidad. Plant Genet. Resour. 2010, 8, 106–115. [Google Scholar] [CrossRef] [Green Version]
- de Almeida, A.-A.F.; Valle, R.R. Ecophysiology of the Cacao Tree. Braz. J. Plant Physiol. 2007, 19, 425–448. [Google Scholar] [CrossRef] [Green Version]
- Badrie, N.; Bekele, F.; Sikora, E.; Sikora, M. Cocoa Agronomy, Quality, Nutritional, and Health Aspects. Crit. Rev. Food Sci. Nutr. 2015, 55, 620–659. [Google Scholar] [CrossRef]
- Toxopeus, H.; Wessel, M. Studies on Pod and Bean Values of Theobroma cacao L. in Nigeria. I. Environmental Effects on West African Amelonado with Particular Attention to Annual Rainfall Distribution. NJAS 1970, 18, 132–139. [Google Scholar] [CrossRef]
- Soria, J. Notas Sobre Las Principales Variedades de Cacao Cultivadas En America Tropical. In Proceedings of the Conference Internationale sur les Recherches Agronomiques Cacaoyères; Bib. Orton IICA/CATIE: Paris, France, 1967; pp. 247–252. [Google Scholar]
- Alves, R.M.; Sebbenn, A.M.; Artero, A.S.; Clement, C.; Figueira, A. High Levels of Genetic Divergence and Inbreeding in Populations of Cupuassu (Theobroma grandiflorum). Tree Genet. Genomes 2007, 3, 289–298. [Google Scholar] [CrossRef]
- Nair, K.P.P. (Ed.) 5—Cocoa (Theobroma cacao L.). In The Agronomy and Economy of Important Tree Crops of the Developing World; Elsevier: London, UK, 2010; pp. 131–180. ISBN 978-0-12-384677-8. [Google Scholar]
- Dand, R. (Ed.) 2—Agronomics of International Cocoa Production. In The International Cocoa Trade, 3rd ed.; Woodhead Publishing: Sawston, UK, 2011; pp. 23–64. ISBN 978-0-85709-125-3. [Google Scholar]
- Motamayor, J.C.; Risterucci, A.M.; Lopez, P.A.; Ortiz, C.F.; Moreno, A.; Lanaud, C. Cacao Domestication I: The Origin of the Cacao Cultivated by the Mayas. Heredity 2002, 89, 380–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfaia, S.S.; Ribeiro, G.A.; Nobre, A.D.; Luizão, R.C.; Luizão, F.J. Evaluation of Soil Fertility in Smallholder Agroforestry Systems and Pastures in Western Amazonia. Agric. Ecosyst. Environ. 2004, 102, 409–414. [Google Scholar] [CrossRef]
- Schroth, G.; Elias, M.E.A.; Macêdo, J.; D’Angelo, S.A.; Lieberei, R. Growth, Yields and Mineral Nutrition of Cupuaçu (Theobroma grandiflorum) in Two Multi-Strata Agroforestry Systems on a Ferralitic Amazonian Upland Soil at Four Fertilization Levels. J. Appl. Bot. 2001, 75, 74. [Google Scholar]
- van Vliet, J.A.; Slingerland, M.A.; Giller, K.E. Mineral Nutrition of Cocoa: A Review; Wageningen UR: Wageningen, The Netherlands, 2015; ISBN 978-94-6257-705-3. [Google Scholar]
- Lucas, R.E.; Davis, J.F. Relationships between ph values of organic soils and availabilities of 12 plant nutrients. Soil Sci. 1961, 92, 177–182. [Google Scholar] [CrossRef]
- Pontillon, J. Cocoa and Chocolate: Production, Utilization, Characteristics; Tec & Doc- Lavoisier: Cachan, France, 1998; ISBN 2-7430-0174-7. [Google Scholar]
- Lachenaud, P.; Bonnot, F.; Oliver, G. Use of Floral Descriptors to Study Variability in Wild Cocoa Trees (Theobroma cacao L.) in French Guiana. Genet. Resour. Crop Evol. 1999, 46, 491–500. [Google Scholar] [CrossRef]
- Schwan, R.F.; Fleet, G.H. Cocoa and Coffee Fermentations; CRC Press: Boca Raton, FL, USA, 2014; ISBN 978-1-4398-4791-6. [Google Scholar]
- Venturieri, G.A. Flowering Levels, Harvest Season and Yields of Cupuassu (Theobroma grandiflorum). Acta Amaz. 2011, 41, 143–152. [Google Scholar] [CrossRef]
- Ramos, A.R.; Venturieri, G.A.; Cuco, S.M.; Castro, N.M. The Site of Self-Incompatibility Action in Cupuassu (Theobroma grandiflorum). Braz. J. Bot. 2005, 28, 569–578. [Google Scholar] [CrossRef] [Green Version]
- Venturieri, G.A. Floral Biology of Cupuassu (Theobroma grandiflorum (Willdenow Ex Sprengel) Schumann). Ph.D. Thesis, University of Reading, Reading, UK, 1994. [Google Scholar]
- Lanaud, C. Nouvelles Donnees Sur La Biologie Du Cacaoyer (Theobroma cacao L.): Diversite Des Populations, Systeme d’incompatibilite, Haploides Spontanes. Leurs Consequences Pour l’amelioration Genetique de Cette Espece. Ph.D. Thesis, Université de Paris-Sud, Orsay, France, 1987. [Google Scholar]
- Cope, F.W. The Mechanism of Pollen Incompatibility in Theobroma cacao L. Heredity 1962, 17, 157–182. [Google Scholar] [CrossRef] [Green Version]
- Knight, R.; Rogers, H.H. Incompatibility in Theobroma cacao. Heredity 1955, 9, 69–77. [Google Scholar] [CrossRef]
- Lachenaud, P. Les cacaoyers de Guyane. In Science & Découverte; Biotope Editions; BIOTOPE: Villefranche-de-Lauragais, French, 2019; ISBN 2-36662-237-6. [Google Scholar]
- Lachenaud, P.; Sounigo, O.; Sallée, B. Les Cacaoyers Spontanés de Guyane Française: État Des Recherches. Acta Bot. Gall. 2005, 152, 325–346. [Google Scholar] [CrossRef] [Green Version]
- Calzavara, B.B.G.; Müller, C.H.; de Nazaré da Costa Kahwage, O. Fruticultura Tropical o Cupuaçuzeiro; Cultivo, Beneficiamento e Utilização Do Fruto. 1984. Available online: https://www.embrapa.br/busca-de-publicacoes/-/publicacao/387253/fruticultura-tropical-o-cupuacuzeiro-cultivo-beneficiamento-e-utilizacao-do-fruto (accessed on 27 November 2022).
