
Plant Milk-Clotting Enzymes for Cheesemaking

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Types and Characteristics of Vegetable Proteases
2.1. Aspartic Proteases
2.2. Cysteine Proteases
2.3. Serine Proteases

{kind=link}
| Plant Source | Tissues | Type | Strengths | Weaknesses | Temperature (°C) | pH | Reference |
|---|---|---|---|---|---|---|---|
| Calotropis gigantea | Latex, stem, flower, and leaf | CP, SP | Latex has the highest MCA/PA ratio | High rate of proteolysis of crude enzyme | 37 | 5.5 | [68] |
| Citrus aurantium | Flower | AP | Raw enzyme extract is capable of coagulating milk in similar times to that required by animal rennet | NR | 65–70 | 4.0 | [20] |
| Zingiber officinale | Rhizomes | CP | The vegetable coagulant is easily extracted through few purification steps | Further studies are needed for industrial application | 60 | 5.5 | [52] |
| Silybum marianum | Flower | AP | Proteases extracted from Silybum marianum clot bovine, caprine, and ovine milks | NR | NA | NA | [71] |
| Balanites aegyptiaca | Fruit | AP, SP | MCA was found from the extract in the fruit pulp | Further studies on the organoleptic acceptability of cheeses produced are necessary | 50 | 5.0, 8.0 | [70] |
| Cynara scolymus | Flower | AP | Cheese yield is similar to that of animal rennet | Prolonged brining period (40 h) is necessary to avoid the development of bitter flavors in the cheese | 40–60 | 4.0 | [72,73] |
| Foeniculum vulgare | Stems | SP | The proteases are active at the temperature and pH parameters used for cheesemaking | The extraction process is complex | 37 | 6.4 | [66] |
| Dregea sinensis | Stems | CP | Purified cysteine protease shows a wide range of activity (pH and temperature) | The optimum temperature is about 80 °C, which is not adopted in the cheesemaking process | 80 | 6.0–9.0 | [56] |
| Bromelia pinguin | Fruit | CP, SP | The enzyme extract is able to coagulate milk in a relatively short period of time | High caseinolytic activity after a long incubation period | 45 | 2.5, 7.5 | [69] |
| Morinda citrifolia | Fruit | CP | With an MCA value of 238.80 ± 5.29 U/mL, Morinda citrifolia fruit extract proves to be a good candidate to replace calf rennet | Slightly bitter taste but good acceptability of cheeses | 50 | 6.0, 7.0 | [55] |
| Vallesia glabra | Leaf, fruits, and seed | AP, CP, SP | The extract obtained from the leaves shows a great activity (0.20 MCU/mL) while in the fruits and seeds it was 0.12 and 0.11 MCU/mL, respectively | Further studies are needed to better characterize the wide variety of proteases present in the raw extract | 65–70 | 4.0 | [67] |
| Solanum tuberosum | Tubers and leaves | AP | The two aspartic proteases are able to operate at optimal cheesemaking conditions (temperature 40–42 °C, pH values 6–6.2) | NR | 30 | 5.0 | [47] |
| Salpichroa origanifolia | Fruit | AP | The activity of the enzyme allows to enrich the cheese with bioactive peptides deriving from the hydrolysis of α-, β-, and κ-casein, which provide a health-promoting effect | NR | 40 | 6.0 | [32] |
