
Metabolomic Analysis Reveals Nutritional Diversity among Three Staple Crops and Three Fruits

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Chemical Reagents
2.3. Metabolite Sample Preparation
2.4. Metabolomic Detection
2.5. Statistical Analysis
3. Results
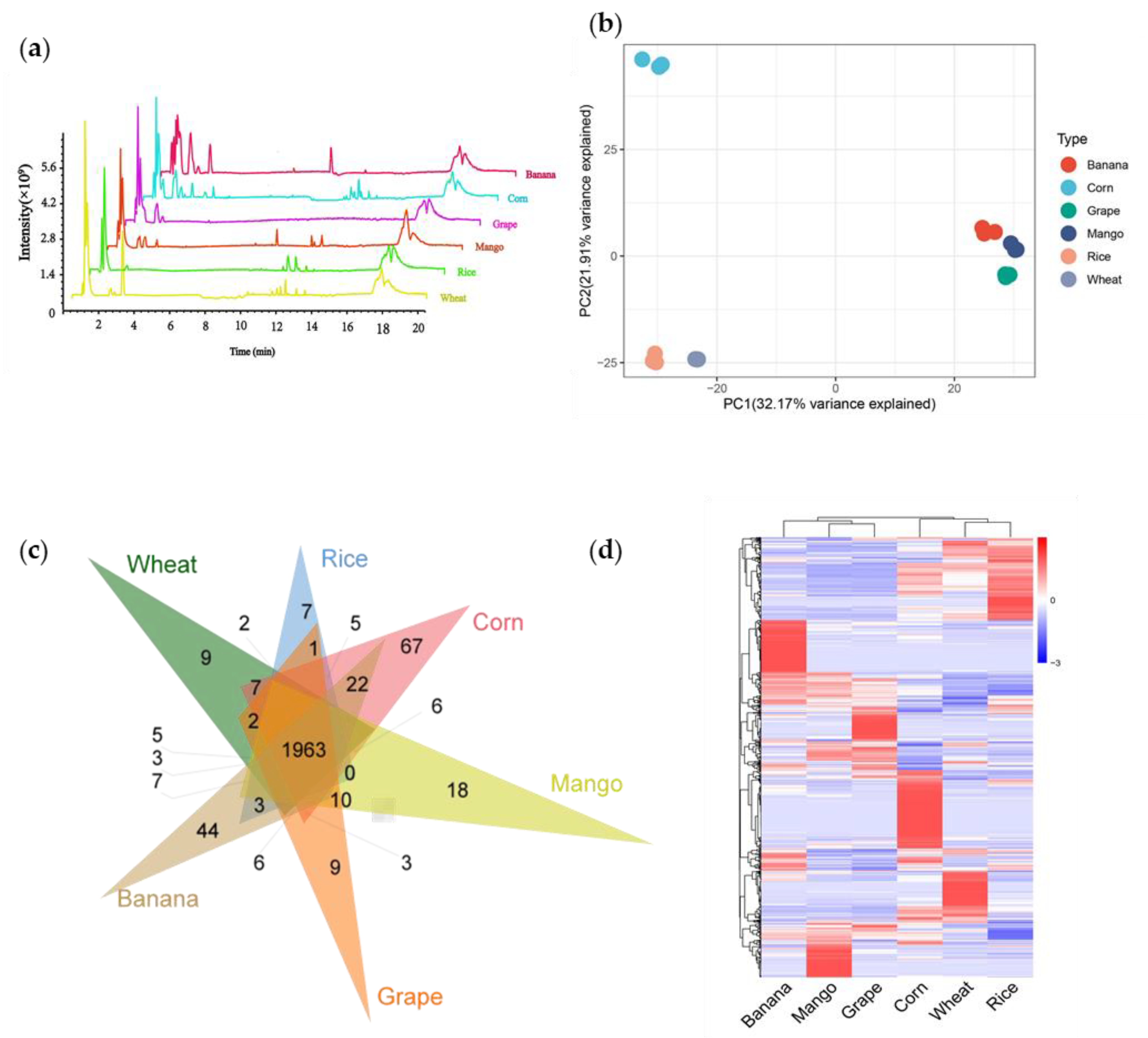
3.1. Metabolic Analysis of Crops and Fruits
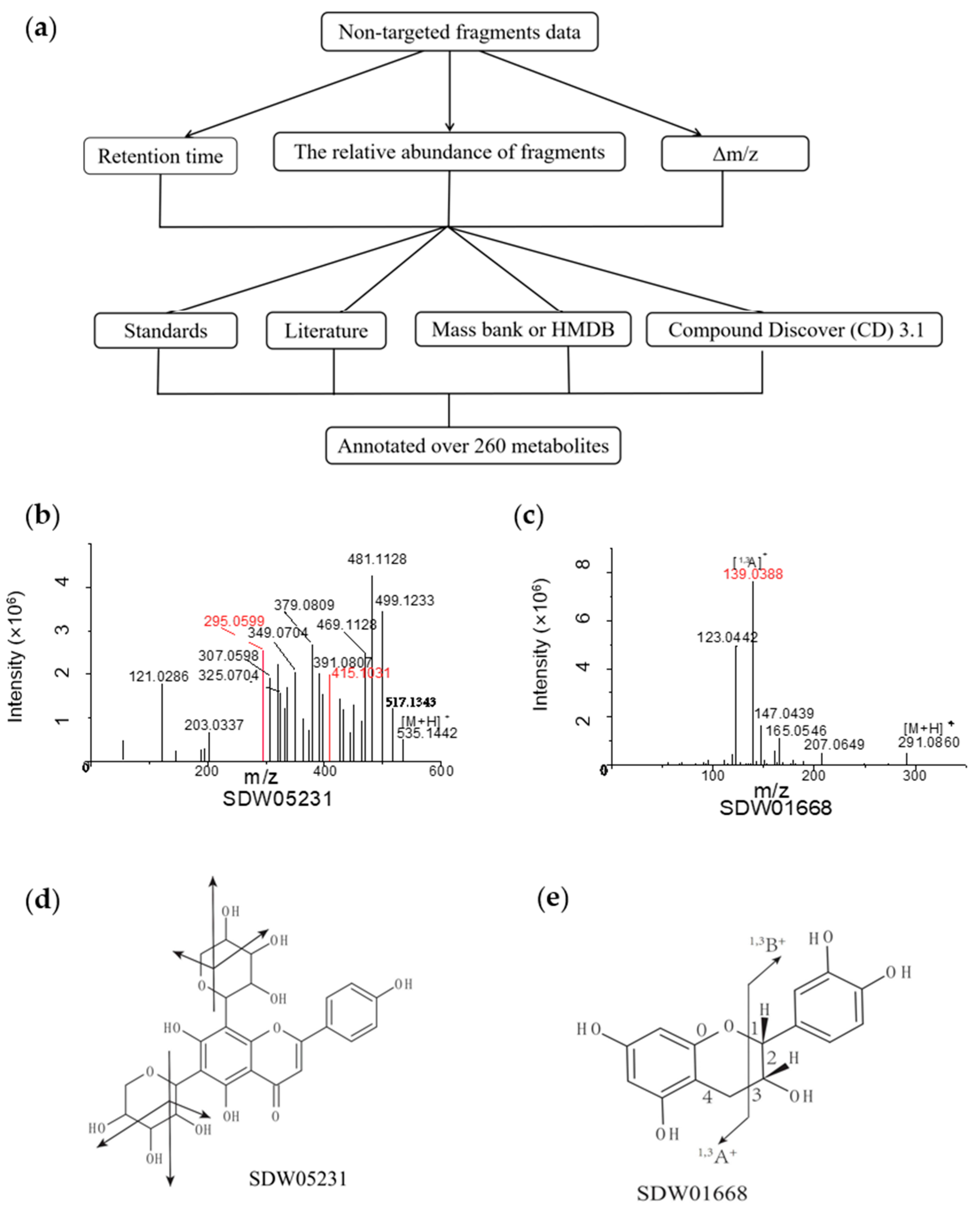
3.2. Characterization of Metabolic Signals
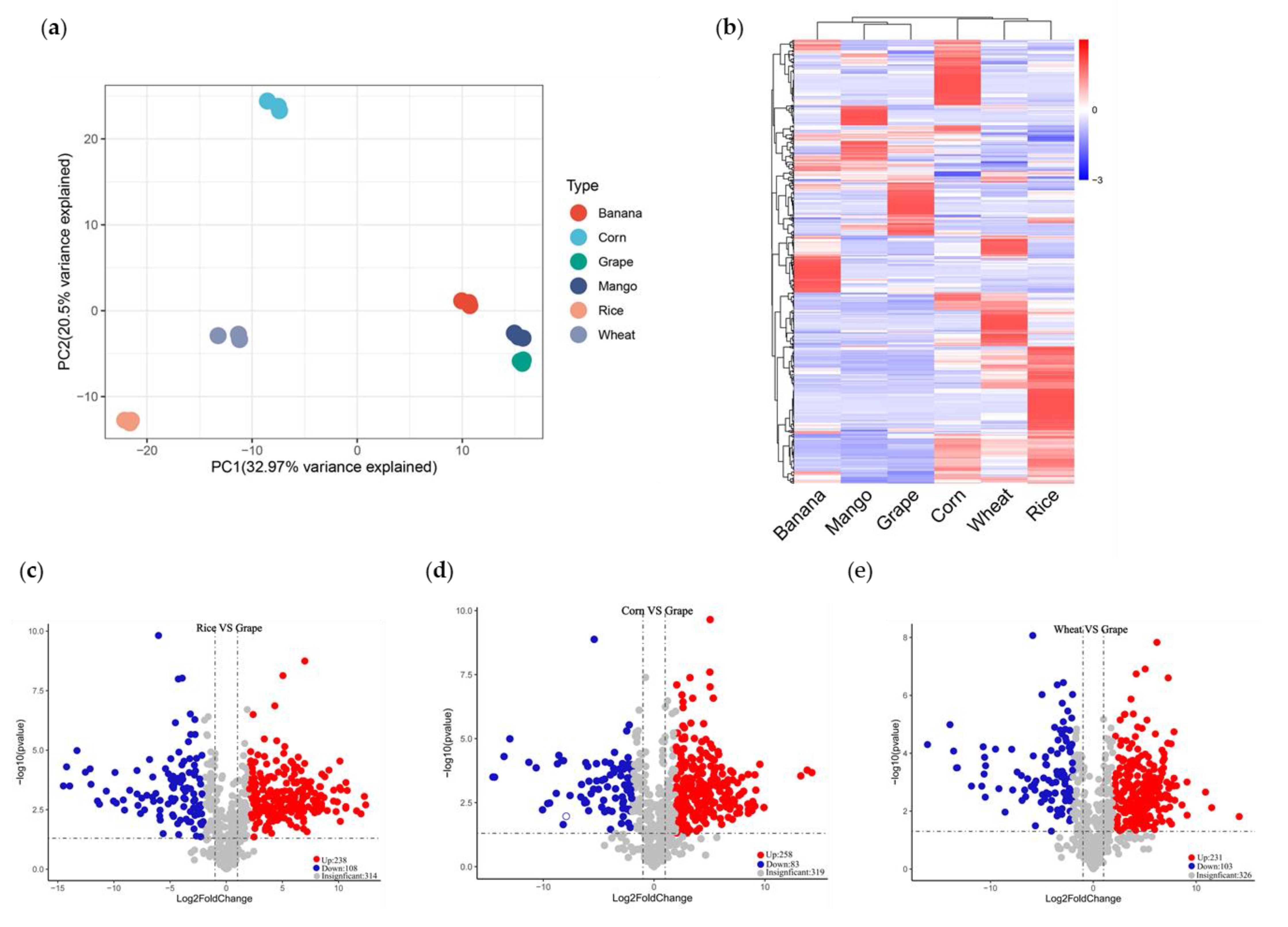
3.3. Whole Metabolome Scale Comparative Analysis of Crops and Fruits
3.4. Comparative Analysis of Common Existing Nutrients in Crops and Fruits
3.5. Comparative Analysis of Species-Specific Nutrients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hindu, V.; Palacios-Rojas, N.; Babu, R.; Suwarno, W.; Rashid, Z.; Usha, R.; Saykhedkar, G.; Nair, S. Identification and validation of genomic regions influencing kernel zinc and iron in maize. Thero. Appl. Genet. 2018, 131, 1443–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, N. The global challenge of hidden hunger: Perspectives from the field. Proc. Nutr. Soc. 2021, 80, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gashu, D.; Nalivata, P.; Amede, T.; Ander, E.; Bailey, E.; Botoman, L.; Chagumaira, C.; Gameda, S.; Haefele, S.; Hailu, K.; et al. The nutritional quality of cereals varies geospatially in Ethiopia and Malawi. Nature 2021, 594, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Palacios-Rojas, N.; McCulley, L.; Kaeppler, M.; Titcomb, T.; Gunaratna, N.; Lopez-Ridaura, S.; Tanumihardjo, S. Mining maize diversity and improving its nutritional aspects within agro-food systems. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1809–1834. [Google Scholar] [CrossRef] [PubMed]
- Goredema-Matongera, N.; Ndhlela, T.; Magorokosho, C.; Kamutando, C.; van Biljon, A.; Labuschagne, M. Multinutrient Biofortification of Maize (Zea mays L.) in Africa: Current Status, Opportunities and Limitations. Nutrients 2021, 13, 1039. [Google Scholar] [CrossRef] [PubMed]