- Sakiroh, S.; Taryono, T.; Purwanti, S. Dynamics of Storage Materials in Cotyledon During Cocoa Seed Germination. Ilmu Pertan. (Agric. Sci.) 2019, 3, 12. [Google Scholar] [CrossRef]
- De Souza, P.; Moreira, L.; Sarmento, D.; Costa, F. Cacao—Theobroma cacao; Academic Press: Cambridge, MA, USA, 2018; pp. 69–76. ISBN 978-0-12-803138-4. Available online: https://www.researchgate.net/publication/323113746_Cacao-Theobroma_cacao (accessed on 27 November 2022).
- Pereira, A.L.F.; Abreu, V.K.G.; Rodrigues, S. Cupuassu—Theobroma grandiflorum. In Exotic Fruits; Rodrigues, S., de Oliveira Silva, E., de Brito, E.S., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 159–162. ISBN 978-0-12-803138-4. [Google Scholar]
- Awang, A.; Mitsui, T. Proteomic Analysis of Theobroma cacao Pod Husk. J. Appl. Glycosci. 2010, 57, 245–264. [Google Scholar] [CrossRef] [Green Version]
- Serra Bonvehí, J.; Ventura Coll, F. Protein Quality Assessment in Cocoa Husk. Food Res. Int. 1999, 32, 201–208. [Google Scholar] [CrossRef]
- Martínez, R.; Torres, P.; Meneses, M.A.; Figueroa, J.G.; Pérez-Álvarez, J.A.; Viuda-Martos, M. Chemical, Technological and in Vitro Antioxidant Properties of Cocoa (Theobroma cacao L.) Co-Products. Food Res. Int. 2012, 49, 39–45. [Google Scholar] [CrossRef]
- Ozung, P.O.; Oko, O.O.K.; Agiang, E.A. Chemical Composition of Differently Treated Forms of Cocoa POD Husk Meal (CPHM). Asian J. Agric. Sci. 2016, 8, 5–9. [Google Scholar] [CrossRef]
- Nguyen, V.T.; Nguyen, N.H. Proximate Composition, Extraction, and Purification of Theobromine from Cacao Pod Husk (Theobroma cacao L.). Technologies 2017, 5, 14. [Google Scholar] [CrossRef]
- Nieto-Figueroa, K.H.; Mendoza-García, N.V.; Gaytán-Martínez, M.; Wall-Medrano, A.; Guadalupe Flavia Loarca-Piña, M.; Campos-Vega, R. Effect of Drying Methods on the Gastrointestinal Fate and Bioactivity of Phytochemicals from Cocoa Pod Husk: In Vitro and in Silico Approaches. Food Res. Int. 2020, 137, 109725. [Google Scholar] [CrossRef]
- Delgado-Ospina, J.; Lucas-González, R.; Viuda-Martos, M.; Fernández-López, J.; Pérez-Álvarez, J.Á.; Martuscelli, M.; Chaves-López, C. Bioactive Compounds and Techno-Functional Properties of High-Fiber Co-Products of the Cacao Agro-Industrial Chain. Heliyon 2021, 7, e06799. [Google Scholar] [CrossRef]
- Sotelo, A.; Alvarez, R.G. Chemical Composition of Wild Theobroma Species and Their Comparison to the Cacao Bean. J. Agric. Food Chem. 1991, 39, 1940–1943. [Google Scholar] [CrossRef]
- Pugliese, A.G.; Tomas-Barberan, F.A.; Truchado, P.; Genovese, M.I. Flavonoids, Proanthocyanidins, Vitamin C, and Antioxidant Activity of Theobroma grandiflorum (Cupuassu) Pulp and Seeds. J. Agric. Food Chem. 2013, 61, 2720–2728. [Google Scholar] [CrossRef]
- Pérez-Mora, W.; Jorrin-Novo, J.V.; Melgarejo, L.M. Substantial Equivalence Analysis in Fruits from Three Theobroma Species through Chemical Composition and Protein Profiling. Food Chem. 2018, 240, 496–504. [Google Scholar] [CrossRef]
- Pugliese, A.G. Compostos Fenólicos do Cupuaçu (Theobroma grandiflorum) e do Cupulate: Composição e Possíveis Benefícios; Universidade de São Paulo: São Paulo, Brazil, 2010. [Google Scholar]
- Servent, A.; Boulanger, R.; Davrieux, F.; Pinot, M.-N.; Tardan, E.; Forestier-Chiron, N.; Hue, C. Assessment of Cocoa (Theobroma cacao L.) Butter Content and Composition throughout Fermentations. Food Res. Int. 2018, 107, 675–682. [Google Scholar] [CrossRef]
- Carvalho, A.; García, N.; Amaya-Farfan, J. Proteins of Cupuacu Seeds (Theobroma grandiflorum Schum) and Changes during Fermentation and Roasting. Food Sci. Technol. 2008, 28, 986–993. [Google Scholar] [CrossRef] [Green Version]
- Martini, M.; Lenci, C.; Figueira, A.; Tavares, D. Localization of the Cotyledon Reserves of Theobroma grandiflorum (Willd. Ex Spreng.) K. Schum., T. Subincanum Mart., T. Bicolor Bonpl. and Their Analogies with T. Cacao L. Braz. J. Bot. 2008, 31, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Pico-Hernández, S.M.; Murillo-Méndez, C.J.; López-Giraldo, L.J. Extraction, Separation, and Evaluation of Antioxidant Effect of the Different Fractions of Polyphenols from Cocoa Beans. Rev. Colomb. De Química 2020, 49, 19–27. [Google Scholar] [CrossRef]
- Adewole, E.; Ajiboye, B.; Oluwafemi, A.O.; Ogunmodede, O. Characterization of cocoa (Theobroma cocoa) pod. Int. J. Sci. Eng. Res. 2013, 4, 1–5. [Google Scholar]
- Lipp, M.; Simoneau, C.; Ulberth, F.; Anklam, E.; Crews, C.; Brereton, P.; Greyt, W.; Schwack, W.; Wiedmaier, C. Composition of Genuine Cocoa Butter and Cocoa Butter Equivalents. J. Food Compos. Anal. 2001, 14, 399–408. [Google Scholar] [CrossRef]
- Asep, E.K.; Jinap, S.; Tan, T.J.; Russly, A.R.; Harcharan, S.; Nazimah, S.A.H. The Effects of Particle Size, Fermentation and Roasting of Cocoa Nibs on Supercritical Fluid Extraction of Cocoa Butter. J. Food Eng. 2008, 85, 450–458. [Google Scholar] [CrossRef]
- Ramos-Escudero, F.; Casimiro-Gonzales, S.; Fernández-Prior, Á.; Cancino Chávez, K.; Gómez-Mendoza, J.; de la Fuente-Carmelino, L.; Muñoz, A.M. Colour, Fatty Acids, Bioactive Compounds, and Total Antioxidant Capacity in Commercial Cocoa Beans (Theobroma cacao L.). LWT 2021, 147, 111629. [Google Scholar] [CrossRef]