| Ficus johannis | Latex | CP | The low tendency to autolysis, that is, autodigestion during storage at room temperature, suggesting a probable use in industrial cheesemaking | Enzyme loses 20% activity at high salt concentrations (1 M NaCl) | 60 | 6.5 | [74] |
| Solanum elaeagnifolium | Fruit | NA | This plant-derived protease is characterized by a good MCA/PA ratio | High concentrations of this coagulant can negatively affect the visco-elastic properties of the cheese | 45 | 6.0 | [75] |
| Actinidia chinensis | Fruit | CP | Kiwi extract exhibits high MCA/PA ratio compared to other plant coagulants | NR | 40 | 5.5 | [58] |
| Cynara cardunculus | Hairy root cultures | AP, SP, CP | Cynara cardunculus roots can produce proteases such as cardosin A with MCA, the phenotypic characteristic of high growth could lead to continuous supply from an application point of view | The low concentration of these proteases prevents industrial implementation | NA | NA | [76] |
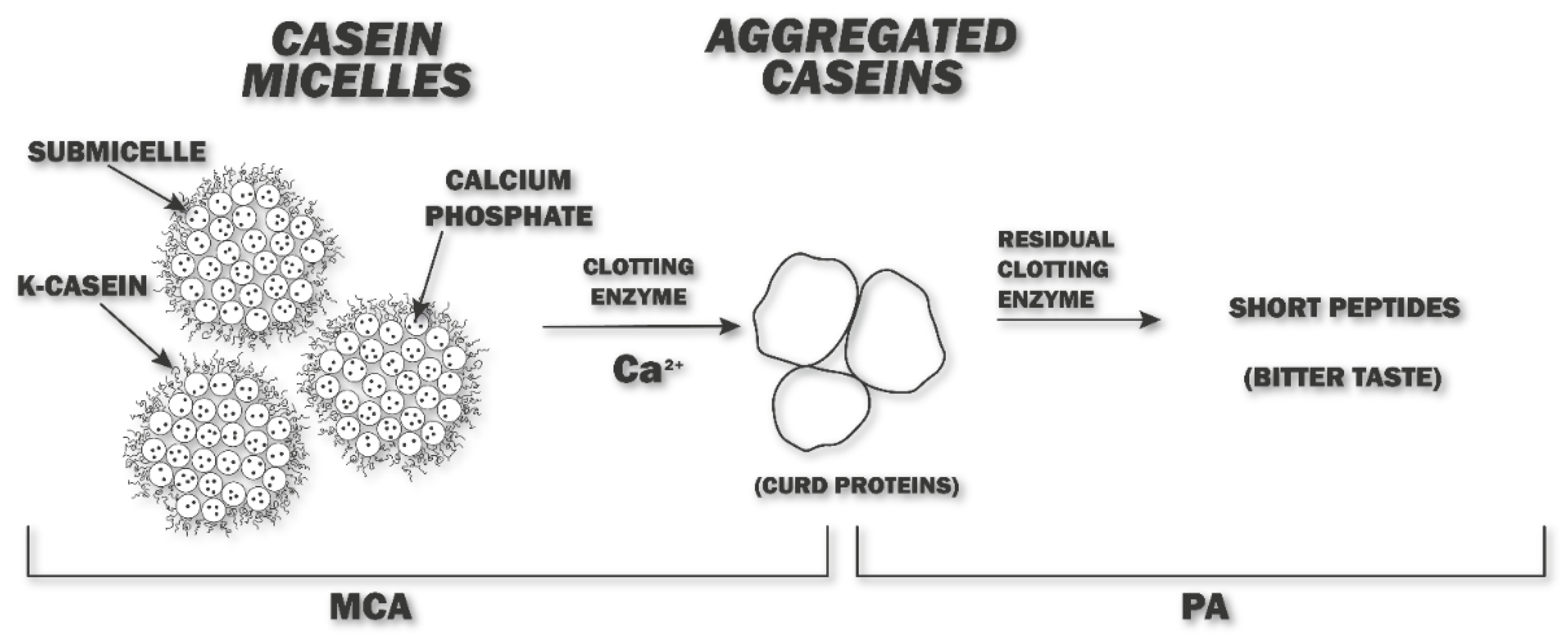
3. Proteolytic Activity and Bitter Taste by Plant Derived Coagulant in Cheese
4. Effect of Plant Coagulants on Cheeses
5. Strategy to Improve the Use of Plant Derived Coagulant
6. Conclusions and Future Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kumar, A.; Grover, S.; Sharma, J.; Batish, V.K. Chymosin and Other Milk Coagulants: Sources and Biotechnological Interventions. Crit. Rev. Biotechnol. 2010, 30, 243–258. [Google Scholar] [CrossRef]
- Britten, M.; Giroux, H.J. Rennet Coagulation of Heated Milk: A Review. Int. Dairy J. 2022, 124, 105179. [Google Scholar] [CrossRef]
- Lee, M.R.; Lee, W.J. The Role of Ca Equilibrium on the Functional Properties of Cheese: A Review. Food Sci. Anim. Resour. 2009, 29, 545–549. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Horne, D.S.; Lucey, J.A. Effect of Insoluble Calcium Concentration on Rennet Coagulation Properties of Milk. J. Dairy Sci. 2007, 90, 2612–2623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucey, J.A.; Fox, P.F. Importance of calcium and phosphate in cheese manufacture: A review. J. Dairy Sci. 1993, 76, 1714–1724. [Google Scholar] [CrossRef]
- Ozcan, T.; Kurdal, E. The Effects of Using a Starter Culture, Lipase, and Protease Enzymes on Ripening of Mihalic Cheese. Int. J. Dairy Technol. 2012, 65, 585–593. [Google Scholar] [CrossRef]
- Nitu, S.; Geicu-Cristea, M.; Matei, F. Milk-Clotting Enzymes Obtained from Plants in Cheesemaking—A Review. Sci. Bull. Ser. F Biotechnol. 2021, 25, 66–75. [Google Scholar]
- Jacob, M.; Jaros, D.; Rohm, H. Recent Advances in Milk Clotting Enzymes. Int. J. Dairy Technol. 2011, 64, 14–33. [Google Scholar] [CrossRef]
- Aktayeva, S.; Akishev, Z.; Khassenov, B. Proteolytic Enzymes in Cheese Making. Eurasian J. Appl. Biotechnol. 2018, 1, 10–16. [Google Scholar] [CrossRef]