- Rondanelli, M.; Miccono, A.; Peroni, G.; Nichetti, M.; Infantino, V.; Spadaccini, D.; Alalwan, T.; Faliva, M.; Perna, S. Rice germ macro- and micronutrients: A new opportunity for the nutraceutics. Nat. Prod. Res. 2021, 35, 1532–1536. [Google Scholar] [CrossRef]
- Yu, S.; Tian, L. Breeding Major Cereal Grains through the Lens of Nutrition Sensitivity. Mol. Plant 2018, 11, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Liu, R. Potential synergy of phytochemicals in cancer prevention: Mechanism of action. J. Nutr. 2004, 134, 3479S–3485S. [Google Scholar] [CrossRef] [Green Version]
- Liu, R. Dietary bioactive compounds and their health implications. J. Food Sci. 2013, 78, A18–A25. [Google Scholar] [CrossRef]
- Brookie, K.; Best, G.; Conner, T. Intake of Raw Fruits and Vegetables Is Associated with Better Mental Health Than Intake of Processed Fruits and Vegetables. Front. Psychol. 2018, 9, 487. [Google Scholar] [CrossRef]
- Duthie, S.; Duthie, G.; Russell, W.; Kyle, J.; Macdiarmid, J.; Rungapamestry, V.; Stephen, S.; Megias-Baeza, C.; Kaniewska, J.; Shaw, L.; et al. Effect of increasing fruit and vegetable intake by dietary intervention on nutritional biomarkers and attitudes to dietary change: A randomised trial. Eur. J. Nutr. 2018, 57, 1855–1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ediriweera, M.; Tennekoon, K.; Samarakoon, S. A Review on Ethnopharmacological Applications, Pharmacological Activities, and Bioactive Compounds of Mangifera indica (Mango). Evid. Based Complement Altern. Med. 2017, 2017, 6949835. [Google Scholar] [CrossRef] [Green Version]
- Ruocco, S.; Stefanini, M.; Stanstrup, J.; Perenzoni, D.; Mattivi, F.; Vrhovsek, U. The metabolomic profile of red non-V. vinifera genotypes. Food Res. Int. (Ott. Ont.) 2017, 98, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Celis, M.; Yahia, E.; Bedoya, R.; Landázuri, P.; Loango, N.; Aguillón, J.; Restrepo, B.; Guerrero Ospina, J. Chemical Composition of Mango (Mangifera indica L.) Fruit: Nutritional and Phytochemical Compounds. Front. Plant Sci. 2019, 10, 1073. [Google Scholar] [CrossRef]