- Lannes, S.C.S.; Medeiros, M.L.; Gioielli, L.A. Rheological Properties of Cupuassu and Cocoa Fats. Grasas Y Aceites 2004, 55, 115–121. [Google Scholar] [CrossRef]
- Alviárez, G.E.; Murillo, A.W.; Murillo, P.E.; Rojano, B.A.; Méndez, A.J.J. Characterization and Lipid Extraction of Amazon Cocoa Seeds (Theobroma grandiflorum). Cienc. En Desarro. 2016, 7, 103–109. [Google Scholar]
- Hernández, M.S.; Barrera, J.A.; Carrillo, M. Oferta y Potencialidades de un Banco de Germoplasma del Género Theobroma en el Enriquecimiento de los Sistemas Productivos de la Región Amazónica; SINCHI: Bogotá, Colombia, 2006. [Google Scholar]
- Castro Robayo, Z.R. Caracterización del Proceso de Fermentación del Grano de COPOAZÚ (Theobroma grandiflorum Willd. ex Spreng). Ph.D. Thesis, Universidad Nacional De Colombia, Bogotá, Colombia, 2010. [Google Scholar]
- Megías-Pérez, R.; Grimbs, S.; D’Souza, R.N.; Bernaert, H.; Kuhnert, N. Profiling, Quantification and Classification of Cocoa Beans Based on Chemometric Analysis of Carbohydrates Using Hydrophilic Interaction Liquid Chromatography Coupled to Mass Spectrometry. Food Chem. 2018, 258, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Coco, F.L.; Lanuzza, F.; Micali, G.; Cappellano, G. Determination of Theobromine, Theophylline, and Caffeine in by-Products of Cupuacu and Cacao Seeds by High-Performance Liquid Chromatography. J. Chromatogr. Sci. 2007, 45, 273–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul Karim, A.; Azlan, A.; Ismail, A.; Hashim, P.; Abd Gani, S.S.; Zainudin, B.H.; Abdullah, N.A. Phenolic Composition, Antioxidant, Anti-Wrinkles and Tyrosinase Inhibitory Activities of Cocoa Pod Extract. BMC Complement. Altern. Med. 2014, 14, 381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irondi, A.E.; Olawuyi, A.D.; Lawal, B.S.; Boligon, A.A.; Olasupo, F.; Sobowale, O. Comparative Inhibitory Effects of Cocoa Bean and Cocoa Pod Husk Extracts on Enzymes Associated with Hyperuricemia and Hypertension in Vitro. Int. Food Res. J. 2019, 26, 557–564. [Google Scholar]
- Wollgast, J.; Anklam, E. Review on Polyphenols in Theobroma cacao: Changes in Composition during the Manufacture of Chocolate and Methodology for Identification and Quantification. Food Res. Int. 2000, 33, 423–447. [Google Scholar] [CrossRef]
- Niemenak, N.; Rohsius, C.; Elwers, S.; Omokolo Ndoumou, D.; Lieberei, R. Comparative Study of Different Cocoa (Theobroma cacao L.) Clones in Terms of Their Phenolics and Anthocyanins Contents. J. Food Compos. Anal. 2006, 19, 612–619. [Google Scholar] [CrossRef]
- Ortega, N.; Romero, M.P.; Macià, A.; Reguant, J.; Anglès, N.; Morelló, J.R.; Motilva, M.J. Obtention and Characterization of Phenolic Extracts from Different Cocoa Sources. J. Agric. Food Chem. 2008, 56, 9621–9627. [Google Scholar] [CrossRef]
- Rusconi, M.; Conti, A. Theobroma cacao L., the Food of the Gods: A Scientific Approach beyond Myths and Claims. Pharmacol. Res. 2010, 61, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Jean-Marie, E.; Bereau, D.; Poucheret, P.; Guzman, C.; Boudard, F.; Robinson, J.-C. Antioxidative and Immunomodulatory Potential of the Endemic French Guiana Wild Cocoa “Guiana”. Foods 2021, 10, 522. [Google Scholar] [CrossRef] [PubMed]
- de Barros, H.R.M.; García-Villalba, R.; Tomás-Barberán, F.A.; Genovese, M.I. Evaluation of the Distribution and Metabolism of Polyphenols Derived from Cupuassu (Theobroma grandiflorum) in Mice Gastrointestinal Tract by UPLC-ESI-QTOF. J. Funct. Foods 2016, 22, 477–489. [Google Scholar] [CrossRef]
- Valadez-Carmona, L.; Plazola-Jacinto, C.P.; Hernández-Ortega, M.; Hernández-Navarro, M.D.; Villarreal, F.; Necoechea-Mondragón, H.; Ortiz-Moreno, A.; Ceballos-Reyes, G. Effects of Microwaves, Hot Air and Freeze-Drying on the Phenolic Compounds, Antioxidant Capacity, Enzyme Activity and Microstructure of Cacao Pod Husks (Theobroma cacao L.). Innov. Food Sci. Emerg. Technol. 2017, 41, 378–386. [Google Scholar] [CrossRef]
- Natsir, S.; Djide, M.N.; Alam, G. Effect of Cocoa Pod Husk Extracts on Prevention of Hypercholesterolemia in Albino Rats (Rattus Novergicus). Farm. Dan Farmakol. 2008, 12, 78–82. [Google Scholar]
- Batista, N.N.; de Andrade, D.P.; Ramos, C.L.; Dias, D.R.; Schwan, R.F. Antioxidant Capacity of Cocoa Beans and Chocolate Assessed by FTIR. Food Res. Int. 2016, 90, 313–319. [Google Scholar] [CrossRef]
- Acosta-Otálvaro, E.; Valencia-Gallego, W.; Mazo-Rivas, J.C.; García-Viguera, C. Cocoa Extract with High Content of Flavan 3-Ols, Procyanidins and Methylxanthines. J. Food Sci. Technol. 2022, 59, 1152–1161. [Google Scholar] [CrossRef]
- Elwers, S.; Zambrano, A.; Rohsius, C.; Lieberei, R. Differences between the Content of Phenolic Compounds in Criollo, Forastero and Trinitario Cocoa Seed (Theobroma cacao L.). Eur. Food Res. Technol. 2009, 229, 937–948. [Google Scholar] [CrossRef]
- Oracz, J.; Zyzelewicz, D.; Nebesny, E. The Content of Polyphenolic Compounds in Cocoa Beans (Theobroma cacao L.), Depending on Variety, Growing Region, and Processing Operations: A Review. Crit. Rev. Food Sci. Nutr. 2013, 55, 1176–1192. [Google Scholar] [CrossRef]