- Feijoo-Siota, L.; Blasco, L.; Rodriguez-Rama, J.L.; Barros-Velázquez, J.; de Miguel, T.; Sánchez-Pérez, A.; Villa, G.T. Recent patents on microbial proteases for the dairy industry. Recent Adv. DNA Gene Seq. 2014, 8, 44–55. [Google Scholar] [CrossRef]
- Lebedev, L.R.; Kosogova, T.A.; Teplyakova, T.V.; Kriger, A.V.; Elchaninov, V.V.; Belov, A.N.; Koval, A.D. Study of Technological Properties of Milk-clotting Enzyme from Irpex lacteus. Foods Raw Mater. 2016, 4, 58–65. [Google Scholar] [CrossRef]
- He, X.; Ren, F.; Guo, H.; Zhang, W.; Song, X.; Gan, B. Purification and Properties of a Milk-clotting Enzyme Produced by Bacillus amyloliquefaciens D4. Korean J. Chem. Eng. 2011, 28, 203–208. [Google Scholar] [CrossRef]
- Claverie-Martin, F.; Vega-Hernàndez, M.C. Aspartic Proteases Used in Cheese Making. In Industrial Enzymes; Springer: Dordrecht, The Netherlands, 2007; pp. 207–219. [Google Scholar]
- Flamm, E.L. How FDA Approved Chymosin: A Case History. Nat. Biotechnol. 1991, 9, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Harboe, M.; Broe, M.L.; Qvist, K.B. The production, action and application of rennet and coagulants. In Technology of Cheesemaking, 2nd ed.; Law, B.A., Tamime, A.Y., Eds.; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2010; pp. 98–129. [Google Scholar]
- Egito, A.S.; Girardet, J.-M.; Laguna, L.E.; Poirson, C.; Mollé, D.; Miclo, L.; Humbert, G.; Gaillard, J.L. Milk-Clotting Activity of Enzyme Extracts from Sunflower and Albizia Seeds and Specific Hydrolysis of Bovine κ-Casein. Int. Dairy J. 2007, 17, 816–825. [Google Scholar] [CrossRef]
- Liu, X.; Wu, Y.; Guan, R.; Jia, G.; Ma, Y.; Zhang, Y. Advances in Research on Calf Rennet Substitutes and Their Effects on Cheese Quality. Food Res. Int. 2021, 149, 110704. [Google Scholar] [CrossRef]
- Zikiou, A.; Esteves, A.C.; Esteves, E.; Rosa, N.; Gomes, S.; Martins, A.P.L.; Zidoune, M.N.; Barros, M. Algerian Cardoon Flowers Express a Large Spectrum of Coagulant Enzymes with Potential Applications in Cheesemaking. Int. Dairy J. 2020, 105, 104689. [Google Scholar] [CrossRef]
- Bueno-Gavilá, E.; Abellán, A.; Bermejo, M.S.; Salazar, E.; Cayuela, J.M.; Prieto-Merino, D.; Tejada, L. Characterization of Proteolytic Activity of Artichoke (Cynara scolymus L.) Flower Extracts on Bovine Casein to Obtain Bioactive Peptides. Animals 2020, 10, 914. [Google Scholar] [CrossRef]
- Mazorra-Manzano, M.A.; Moreno-Hernández, J.M.; Ramírez-Suarez, J.C.; de Torres-Llanez, M.J.; González-Córdova, A.F.; Vallejo-Córdoba, B. Sour Orange Citrus aurantium L. Flowers: A New Vegetable Source of Milk-Clotting Proteases. LWT Food Sci. Technol. 2013, 54, 325–330. [Google Scholar] [CrossRef]
- Shah, M.A.; Mir, S.A.; Paray, M.A. Plant Proteases as Milk-Clotting Enzymes in Cheesemaking: A Review. Dairy Sci. Technol. 2014, 94, 5–16. [Google Scholar] [CrossRef]
- Almeida, C.M.; Simões, I. Cardoon-Based Rennets for Cheese Production. Appl. Microbiol. Biotechnol. 2018, 102, 4675–4686. [Google Scholar] [CrossRef]