- Lebaka, V.; Wee, Y.; Ye, W.; Korivi, M. Nutritional Composition and Bioactive Compounds in Three Different Parts of Mango Fruit. Int. J. Environ. Res. Public Health 2021, 18, 741. [Google Scholar] [CrossRef] [PubMed]
- Mwaurah, P.; Kumar, S.; Kumar, N.; Panghal, A.; Attkan, A.; Singh, V.; Garg, M. Physicochemical characteristics, bioactive compounds and industrial applications of mango kernel and its products: A review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2421–2446. [Google Scholar] [CrossRef]
- Fu, X.; Cheng, S.; Feng, C.; Kang, M.; Huang, B.; Jiang, Y.; Duan, X.; Grierson, D.; Yang, Z. Lycopene cyclases determine high α-/β-carotene ratio and increased carotenoids in bananas ripening at high temperatures. Food Chem. 2019, 283, 131–140. [Google Scholar] [CrossRef]
- Liu, Q.; Tang, G.; Zhao, C.; Feng, X.; Xu, X.; Cao, S.; Meng, X.; Li, S.; Gan, R.; Li, H. Comparison of Antioxidant Activities of Different Grape Varieties. Molecules 2018, 23, 2432. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Laskar, M.; Sarker, S.; Choudhury, M.; Choudhury, P.; Mitra, A.; Jamil, S.; Lathiff, S.; Abdullah, S.; Basar, N.; et al. Prediction of Anti-Alzheimer’s Activity of Flavonoids Targeting Acetylcholinesterase in silico. Phytochem. Anal. 2017, 28, 324–331. [Google Scholar] [CrossRef]
- Chen, J.; Hu, X.; Shi, T.; Yin, H.; Sun, D.; Hao, Y.; Xia, X.; Luo, J.; Fernie, A.; He, Z.; et al. Metabolite-based genome-wide association study enables dissection of the flavonoid decoration pathway of wheat kernels. Plant Biotechnol. J. 2020, 18, 1722–1735. [Google Scholar] [CrossRef]