- Dang, Y.K.T.; Nguyen, H.V.H. Effects of Maturity at Harvest and Fermentation Conditions on Bioactive Compounds of Cocoa Beans. Plant Foods Hum. Nutr. 2019, 74, 54–60. [Google Scholar] [CrossRef]
- Urbańska, B.; Kowalska, J.; Lacerda Ramos, C. Comparison of the Total Polyphenol Content and Antioxidant Activity of Chocolate Obtained from Roasted and Unroasted Cocoa Beans from Different Regions of the World. Antioxidants 2019, 8, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Oliveira, T.B.; Genovese, M.I. Chemical Composition of Cupuassu (Theobroma grandiflorum) and Cocoa (Theobroma cacao) Liquors and Their Effects on Streptozotocin-Induced Diabetic Rats. Food Res. Int. 2013, 51, 929–935. [Google Scholar] [CrossRef] [Green Version]
- Ashihara, H.; Kato, M.; Crozier, A. Distribution, Biosynthesis and Catabolism of Methylxanthines in Plants. In Methylxanthines; Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2011; Volume 200, pp. 11–31. ISBN 978-3-642-13442-5. [Google Scholar]
- Grases, F.; Rodriguez, A.; Costa-Bauza, A. Theobromine Inhibits Uric Acid Crystallization. A Potential Application in the Treatment of Uric Acid Nephrolithiasis. PLoS ONE 2014, 9, e111184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sotelo, C.L.; Alvis, B.A.; Arrázola, P.G. Evaluation of Epicatechin, Theobromine and Caffeine in Cacao Husks (Theobroma cacao, L.), Determination of the Antioxidant Capacity. Rev. Colomb. De Cienc. Hortícolas 2015, 9, 124–134. [Google Scholar] [CrossRef]
- Peláez, P.P.; Bardón, I.; Camasca, P. Methylxanthine and Catechin Content of Fresh and Fermented Cocoa Beans, Dried Cocoa Beans, and Cocoa Liquor. Sci. Agropecu. 2016, 7, 355–365. [Google Scholar] [CrossRef]
- Hernández-Hernández, C.; Viera-Alcaide, I.; Morales-Sillero, A.M.; Fernández-Bolaños, J.; Rodríguez-Gutiérrez, G. Bioactive Compounds in Mexican Genotypes of Cocoa Cotyledon and Husk. Food Chem. 2018, 240, 831–839. [Google Scholar] [CrossRef] [Green Version]
- Campos-Vega, R.; Nieto-Figueroa, K.H.; Oomah, B.D. Cocoa (Theobroma cacao L.) Pod Husk: Renewable Source of Bioactive Compounds. Trends Food Sci. Technol. 2018, 81, 172–184. [Google Scholar] [CrossRef]
- Oracz, J.; Nebesny, E. Antioxidant Properties of Cocoa Beans (Theobroma cacao L.): Influence of Cultivar and Roasting Conditions. Int. J. Food Prop. 2016, 19, 1242–1258. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.W.; Kim, Y.J.; Lee, H.J.; Lee, C.Y. Cocoa Has More Phenolic Phytochemicals and a Higher Antioxidant Capacity than Teas and Red Wine. J. Agric. Food Chem. 2003, 51, 7292–7295. [Google Scholar] [CrossRef]
- Cuellar-Álvarez, L.; Cuellar-Álvarez, N.; Galeano-García, P.; Suárez Salazar, J.C. Effect of Fermentation Time on Phenolic Content and Antioxidant Potential in Cupuassu (Theobroma grandiflorum (Willd. Ex Spreng.) K.Schum.) Beans. Acta Agronómica 2017, 66, 473–479. [Google Scholar] [CrossRef] [Green Version]
- Carrillo, L.C.; Londoño-Londoño, J.; Gil, A. Comparison of Polyphenol, Methylxanthines and Antioxidant Activity in Theobroma cacao Beans from Different Cocoa-Growing Areas in Colombia. Food Res. Int. 2014, 60, 273–280. [Google Scholar] [CrossRef]
- Gültekin-Özgüven, M.; Berktaş, I.; Özçelik, B. Change in Stability of Procyanidins, Antioxidant Capacity and in-Vitro Bioaccessibility during Processing of Cocoa Powder from Cocoa Beans. LWT Food Sci. Technol. 2016, 72, 559–565. [Google Scholar] [CrossRef]
- Martín, M.A.; Ramos, S.; Mateos, R.; Izquierdo-Pulido, M.; Bravo, L.; Goya, L. Protection of Human HepG2 Cells against Oxidative Stress by the Flavonoid Epicatechin. Phytother. Res. PTR 2010, 24, 503–509. [Google Scholar] [CrossRef]
- Martins, T.F.; Palomino, O.M.; Álvarez-Cilleros, D.; Martín, M.A.; Ramos, S.; Goya, L. Cocoa Flavanols Protect Human Endothelial Cells from Oxidative Stress. Plant Foods Hum. Nutr. 2020, 75, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Pinent, M.; Castell-Auví, A.; Genovese, M.I.; Serrano, J.; Casanova, A.; Blay, M.; Ardévol, A. Antioxidant Effects of Proanthocyanidin-Rich Natural Extracts from Grape Seed and Cupuassu on Gastrointestinal Mucosa. J. Sci. Food Agric. 2016, 96, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Figueira, A.; Janick, J.; BeMiller, J.N. New Products from Theobroma cacao: Seed Pulp and Pod Gum. New Crops 1993, 475, 478. [Google Scholar]
- Carta, S.; Tsiplakou, E.; Mitsiopoulou, C.; Pulina, G.; Nudda, A. Cocoa Husks Fed to Lactating Dairy Ewes Affect Milk Fatty Acid Profile and Oxidative Status of Blood and Milk. Small Rumin. Res. 2022, 207, 106599. [Google Scholar] [CrossRef]
- López-Yerena, A.; Perez, M.; Vallverdú-Queralt, A.; Escribano-Ferrer, E. Insights into the Binding of Dietary Phenolic Compounds to Human Serum Albumin and Food-Drug Interactions. Pharmaceutics 2020, 12, 1123. [Google Scholar] [CrossRef]
- Cucaita, N.A.; Hernández, M.; Gutiérrez, R.H. Comparison between Chocolate and an Analog Product Made from Copoazú (Theobroma grandiflorum). Acta Hortic. 2014, 1047, 231–236. [Google Scholar] [CrossRef]