- Schaller, A. A Cut above the Rest: The Regulatory Function of Plant Proteases. Planta 2004, 220, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Mazorra-Manzano, M.A.; Moreno-Hernández, J.M.; Ramírez-Suarez, J.C. Milk-clotting plant proteases for cheesemaking. In Biotechnological Applications of Plant Proteolytic Enzymes; Springer International Publishing: Cham, Switzerland, 2018; pp. 21–41. ISBN 9783319971322. [Google Scholar]
- Ahmed, I.A.M.; Babiker, E.E.; Mori, N. pH Stability and Influence of Salts on Activity of a Milk-Clotting Enzyme from Solanum dubium Seeds and Its Enzymatic Action on Bovine Caseins. LWT Food Sci. Technol. 2010, 43, 759–764. [Google Scholar] [CrossRef]
- Piero, A.R.L.; Puglisi, I.; Petrone, G. Characterization of the Purified Actinidin as a Plant Coagulant of Bovine Milk. Eur. Food Res. Technol. 2011, 233, 517–524. [Google Scholar] [CrossRef]
- Jadhav, S.; Patil, V.N.; Naik, P.; Kadam, R.M. Studies on Chemical Quality of Ginger (Zingiber officinale L.) Milk Shake. Asian J. Dairy Food Res. 2018, 37, 13–17. [Google Scholar] [CrossRef]
- Luo, J.; Xiao, C.; Zhang, H.; Ren, F.; Lei, X.; Yang, Z.; Yu, Z. Characterization and Milk Coagulating Properties of Cynanchum otophyllum Schneid. proteases. J. Dairy Sci. 2018, 101, 2842–2850. [Google Scholar] [CrossRef]
- Drøhse, H.B.; Foltmann, B. Specificity of Milk-Clotting Enzymes towards Bovine κ-Casein. Biochim. Biophys. Acta BBA-Protein Struct. Mol. Enzym. 1989, 995, 221–224. [Google Scholar] [CrossRef]
- Zhang, Y.; Xia, Y.; Liu, X.; Xiong, Z.; Wang, S.; Zhang, N.; Ai, L. High-Level Expression and Substrate-Binding Region Modification of a Novel BL312 Milk-Clotting Enzyme to Enhance the Ratio of Milk-Clotting Activity to Proteolytic Activity. J. Agric. Food Chem. 2019, 67, 13684–13693. [Google Scholar] [CrossRef]
- Cavalli, S.V.; Lufrano, D.; Colombo, M.L.; Priolo, N. Properties and Applications of Phytepsins from Thistle Flowers. Phytochemistry 2013, 92, 16–32. [Google Scholar] [CrossRef]
- Rocha, G.F.; Cotabarren, J.; Obregón, W.D.; Fernández, G.; Rosso, A.M.; Parisi, M.G. Milk-Clotting and Hydrolytic Activities of an Aspartic Protease from Salpichroa Origanifolia Fruits on Individual Caseins. Int. J. Biol. Macromol. 2021, 192, 931–938. [Google Scholar] [CrossRef]
- Amira, A.B.; Besbes, S.; Attia, H.; Blecker, C. Milk-Clotting Properties of Plant Rennets and Their Enzymatic, Rheological, and Sensory Role in Cheese Making: A Review. Int. J. Food Prop. 2017, 20, S76–S93. [Google Scholar] [CrossRef] [Green Version]
- Mandujano-González, V.; Villa-Tanaca, L.; Anducho-Reyes, M.A.; Mercado-Flores, Y. Secreted Fungal Aspartic Proteases: A Review. Rev. Iberoam. Micol. 2016, 33, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Szecsi, P.B. The Aspartic Proteases. Scand. J. Clin. Lab. Investig. 1992, 52, 5–22. [Google Scholar] [CrossRef]
- Davies, D.R. The Structure and Function of the Aspartic Proteinases. Annu. Rev. Biophys. Biophys. Chem. 1990, 19, 189–215. [Google Scholar] [CrossRef] [PubMed]