- Xu, G.; Cao, J.; Wang, X.; Chen, Q.; Jin, W.; Li, Z.; Tian, F. Evolutionary Metabolomics Identifies Substantial Metabolic Divergence between Maize and Its Wild Ancestor, Teosinte. Plant Cell 2019, 31, 1990–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, M.; Shahzad, R.; Gul, A.; Subthain, H.; Shen, S.; Lei, L.; Zheng, Z.; Zhou, J.; Lu, D.; Wang, S.; et al. Differentially evolved glucosyltransferases determine natural variation of rice flavone accumulation and UV-tolerance. Nat. Commun. 2017, 8, 1975. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.; Zhu, A.; Jia, J.; Hu, X.; Chen, J.; Liu, W.; Ren, X.; Sun, D.; Fernie, A.; Cui, F.; et al. Metabolomics analysis and metabolite-agronomic trait associations using kernels of wheat (Triticum aestivum) recombinant inbred lines. Plant J. 2020, 103, 279–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, W.; Jin, M.; Li, K.; Liu, H.; Xiao, Y.; Zhao, M.; Alseekh, S.; Li, W.; de Abreu, E.; Lima, F.; et al. An integrated multi-layered analysis of the metabolic networks of different tissues uncovers key genetic components of primary metabolism in maize. Plant J. 2018, 93, 1116–1128. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Gao, Y.; Chen, W.; Wang, W.; Gong, L.; Liu, X.; Luo, J. Spatio-temporal distribution of phenolamides and the genetics of natural variation of hydroxycinnamoyl spermidine in rice. Mol. Plant 2015, 8, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Peng, Z.; Zhang, H.; Li, W.; Yuan, Z.; Xie, Z.; Zhang, H.; Cheng, Y.; Chen, J.; Xu, J. Comparative profiling and natural variation of polymethoxylated flavones in various citrus germplasms. Food Chem. 2021, 354, 129499. [Google Scholar] [CrossRef]
- Lombardo, V.; Osorio, S.; Borsani, J.; Lauxmann, M.; Bustamante, C.; Budde, C.; Andreo, C.; Lara, M.; Fernie, A.; Drincovich, M. Metabolic profiling during peach fruit development and ripening reveals the metabolic networks that underpin each developmental stage. Mol. Plant 2011, 157, 1696–1710. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Yu, S.; Xiong, L.; Luo, J. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: Application in the study of rice metabolomics. Mol. Plant 2013, 6, 1769–1780. [Google Scholar] [CrossRef] [Green Version]