- Camu, N.; Winter, T.D.; Addo, S.K.; Takrama, J.S.; Bernaert, H.; Vuyst, L.D. Fermentation of Cocoa Beans: Influence of Microbial Activities and Polyphenol Concentrations on the Flavour of Chocolate. J. Sci. Food Agric. 2008, 88, 2288–2297. [Google Scholar] [CrossRef]
- Santander Muñoz, M.; Rodríguez Cortina, J.; Vaillant, F.E.; Escobar Parra, S. An Overview of the Physical and Biochemical Transformation of Cocoa Seeds to Beans and to Chocolate: Flavor Formation. Crit. Rev. Food Sci. Nutr. 2020, 60, 1593–1613. [Google Scholar] [CrossRef] [PubMed]
- Suazo, Y.; Davidov-Pardo, G.; Arozarena, I. Effect of Fermentation and Roasting on the Phenolic Concentration and Antioxidant Activity of Cocoa from Nicaragua. J. Food Qual. 2014, 37, 50–56. [Google Scholar] [CrossRef]
- Bordiga, M.; Locatelli, M.; Travaglia, F.; Coïsson, J.D.; Mazza, G.; Arlorio, M. Evaluation of the Effect of Processing on Cocoa Polyphenols: Antiradical Activity, Anthocyanins and Procyanidins Profiling from Raw Beans to Chocolate. Int. J. Food Sci. Technol. 2015, 50, 840–848. [Google Scholar] [CrossRef]
- Genovese, M.I.; Lannes, S.C. da S. Comparison of Total Phenolic Content and Antiradical Capacity of Powders and “Chocolates” from Cocoa and Cupuassu. Food Sci. Technol. 2009, 29, 810–814. [Google Scholar] [CrossRef] [Green Version]
- Todorovic, V.; Redovnikovic, I.R.; Todorovic, Z.; Jankovic, G.; Dodevska, M.; Sobajic, S. Polyphenols, Methylxanthines, and Antioxidant Capacity of Chocolates Produced in Serbia. J. Food Compos. Anal. 2015, 41, 137–143. [Google Scholar] [CrossRef]
- Bagri, P.; Ali, M.; Aeri, V.; Bhowmik, M.; Sultana, S. Antidiabetic Effect of Punica Granatum Flowers: Effect on Hyperlipidemia, Pancreatic Cells Lipid Peroxidation and Antioxidant Enzymes in Experimental Diabetes. Food Chem. Toxicol. 2009, 47, 50–54. [Google Scholar] [CrossRef]
- de Oliveira, T.B.; Rogero, M.M.; Genovese, M.I. Poliphenolic-Rich Extracts from Cocoa (Theobroma cacao L.) and Cupuassu (Theobroma grandiflorum Willd. Ex Spreng. K. Shum) Liquors: A Comparison of Metabolic Effects in High-Fat Fed Rats. PharmaNutrition 2015, 3, 20–28. [Google Scholar] [CrossRef]
- Angulo, P. Nonalcoholic Fatty Liver Disease. N. Engl. J. Med. 2002, 346, 1221–1231. [Google Scholar] [CrossRef] [Green Version]
- Abbas, A.K.; Lichtman, A.H. Les Bases de L’immunologie Fondamentale et Clinique; Elsevier Masson: Paris, France, 2008; ISBN 978-2-8101-0023-1. [Google Scholar]
- Clos, J. Immunité Chez les Animaux et les Végétaux; Lavoisier: Paris, France, 2012; ISBN 978-2-257-70522-8. [Google Scholar]
- Pasquier, C. Stress oxydatif et inflammation. Rev. Française Des Lab. 1995, 1995, 87–92. [Google Scholar] [CrossRef]
- Kurutas, E.B. The Importance of Antioxidants Which Play the Role in Cellular Response against Oxidative/Nitrosative Stress: Current State. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Liang, H.; Zen, K. Molecular Mechanisms That Influence the Macrophage M1-M2 Polarization Balance. Front. Immunol. 2014, 5, 614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, M.J.; Liu, Z. Crosstalk of Reactive Oxygen Species and NF-ΚB Signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosser, D.M. The Many Faces of Macrophage Activation. J. Leukoc. Biol. 2003, 73, 209–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dugo, L.; Belluomo, M.G.; Fanali, C.; Russo, M.; Cacciola, F.; Maccarrone, M.; Sardanelli, A.M. Effect of Cocoa Polyphenolic Extract on Macrophage Polarization from Proinflammatory M1 to Anti-Inflammatory M2 State. Oxidative Med. Cell Longev. 2017, 2017, 6293740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Punaro, G.R.; Lima, D.Y.; Rodrigues, A.M.; Pugliero, S.; Mouro, M.G.; Rogero, M.M.; Higa, E.M.S. Cupuaçu Extract Reduces Nitrosative Stress and Modulates Inflammatory Mediators in the Kidneys of Experimental Diabetes. Clin. Nutr. 2019, 38, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Curimbaba, T.F.S.; Almeida-Junior, L.D.; Chagas, A.S.; Quaglio, A.E.V.; Herculano, A.M.; Di Stasi, L.C. Prebiotic, Antioxidant and Anti-Inflammatory Properties of Edible Amazon Fruits. Food Biosci. 2020, 36, 100599. [Google Scholar] [CrossRef]
- Ramiro, E.; Franch, À.; Castellote, C.; Pérez-Cano, F.; Permanyer, J.; Izquierdo-Pulido, M.; Castell, M. Flavonoids from Theobroma cacao Down-Regulate Inflammatory Mediators. J. Agric. Food Chem. 2005, 53, 8506–8511. [Google Scholar] [CrossRef]
- Mao, T.K.; van de Water, J.; Keen, C.L.; Schmitz, H.H.; Gershwin, M.E. Modulation of TNF-α Secretion in Peripheral Blood Mononuclear Cells by Cocoa Flavanols and Procyanidins. J. Immunol. Res. 2002, 9, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Berezo, T.; Ramiro-Puig, E.; Pérez-Cano, F.J.; Castellote, C.; Permanyer, J.; Franch, A.; Castell, M. Influence of a Cocoa-Enriched Diet on Specific Immune Response in Ovalbumin-Sensitized Rats. Mol. Nutr. Food Res. 2009, 53, 389–397. [Google Scholar] [CrossRef]
- Abril-Gil, M.; Pérez-Cano, F.J.; Franch, À.; Castell, M. Effect of a Cocoa-Enriched Diet on Immune Response and Anaphylaxis in a Food Allergy Model in Brown Norway Rats. J. Nutr. Biochem. 2016, 27, 317–326. [Google Scholar] [CrossRef]