- Yegin, S.; Fernandez-Lahore, M.; Salgado, A.J.G.; Guvenc, U.; Goksungur, Y.; Tari, C. Aspartic Proteinases from Mucor Spp. in Cheese Manufacturing. Appl. Microbiol. Biotechnol. 2011, 89, 949–960. [Google Scholar] [CrossRef] [Green Version]
- Rawlings, N.D.; Bateman, A. Pepsin Homologues in Bacteria. BMC Genom. 2009, 10, 437. [Google Scholar] [CrossRef] [Green Version]
- Nugent, P.G.; Albert, A.; Orprayoon, P.; Wilsher, J.; Pitts, J.E.; Blundell, T.L.; Dhanaraj, V. Protein Engineering Loops in Aspartic Proteinases: Site-Directed Mutagenesis, Biochemical Characterization and X-ray Analysis of Chymosin with a Replaced Loop from Rhizopus pepsin. Protein Eng. Des. Sel. 1996, 9, 885–893. [Google Scholar] [CrossRef] [Green Version]
- Salehi, M.; Aghamaali, M.R.; Sajedi, R.H.; Asghari, S.M.; Jorjani, E. Purification and Characterization of a Milk-Clotting Aspartic Protease from Withania Coagulans Fruit. Int. J. Biol. Macromol. 2017, 98, 847–854. [Google Scholar] [CrossRef]
- Lufrano, D.; Faro, R.; Castanheira, P.; Parisi, G.; Veríssimo, P.; Vairo-Cavalli, S.; Simões, I.; Faro, C. Molecular Cloning and Characterization of Procirsin, an Active Aspartic Protease Precursor from Cirsium vulgare (Asteraceae). Phytochemistry 2012, 81, 7–18. [Google Scholar] [CrossRef]
- Verissimo, P.; Esteves, C.; Faro, C.; Pires, E. The Vegetable Rennet of Cynara cardunculus L. Contains Two Proteinases with Chymosin and Pepsin-like Specificities. Biotechnol. Lett. 1995, 17, 621–626. [Google Scholar] [CrossRef]
- Sarmento, A.C.; Lopes, H.; Oliveira, C.S.; Vitorino, R.; Samyn, B.; Sergeant, K.; Debyser, G.; van Beeumen, J.; Domingues, P.; Amado, F.; et al. Multiplicity of Aspartic Proteinases from Cynara cardunculus L. Planta 2009, 230, 429–439. [Google Scholar] [CrossRef]
- Macedo, I.Q.; Faro, C.J.; Pires, E.M. Caseinolytic Specificity of Cardosin, an Aspartic Protease from the Cardoon Cynara cardunculus L.: Action on Bovine αs- and β-Casein and Comparison with Chymosin. J. Agric. Food Chem. 1996, 44, 42–47. [Google Scholar] [CrossRef] [Green Version]
- Sousa, M.J.; Malcata, F.X. Advances in the Role of a Plant Coagulant (Cynara cardunculus) in Vitro and during Ripening of Cheeses from Several Milk Species. Le Lait 2002, 82, 151–170. [Google Scholar] [CrossRef] [Green Version]
- Roseiro, L.B.; Barbosa, M.; Ames, J.M.; Wilbey, R.A. Cheesemaking with Vegetable Coagulants-the Use of Cynara L. for the Production of Ovine Milk Cheeses. Int. J. Dairy Technol. 2003, 56, 76–85. [Google Scholar] [CrossRef]
- Tito, F.R.; Pepe, A.; Tonon, C.V.; Daleo, G.R.; Guevara, M.G. Determination and Characterisation of Milk-Clotting Activity of Two Solanum tuberosum Aspartic Proteases (StAPs). Int. Dairy J. 2020, 104, 104645. [Google Scholar] [CrossRef]