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K.; et al. MassBank: A public repository for sharing mass spectral data for life sciences. J. Proteome Res. 2010, 45, 703–714. [Google Scholar] [CrossRef]
- Wishart, D.; Tzur, D.; Knox, C.; Eisner, R.; Guo, A.; Young, N.; Cheng, D.; Jewell, K.; Arndt, D.; Sawhney, S.; et al. HMDB: The Human Metabolome Database. Nucleic Acids Res. 2007, 35, D521–D526. [Google Scholar] [CrossRef]
- Chen, W.; Wang, W.; Peng, M.; Gong, L.; Gao, Y.; Wan, J.; Wang, S.; Shi, L.; Zhou, B.; Li, Z.; et al. Comparative and parallel genome-wide association studies for metabolic and agronomic traits in cereals. Nat. Commun. 2016, 7, 12767. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Li, K.; Shi, Y.; Li, Y.; Dong, L.; Liu, L.; Li, M.; Ren, H.; Liu, X.; Fang, C.; et al. Cross-Species Comparison of Metabolomics to Decipher the Metabolic Diversity in Ten Fruits. Metabolites 2021, 11, 164. [Google Scholar] [CrossRef]
- Deng, M.; Zhang, X.; Luo, J.; Liu, H.; Wen, W.; Luo, H.; Yan, J.; Xiao, Y. Metabolomics analysis reveals differences in evolution between maize and rice. Plant J. 2020, 103, 1710–1722. [Google Scholar] [CrossRef] [PubMed]
- Zhu, A.; Zhou, Q.; Hu, S.; Wang, F.; Tian, Z.; Hu, X.; Liu, H.; Jiang, D.; Chen, W. Metabolomic analysis of the grain pearling fractions of six bread wheat varieties. Food Chem. 2022, 369, 130881. [Google Scholar] [CrossRef] [PubMed]
- Schneider, G.; Salazar, D.; Hildreth, S.; Helm, R.; Whitehead, S. Comparative Metabolomics of Fruits and Leaves in a Hyperdiverse Lineage Suggests Fruits Are a Key Incubator of Phytochemical Diversification. Front Plant Sci. 2021, 12, 693739. [Google Scholar] [CrossRef] [PubMed]
- Hectors, K.; Van Oevelen, S.; Geuns, J.; Guisez, Y.; Jansen, M.; Prinsen, E. Dynamic changes in plant secondary metabolites during UV acclimation in Arabidopsis thaliana. Physiol. Plant. 2014, 152, 219–230. [Google Scholar] [CrossRef]