- Rossin, D.; Barbosa-Pereira, L.; Iaia, N.; Sottero, B.; Danzero, A.C.; Poli, G.; Zeppa, G.; Biasi, F. Protective Effect of Cocoa Bean Shell against Intestinal Damage: An Example of Byproduct Valorization. Antioxidants 2021, 10, 280. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Cilleros, D.; Ramos, S.; López-Oliva, M.E.; Escrivá, F.; Álvarez, C.; Fernández-Millán, E.; Martín, M.Á. Cocoa Diet Modulates Gut Microbiota Composition and Improves Intestinal Health in Zucker Diabetic Rats. Food Res. Int. 2020, 132, 109058. [Google Scholar] [CrossRef] [PubMed]
- Andrade, J.K.S.; Barros, R.G.C.; Pereira, U.C.; Gualberto, N.C.; de Oliveira, C.S.; Shanmugam, S.; Narain, N. α-Amylase Inhibition, Cytotoxicity and Influence of the in Vitro Gastrointestinal Digestion on the Bioaccessibility of Phenolic Compounds in the Peel and Seed of Theobroma grandiflorum. Food Chem. 2022, 373, 131494. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Ademosun, A.O.; Ademiluyi, A.O.; Omojokun, O.S.; Nwanna, E.E.; Longe, K.O. In Vitro Studies on the Antioxidant Property and Inhibition of α-Amylase, α-Glucosidase, and Angiotensin I-Converting Enzyme by Polyphenol-Rich Extracts from Cocoa (Theobroma cacao) Bean. Pathol. Res. Int. 2014, 2014, e549287. [Google Scholar] [CrossRef]
- Ryan, C.M.; Khoo, W.; Ye, L.; Lambert, J.D.; O’Keefe, S.F.; Neilson, A.P. Loss of Native Flavanols during Fermentation and Roasting Does Not Necessarily Reduce Digestive Enzyme-Inhibiting Bioactivities of Cocoa. J. Agric. Food Chem. 2016, 64, 3616–3625. [Google Scholar] [CrossRef] [PubMed]
- Cantele, C.; Rojo-Poveda, O.; Bertolino, M.; Ghirardello, D.; Cardenia, V.; Barbosa-Pereira, L.; Zeppa, G. In Vitro Bioaccessibility and Functional Properties of Phenolic Compounds from Enriched Beverages Based on Cocoa Bean Shell. Foods 2020, 9, 715. [Google Scholar] [CrossRef] [PubMed]
- Tabone, M.; García-Merino, J.A.; Bressa, C.; Rocha Guzman, N.E.; Herrera Rocha, K.; Chu Van, E.; Castelli, F.A.; Fenaille, F.; Larrosa, M. Chronic Consumption of Cocoa Rich in Procyanidins Has a Marginal Impact on Gut Microbiota and on Serum and Fecal Metabolomes in Male Endurance Athletes. J. Agric. Food Chem. 2022, 70, 1878–1889. [Google Scholar] [CrossRef]
- Shin, J.-H.; Kim, C.-S.; Cha, L.; Kim, S.; Lee, S.; Chae, S.; Chun, W.Y.; Shin, D.-M. Consumption of 85% Cocoa Dark Chocolate Improves Mood in Association with Gut Microbial Changes in Healthy Adults: A Randomized Controlled Trial. J. Nutr. Biochem. 2022, 99, 108854. [Google Scholar] [CrossRef]
- Rodríguez-Lagunas, M.J.; Vicente, F.; Pereira, P.; Castell, M.; Pérez-Cano, F.J. Relationship between Cocoa Intake and Healthy Status: A Pilot Study in University Students. Molecules 2019, 24, 812. [Google Scholar] [CrossRef] [Green Version]
- Urpi-Sarda, M.; Ramiro-Puig, E.; Khan, N.; Ramos-Romero, S.; Llorach, R.; Castell, M.; Gonzalez-Manzano, S.; Santos-Buelga, C.; Andres-Lacueva, C. Distribution of Epicatechin Metabolites in Lymphoid Tissues and Testes of Young Rats with a Cocoa-Enriched Diet. Br. J. Nutr. 2010, 103, 1393–1397. [Google Scholar] [CrossRef] [Green Version]
- Ramiro-Puig, E.; Urpí-Sardà, M.; Pérez-Cano, F.J.; Franch, À.; Castellote, C.; Andrés-Lacueva, C.; Izquierdo-Pulido, M.; Castell, M. Cocoa-Enriched Diet Enhances Antioxidant Enzyme Activity and Modulates Lymphocyte Composition in Thymus from Young Rats. J. Agric. Food Chem. 2007, 55, 6431–6438. [Google Scholar] [CrossRef] [PubMed]
- Barrera-Reyes, P.K.; Hernández-Ramírez, N.; Cortés, J.; Poquet, L.; Redeuil, K.; Rangel-Escareño, C.; Kussmann, M.; Silva-Zolezzi, I.; Tejero, M.E. Gene Expression Changes by High-Polyphenols Cocoa Powder Intake: A Randomized Crossover Clinical Study. Eur. J. Nutr. 2019, 58, 1887–1898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munguia, L.; Rubio-Gayosso, I.; Ramirez-Sanchez, I.; Ortiz, A.; Hidalgo, I.; Gonzalez, C.; Meaney, E.; Villarreal, F.; Najera, N.; Ceballos, G. High Flavonoid Cocoa Supplement Ameliorates Plasma Oxidative Stress and Inflammation Levels While Improving Mobility and Quality of Life in Older Subjects: A Double-Blind Randomized Clinical Trial. J. Gerontol. Ser. A 2019, 74, 1620–1627. [Google Scholar] [CrossRef] [PubMed]
- Saura-Calixto, F.; Serrano, J.; Goñi, I. Intake and Bioaccessibility of Total Polyphenols in a Whole Diet. Food Chem. 2007, 101, 492–501. [Google Scholar] [CrossRef] [Green Version]
- Kamiloglu, S.; Tomas, M.; Ozdal, T.; Capanoglu, E. Effect of Food Matrix on the Content and Bioavailability of Flavonoids. Trends Food Sci. Technol. 2021, 117, 15–33. [Google Scholar] [CrossRef]
- Neilson, A.P.; Ferruzzi, M.G. Influence of Formulation and Processing on Absorption and Metabolism of Flavan-3-Ols from Tea and Cocoa. Annu. Rev. Food Sci. Technol. 2011, 2, 125–151. [Google Scholar] [CrossRef]
- Rein, M.J.; Renouf, M.; Cruz-Hernandez, C.; Actis-Goretta, L.; Thakkar, S.K.; da Silva Pinto, M. Bioavailability of Bioactive Food Compounds: A Challenging Journey to Bioefficacy. Br. J. Clin. Pharmacol. 2013, 75, 588–602. [Google Scholar] [CrossRef] [Green Version]