- Faheem, M.; Martins-de-Sa, D.; Vidal, J.F.D.; Álvares, A.C.M.; Brandão-Neto, J.; Bird, L.E.; Tully, M.D.; von Delft, F.; Souto, B.M.; Quirino, B.F.; et al. Functional and Structural Characterization of a Novel Putative Cysteine Protease Cell Wall-Modifying Multi-Domain Enzyme Selected from a Microbial Metagenome. Sci. Rep. 2016, 6, 38031. [Google Scholar] [CrossRef]
- Cstorer, A.; Ménard, R. Catalytic Mechanism in Papain Family of Cysteine Peptidases. In Methods in Enzymology; Abelson, J., Simon, M., Verdine, G., Pyle, A., Eds.; Academic Press: Cambridge, UK, 1994; Volume 244, pp. 486–500. [Google Scholar]
- Coulombe, R.; Grochulski, P.; Sivaraman, J.; Ménard, R.; Mort, J.S.; Cygler, M. Structure of Human Procathepsin L Reveals the Molecular Basis of Inhibition by the Prosegment. EMBO J. 1996, 15, 5492–5503. [Google Scholar] [CrossRef]
- Konno, K.; Hirayama, C.; Nakamura, M.; Tateishi, K.; Tamura, Y.; Hattori, M.; Kohno, K. Papain Protects Papaya Trees from Herbivorous Insects: Role of Cysteine Proteases in Latex. Plant J. 2004, 37, 370–378. [Google Scholar] [CrossRef]
- Hashim, M.M.; Mingsheng, D.; Iqbal, M.F.; Xiaohong, C. Ginger Rhizome as a Potential Source of Milk Coagulating Cysteine Protease. Phytochemistry 2011, 72, 458–464. [Google Scholar] [CrossRef]
- Duarte, A.R.; Duarte, D.M.R.; Moreira, K.A.; Cavalcanti, M.T.H.; de Lima-Filho, J.L.; Porto, A.L.F. Jacaratia corumbensis O. Kuntze a New Vegetable Source for Milk-Clotting Enzymes. Braz. Arch. Biol. Technol. 2009, 52, 1–9. [Google Scholar] [CrossRef]
- Puglisi, I.; Petrone, G.; Piero, A.R.L. A Kiwi Juice Aqueous Solution as Coagulant of Bovine Milk and Its Potential in Mozzarella Cheese Manufacture. Food Bioprod. Process. 2014, 92, 67–72. [Google Scholar] [CrossRef]
- De Farias, V.A.; da Rocha Lima, A.D.; Costa, A.S.; de Freitas, C.D.T.; da Silva Araújo, I.M.; dos Santos Garruti, D.; de Figueiredo, E.A.T.; de Oliveira, H.D. Noni (Morinda citrifolia L.) Fruit as a New Source of Milk-Clotting Cysteine Proteases. Food Res. Int. 2020, 127, 108689. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Zhao, C.; Shi, Y.; Wei, G.; Yang, K.; Wang, X.; Huang, A. Proteomics Analysis of the Bio-Functions of Dregea sinensis Stems Provides Insights Regarding Milk-Clotting Enzyme. Food Res. Int. 2021, 144, 110340. [Google Scholar] [CrossRef] [PubMed]
- Serra, A.; Conte, G.; Corrales-Retana, L.; Casarosa, L.; Ciucci, F.; Mele, M. Nutraceutical and Technological Properties of Buffalo and Sheep Cheese Produced by the Addition of Kiwi Juice as a Coagulant. Foods 2020, 9, 637. [Google Scholar] [CrossRef]
- Mazorra-Manzano, M.A.; Perea-Gutiérrez, T.C.; Lugo-Sánchez, M.E.; Ramirez-Suarez, J.C.; Torres-Llanez, M.J.; González-Córdova, A.F.; Vallejo-Cordoba, B. Comparison of the Milk-Clotting Properties of Three Plant Extracts. Food Chem. 2013, 141, 1902–1907. [Google Scholar] [CrossRef] [PubMed]
- Fguiri, I.; Atigui, M.; Sboui, A.; Samira, A.; Marzougui, C.; Dbara, M.; Khorchani, T. Camel Milk-Clotting Using Plant Extracts as a Substitute to Commercial Rennet. J. Chem. 2021, 2021, 6680246. [Google Scholar] [CrossRef]