- Fang, C.; Luo, J. Metabolic GWAS-based dissection of genetic bases underlying the diversity of plant metabolism. Plant J. 2019, 97, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Navarro, J.; Izquierdo-Cañas, P.; Mena-Morales, A.; Martínez-Gascueña, J.; Chacón-Vozmediano, J.; García-Romero, E.; Hermosín-Gutiérrez, I.; Gómez-Alonso, S. Phenolic compounds profile of different berry parts from novel Vitis vinifera L. red grape genotypes and Tempranillo using HPLC-DAD-ESI-MS/MS: A varietal differentiation tool. Food Chem. 2019, 295, 350–360. [Google Scholar] [CrossRef]
- Barbosa Gámez, I.; Caballero Montoya, K.; Ledesma, N.; Sáyago Ayerdi, S.; García Magaña, M.; Bishop von Wettberg, E.; Montalvo-González, E. Changes in the nutritional quality of five Mangifera species harvested at two maturity stages. J. Sci. Food Agric. 2017, 97, 4987–4994. [Google Scholar] [CrossRef]
- Tan, L.; Jin, Z.; Ge, Y.; Nadeem, H.; Cheng, Z.; Azeem, F.; Zhan, R. Comprehensive ESI-Q TRAP-MS/MS based characterization of metabolome of two mango (Mangifera indica L.) cultivars from China. Sci. Rep. 2020, 10, 20017. [Google Scholar] [CrossRef]
- Ma, W.; Waffo-Téguo, P.; Jourdes, M.; Li, H.; Teissedre, P. First evidence of epicatechin vanillate in grape seed and red wine. Food Chem. 2018, 259, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Sabaghi, M.; Hoseyni, S.; Tavasoli, S.; Mozafari, M.; Katouzian, I. Strategies of confining green tea catechin compounds in nano-biopolymeric matrices: A review. Colloids Surf. B Biointerfaces 2021, 204, 111781. [Google Scholar] [CrossRef] [PubMed]
- Xia, E.; Deng, G.; Guo, Y.; Li, H. Biological activities of polyphenols from grapes. Int. J. Mol. Sci. 2010, 11, 622–646. [Google Scholar] [CrossRef] [PubMed]
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2021.Transforming food Systems for Food Security, Improved Nutrition and Affordable Healthy Diets for All; FAO: Rome, Italy, 2021. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | ID | Q1 (Da) | RT (min) | Compounds | Class |
|---|---|---|---|---|---|
| Crops | hsf030 | 301.1 | 5.63 | Diosmetin | Flavonoid |