- Sorrenti, V.; Fortinguerra, S.; Caudullo, G.; Buriani, A. Deciphering the Role of Polyphenols in Sports Performance: From Nutritional Genomics to the Gut Microbiota toward Phytonutritional Epigenomics. Nutrients 2020, 12, 1265. [Google Scholar] [CrossRef]
- Sorrenti, V.; Ali, S.; Mancin, L.; Davinelli, S.; Paoli, A.; Scapagnini, G. Cocoa Polyphenols and Gut Microbiota Interplay: Bioavailability, Prebiotic Effect, and Impact on Human Health. Nutrients 2020, 12, 1908. [Google Scholar] [CrossRef]
- Baba, S.; Osakabe, N.; Yasuda, A.; Natsume, M.; Takizawa, T.; Nakamura, T.; Terao, J. Bioavailability of (–)-Epicatechin upon Intake of Chocolate and Cocoa in Human Volunteers. Free. Radic. Res. 2000, 33, 635–641. [Google Scholar] [CrossRef]
- Kuhnle, G.; Spencer, J.P.; Schroeter, H.; Shenoy, B.; Debnam, E.S.; Srai, S.K.; Rice-Evans, C.; Hahn, U. Epicatechin and Catechin Are O-Methylated and Glucuronidated in the Small Intestine. Biochem. Biophys. Res. Commun. 2000, 277, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Roura, E.; Andrés-Lacueva, C.; Jáuregui, O.; Badia, E.; Estruch, R.; Izquierdo-Pulido, M.; Lamuela-Raventós, R.M. Rapid Liquid Chromatography Tandem Mass Spectrometry Assay to Quantify Plasma (–)-Epicatechin Metabolites after Ingestion of a Standard Portion of Cocoa Beverage in Humans. J. Agric. Food Chem. 2005, 53, 6190–6194. [Google Scholar] [CrossRef] [PubMed]
- D’Archivio, M.; Filesi, C.; Di Benedetto, R.; Gargiulo, R.; Giovannini, C.; Masella, R. Polyphenols, Dietary Sources and Bioavailability. Ann. Dell’istituto Super. Di Sanita 2007, 43, 348–361. [Google Scholar]
- Montagnana, M.; Danese, E.; Angelino, D.; Mena, P.; Rosi, A.; Benati, M.; Gelati, M.; Salvagno, G.L.; Favaloro, E.J.; Del Rio, D.; et al. Dark Chocolate Modulates Platelet Function with a Mechanism Mediated by Flavan-3-Ol Metabolites. Medicine 2018, 97, e13432. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and Bioefficacy of Polyphenols in Humans. I. Review of 97 Bioavailability Studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [Green Version]
- Tsang, C.; Auger, C.; Mullen, W.; Bornet, A.; Rouanet, J.-M.; Crozier, A.; Teissedre, P.-L. The Absorption, Metabolism and Excretion of Flavan-3-Ols and Procyanidins Following the Ingestion of a Grape Seed Extract by Rats. Br. J. Nutr. 2005, 94, 170–181. [Google Scholar] [CrossRef]
- Serra, A.; Macià, A.; Romero, M.-P.; Valls, J.; Bladé, C.; Arola, L.; Motilva, M.-J. Bioavailability of Procyanidin Dimers and Trimers and Matrix Food Effects in in Vitro and in Vivo Models. Br. J. Nutr. 2010, 103, 944–952. [Google Scholar] [CrossRef]
- Déprez, S.; Brezillon, C.; Rabot, S.; Philippe, C.; Mila, I.; Lapierre, C.; Scalbert, A. Polymeric Proanthocyanidins Are Catabolized by Human Colonic Microflora into Low-Molecular-Weight Phenolic Acids. J. Nutr. 2000, 130, 2733–2738. [Google Scholar] [CrossRef]
- Dai, P.; Zhu, L.; Luo, F.; Lu, L.; Li, Q.; Wang, L.; Wang, Y.; Wang, X.; Hu, M.; Liu, Z. Triple Recycling Processes Impact Systemic and Local Bioavailability of Orally Administered Flavonoids. AAPS J. 2015, 17, 723–736. [Google Scholar] [CrossRef] [Green Version]
- Dantas, A.M.; Mafaldo, I.M.; de Oliveira, P.M.L.; dos Lima, M.S.; Magnani, M.; Borges, G.D. Bioaccessibility of Phenolic Compounds in Native and Exotic Frozen Pulps Explored in Brazil Using a Digestion Model Coupled with a Simulated Intestinal Barrier. Food Chem. 2019, 274, 202–214. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. FAOSTAT Statistical Database; FAO: Rome, Italy, 1997. [Google Scholar]
- Renier, C.; Vandromme, M.; Meyfroidt, P.; Ribeiro, V.; Kalischek, N.; zu Ermgassen, E.K.H.J. Transparency, Traceability and Deforestation in the Ivorian Cocoa Supply Chain. agriRxiv 2022, 2022, 20220421039. [Google Scholar] [CrossRef]
- Schroth, G.; Läderach, P.; Martinez-Valle, A.I.; Bunn, C.; Jassogne, L. Vulnerability to Climate Change of Cocoa in West Africa: Patterns, Opportunities and Limits to Adaptation. Sci. Total Environ. 2016, 556, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Santosa, E.; Sakti, G.P.; Fattah, M.Z.; Zaman, S.; Wahjar, A. Cocoa Production Stability in Relation to Changing Rainfall and Temperature in East Java, Indonesia. JTCS 2018, 5, 6–17. [Google Scholar] [CrossRef]
- Alves, R.M.; Bastos, A.J.R.; da Chaves, S.F.S.; Fernandes, J.R.Q. Spondias Mombin and Theobroma grandiflorum in Agroforestry System: Productivity Implications. Rev. Bras. De Frutic. 2018, 40, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Pereira, A.L.F.; Feitosa, W.S.C.; Abreu, V.K.G.; de Lemos, T.O.; Gomes, W.F.; Narain, N.; Rodrigues, S. Impact of Fermentation Conditions on the Quality and Sensory Properties of a Probiotic Cupuassu (Theobroma grandiflorum) Beverage. Food Res. Int. 2017, 100, 603–611. [Google Scholar] [CrossRef]
- Costa, M.P.; Frasao, B.S.; Silva, A.C.O.; Freitas, M.Q.; Franco, R.M.; Conte-Junior, C.A. Cupuassu (Theobroma grandiflorum) Pulp, Probiotic, and Prebiotic: Influence on Color, Apparent Viscosity, and Texture of Goat Milk Yogurts. J. Dairy Sci. 2015, 98, 5995–6003. [Google Scholar] [CrossRef]
- International trade centre (ITC), Trade Statistics for International Business Development; International Trade Centre: Geneva, Switzerland, 1999.