- Dunn, B.M. Overview of Pepsin-like Aspartic Peptidases. Curr. Protoc. Protein Sci. 2001, 25, 21–23. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J. Families of serine peptidases. In Methods in Enzymology; Abelson, J., Simon, M., Verdine, G., Pyle, A., Eds.; Academic Press: Cambridge, UK, 1994; Volume 244, pp. 19–61. [Google Scholar]
- Ahmed, I.A.M.; Morishima, I.; Babiker, E.E.; Mori, N. Characterisation of Partially Purified Milk-Clotting Enzyme from Solanum dubium Fresen Seeds. Food Chem. 2009, 116, 395–400. [Google Scholar] [CrossRef]
- Uchikoba, T.; Kaneda, M. Milk-Clotting Activity of Cucumisin, a Plant Serine Protease from Melon Fruit. Appl. Biochem. Biotechnol. 1996, 56, 325–330. [Google Scholar] [CrossRef]
- Kumari, M.; Sharma, A.; Jagannadham, M.V. Religiosin B, a Milk-Clotting Serine Protease from Ficus religiosa. Food Chem. 2012, 131, 1295–1303. [Google Scholar] [CrossRef]
- Tripathi, P.; Tomar, R.; Jagannadham, M.V. Purification and Biochemical Characterisation of a Novel Protease Streblin. Food Chem. 2011, 125, 1005–1012. [Google Scholar] [CrossRef]
- Bey, N.; Debbebi, H.; Abidi, F.; Marzouki, M.N.; Salah, A.B. The Non-Edible Parts of Fennel (Fœniculum vulgare) as a New Milk-Clotting Protease Source. Ind. Crops Prod. 2018, 112, 181–187. [Google Scholar] [CrossRef]
- González-Velázquez, D.A.; Mazorra-Manzano, M.A.; Martínez-Porchas, M.; Huerta-Ocampo, J.A.; Vallejo-Córdoba, B.; Mora-Cortes, W.G.; Moreno-Hernández, J.M.; Ramírez-Suarez, J.C. Exploring the Milk-Clotting and Proteolytic Activities in Different Tissues of Vallesia glabra: A New Source of Plant Proteolytic Enzymes. Appl. Biochem. Biotechnol. 2021, 193, 389–404. [Google Scholar] [CrossRef] [PubMed]
- Anusha, R.; Singh, M.K.; Bindhu, O.S. Characterisation of Potential Milk Coagulants from Calotropis gigantea Plant Parts and Their Hydrolytic Pattern of Bovine Casein. Eur. Food Res. Technol. 2014, 238, 997–1006. [Google Scholar] [CrossRef]
- Moreno-Hernández, J.M.; Hernández-Mancillas, X.D.; Navarrete, E.L.C.; Mazorra-Manzano, M.Á.; Osuna-Ruiz, I.; Rodríguez-Tirado, V.A.; Salazar-Leyva, J.A. Partial Characterization of the Proteolytic Properties of an Enzymatic Extract From “Aguama” Bromelia pinguin L. Fruit Grown in Mexico. Appl. Biochem. Biotechnol. 2017, 182, 181–196. [Google Scholar] [CrossRef]
- Beka, R.G.; Krier, F.; Botquin, M.; Guiama, V.D.; Donn, P.; Libouga, D.G.; Mbofung, C.M.; Dimitrov, K.; Slomianny, M.C.; Guillochon, D.; et al. Characterisation of a Milk-Clotting Extract from Balanites aegyptiaca Fruit Pulp. Int. Dairy J. 2014, 34, 25–31. [Google Scholar] [CrossRef]
- Cavalli, S.V.; Silva, S.V.; Cimino, C.; Malcata, F.X.; Priolo, N. Hydrolysis of Caprine and Ovine Milk Proteins, Brought about by Aspartic Peptidases from Silybum marianum Flowers. Food Chem. 2008, 106, 997–1003. [Google Scholar] [CrossRef]