| MP0307 | 301.05 | 8.04 | Chrysoeriol | Flavonoid | |
| MP0566 | 565.1 | 5.64 | C-pentosyl-apigenin O-hexoside | Flavonoid | |
| MP0577 | 581.1 | 4.98 | C-pentosyl-luteolin O-hexoside | Flavonoid | |
| hsl047 | 325.3 | 12.49 | Heneicosanoic acid (C21:0) | Lipids | |
| MP0557 | 548.35 | 10.8 | LysoPC 20:2 | Lipids | |
| MP2054 | 307.3 | 11.39 | cis-11,14,17-Eicosatrienoic Acid (C20:3) | Lipids | |
| hsb025 | 166.04 | 3.46 | 2-(Formylamino)benzoic acid | Others | |
| MP0885 | 287 | 12.14 | 5-deoxo-ent-10-oxodepressin | Others | |
| hsa126 | 161.2 | 0.72 | D-Alanyl-D-Alanine | Amino acid and its derivatives | |
| MP0576 | 580.9 | 5.18 | inositol pentakisphosphate | Organic acid and its derivates | |
| Fruits | hsc303 | 307 | 3.5 | (-)-Epigallocatechin | Flavonoid |
| hsc317 | 291.08 | 3.58 | Catechin | Flavonoid | |
| hsc325 | 307 | 2.57 | (+)-Gallocatechin | Flavonoid | |
| hsc327 | 307 | 2.12 | (-)-gallatechin | Flavonoid | |
| hsf421 | 451 | 3.72 | Eriodictyol 7-O-glucoside | Flavonoid |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Y.; Guo, Y.; Wang, Y.; Li, M.; Li, K.; Liu, X.; Fang, C.; Luo, J. Metabolomic Analysis Reveals Nutritional Diversity among Three Staple Crops and Three Fruits. Foods 2022, 11, 550. https://doi.org/10.3390/foods11040550
Shi Y, Guo Y, Wang Y, Li M, Li K, Liu X, Fang C, Luo J. Metabolomic Analysis Reveals Nutritional Diversity among Three Staple Crops and Three Fruits. Foods. 2022; 11(4):550. https://doi.org/10.3390/foods11040550
Chicago/Turabian StyleShi, Yunxia, Yanxiu Guo, Yuhui Wang, Mingyang Li, Kang Li, Xianqing Liu, Chuanying Fang, and Jie Luo. 2022. "Metabolomic Analysis Reveals Nutritional Diversity among Three Staple Crops and Three Fruits" Foods 11, no. 4: 550. https://doi.org/10.3390/foods11040550
APA StyleShi, Y., Guo, Y., Wang, Y., Li, M., Li, K., Liu, X., Fang, C., & Luo, J. (2022). Metabolomic Analysis Reveals Nutritional Diversity among Three Staple Crops and Three Fruits. Foods, 11(4), 550. https://doi.org/10.3390/foods11040550