- Santana, N.B.; Dias, J.C.T.; Rezende, R.P.; Franco, M.; Oliveira, L.K.S.; Souza, L.O. Production of Xylitol and Bio-Detoxification of Cocoa Pod Husk Hemicellulose Hydrolysate by Candida Boidinii XM02G. PLoS ONE 2018, 13, e0195206. [Google Scholar] [CrossRef]
- Lateef, A.; Azeez, M.A.; Asafa, T.B.; Yekeen, T.A.; Akinboro, A.; Oladipo, I.C.; Azeez, L.; Ojo, S.A.; Gueguim-Kana, E.B.; Beukes, L.S. Cocoa Pod Husk Extract-Mediated Biosynthesis of Silver Nanoparticles: Its Antimicrobial, Antioxidant and Larvicidal Activities. J. Nanostructure Chem. 2016, 6, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Oduro-Mensah, D.; Ocloo, A.; Nortey, T.; Antwi, S.; Okine, L.K.; Adamafio, N.A. Nutritional Value and Safety of Animal Feed Supplemented with Talaromyces Verruculosus-Treated Cocoa Pod Husks. Sci. Rep. 2020, 10, 13163. [Google Scholar] [CrossRef]
- Zinla, D.; Gbaha, P.; Koffi, P.M.E.; Koua, B.K. Characterization of Rice, Coffee and Cocoa Crops Residues as Fuel of Thermal Power Plant in Côte d’Ivoire. Fuel 2021, 283, 119250. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sections | Species |
|---|---|
| Andropetalum | T. mammosum Cuatr. & Leon |
| Glossopetalum | T. angustifolium Moçiño & Sesse, T. canumanense Pires & Fróes, T. chocoense Cuatr., T. cirmolinae Cuatr., T. grandiflorum (Willd. ex Spreng.) Schum., T. hylaeum Cuatr., T. nemorale Cuatr., T. obovatum Klotzsch ex Bernoulli, T. simiarum Donn. Smith., T. sinuosum Pavón ex Hubber, T. stipulatum Cuatr., T. subincanum Mart |
| Oreanthes: | T. bernouillii Pittier, T. glaucum Karst, T. speciosum Willd., T. sylvestre Mart, T. velutinum Benoist |
| Rhytidocarpus | T. bicolor Humb. & Bonpl. |
| Telmatocarpus | T. gileri Cuatr., T. microcarpum Mart. |
| Theobroma | T. cacao L. |
| Beans | Pods | |||
|---|---|---|---|---|
| Cupuassu | Cocoa | Cupuassu | Cocoa | |
| Proteins (g/100 g dry weight) | 2.2–26.2 [51,52,53,55] | 2.5–14.4 [54,56,57,58] | - | 2.4–17.6 [45,46,47,48,49,50] |
| Lipids (g/100 g dry weight) | 20–60 [42,51,53] | 20–60 [54] | - | 0.6–2.3 [45,46,48,49,50] |
| Total sugars (g/100 g dry weight) | 1.3–1.6 [66] | 0.1–3.1 [67] | - | 1.7 [45] |
| Polyphenols (TPC-mg AG eq./g dry weight) | 7.8 [85] | 9.8–202.2 [75,79,80] | - | 3.2–56.5 [46,69,77,78] |
| Caffeine (g/100 g dry weight) | 0.05 [68] | 0.2–2.1 [1,51,83,88,89,90,91] | - | 0.002–0.004 [1,51] |
| Theobromine (g/100 g dry weight) | 0.1 [68] | 0.2–7.6 [1,51,83,88,89,90,91] | - | 0.002–0.4 [1,51] |
| Antioxydant activities | Model | Matrix | Action | Sources |
| Cell model-human EA. Hy926 and human HepG2 | Cocoa | ↓ MDA level, ↓ ROS level | [97,98] | |
| ↑ GPx and GR activities | ||||
| Caco-2 epithelial cell and murine STC-1 enteroendocrine cells | Cupuassu | ↓ ROS levels | [99] | |
| Rats model | Cupuassu | ↓ ROS levels | ||
| Ewes model | Cocoa | Not affect AO plasmatic activities, | [101] | |
| ↑ SOD activities, | ||||
| ↓ protein carbonyl levels | ||||
| HFD animal model | Cocoa and Cupuassu | ↑ plasmatic GPx, SOD and hepatic CAT activities | [111] | |
| ↓hepatic and plasmatic MDA levels | ||||
| ↑FRAP and DPPH plasmatic activities | ||||
| Cupuassu | ↓ brain GPx activities | |||
| STZ-induced diabete rats model | Cocoa and Cupuassu | ↑CAT kidney, SOD plasma and plasmatic GPx | [85] | |
| ↑ plasmatic, liver and kidney FRAP and plasmatic ORAC activities | ||||
| Cocoa | ↑ liver GPx activity | |||
| Cupuassu | ↑ bain CAT and SOD, | |||
| ↑ kidney GPx | ||||
| Immunomodulatory activities | human THP-1 macrophages M1 model | Cocoa | ↓TNF-α, IL-6, IL-1β, and IL-12 levels, | [120] |
| ↑ IL-10 level, | ||||
| Provide M1/M2 metabolic switch (similar levels of IL-10 and IL-12) | ||||
| MiMC cell model | Cupuassu | ↓ NO and ROS levels, | [121] | |
| in vivo diabetic rat model | Cupuassu | ↓ renal NO level, kidney eNOS and 3-nitrotyrosine, | ||
| ↓ renal NF-KB and IL-6 levels | ||||
| in vivo TNBS-induced rats model | Cupuassu | ↓ neutrophil myeloperoxidase and alkaline phosphatase activities, | [122] | |
| ↓ IL-6 and IL-1 levels | ||||
| ovalbumin-sensitized rats models | Cocoa | ↓ OVA-specific antibodies IgM, IgG1, IgG2a and IgG2c and IgE levels | [125,126] | |
| Rat model | Cocoa | ↓ the levels of CD8+, CD4- (and CD8-, CD4-) TCRa-b low (immature) cells, | [137] | |
| ↓ CD8+, CD4+ thymphocytes | ||||
| ↑ CD8-, CD4- and both CD4+ cells levels | ||||
| Impact on intestinal tract | Caco2 cells | Cocoa | ↓ IL8 and MCP1 levels, restoring claudin-1, occludin and JAM-1 levels | [127] |
| ZDF rats | Cocoa | ↓ TNF-α, IL-6 and MCP1 levels, and ↓ cd45 levels | [136] | |
| restauring ZO-1 | ||||
| HFD rats | Cocoa | ↓ ALT and AST activities | [107] | |
| ZDF rats | Cocoa | ↑ Proteobacteria, Tenericutes and Actinobacteria phyla, | [128] | |
| ↑ firmicutes and deferribacteres phyla, | ||||
| ↓ cyanobacteries phylum | ||||
| Clinical trials | Human | Cocoa | ↑ Blautia and Lachnospira genera, | [133,134] |
| ↓ Agathobacter genus and Faecalibacterium prausnitzii | ||||
| Cocoa | modulated the expression of gene involving in ↓ of ROS production, Ca2+ modulation and activation of leukocytes and viral response. | [138] | ||
| Cocoa | ↓ IL6, malondialdehyde and protein carbonyl levels, ↑ 6-min walked distance assay, ↓ sit-up test, ↑ hand strength assay | [139] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jean-Marie, E.; Jiang, W.; Bereau, D.; Robinson, J.-C. Theobroma cacao and Theobroma grandiflorum: Botany, Composition and Pharmacological Activities of Pods and Seeds. Foods 2022, 11, 3966. https://doi.org/10.3390/foods11243966
Jean-Marie E, Jiang W, Bereau D, Robinson J-C. Theobroma cacao and Theobroma grandiflorum: Botany, Composition and Pharmacological Activities of Pods and Seeds. Foods. 2022; 11(24):3966. https://doi.org/10.3390/foods11243966
Chicago/Turabian StyleJean-Marie, Elodie, Weiwen Jiang, Didier Bereau, and Jean-Charles Robinson. 2022. "Theobroma cacao and Theobroma grandiflorum: Botany, Composition and Pharmacological Activities of Pods and Seeds" Foods 11, no. 24: 3966. https://doi.org/10.3390/foods11243966
APA StyleJean-Marie, E., Jiang, W., Bereau, D., & Robinson, J. -C. (2022). Theobroma cacao and Theobroma grandiflorum: Botany, Composition and Pharmacological Activities of Pods and Seeds. Foods, 11(24), 3966. https://doi.org/10.3390/foods11243966