- Llorente, B.E.; Obregón, W.D.; Avilés, F.X.; Caffini, N.O.; Vairo-Cavalli, S. Use of Artichoke (Cynara scolymus) Flower Extract as a Substitute for Bovine Rennet in the Manufacture of Gouda-Type Cheese: Characterization of Aspartic Proteases. Food Chem. 2014, 159, 55–63. [Google Scholar] [CrossRef]
- Chazarra, S.; Sidrach, L.; Lopez-Molina, D.; Rodríguez-López, J.N. Characterization of the milk-clotting properties of extracts from artichoke (Cynara scolymus, L.) flowers. Int. Dairy J. 2007, 17, 1393–1400. [Google Scholar] [CrossRef]
- Afsharnezhad, M.; Shahangian, S.S.; Sariri, R. A Novel Milk-Clotting Cysteine Protease from Ficus johannis: Purification and Characterization. Int. J. Biol. Macromol. 2019, 121, 173–182. [Google Scholar] [CrossRef]
- Gutiérrez-Méndez, N.; Balderrama-Carmona, A.; García-Sandoval, S.; Ramírez-Vigil, P.; Leal-Ramos, M.; García-Triana, A. Proteolysis and Rheological Properties of Cream Cheese Made with a Plant-Derived Coagulant from Solanum elaeagnifolium. Foods 2019, 8, 44. [Google Scholar] [CrossRef] [Green Version]
- Folgado, A.; Serra, A.T.; Prazeres, I.; Bento-Silva, A.; Bronze, M.R.; Abranches, R. Hairy Root Cultures of Cynara cardunculus L. as a Valuable Source of Hydroxycinnamic Acid Compounds. PCTOC 2021, 147, 37–47. [Google Scholar] [CrossRef]
- Vairo-Cavalli, S.; Claver, S.; Priolo, N.; Natalucci, C. Extraction and Partial Characterization of a Coagulant Preparation from Silybum marianum Flowers. Its Action on Bovine Caseinate. J. Dairy Res. 2005, 72, 271–275. [Google Scholar] [CrossRef]
- Llorente, B.E.; Brutti, C.B.; Caffini, N.O. Purification and Characterization of a Milk-Clotting Aspartic Proteinase from Globe Artichoke (Cynara scolymus L.). J. Agric. Food Chem. 2004, 52, 8182–8189. [Google Scholar] [CrossRef]
- Meng, F.; Zhao, H.; Lu, F.; Bie, X.; Lu, Z.; Lu, Y. Novel Bacillus Milk-Clotting Enzyme Produces Diverse Functional Peptides in Semihard Cheese. J. Agric. Food Chem. 2021, 69, 2784–2792. [Google Scholar] [CrossRef]
- Visser, S. Proteolytic Enzymes and Their Relation to Cheese Ripening and Flavor: An Overview. J. Dairy Sci. 1993, 76, 329–350. [Google Scholar] [CrossRef]
- Saha, B.C.; Hayashi, K. Debittering of Protein Hydrolyzates. Biotechnol. Adv. 2001, 19, 355–370. [Google Scholar] [CrossRef]
- Ney, K.H. Bitterness of peptides: Amino acid composition and chain length. Food Tast. Chem. 1979, 115, 149–173. [Google Scholar]
- Wieser, H.; Belitz, H.-D. Zusammenhänge zwischen Struktur und Bittergeschmack bei Aminosäuren und Peptiden. Z. Lebensm. Unters. Forsch. 1975, 159, 65–72. [Google Scholar] [CrossRef]
- Lee, K.D.; Lo, C.G.; Warthesen, J.J. Removal of Bitterness from the Bitter Peptides Extracted from Cheddar Cheese with Peptidases from Lactococcus lactis ssp. cremoris SK11. J. Dairy Sci. 1996, 79, 1521–1528. [Google Scholar] [CrossRef]
- Tchorbanov, B.; Marinova, M.; Grozeva, L. Debittering of Protein Hydrolysates by Lactobacillus LBL-4 Aminopeptidase. Enzym. Res. 2011, 2011, 538676. [Google Scholar] [CrossRef] [Green Version]
- Azarnia, S.; Robert, N.; Lee, B. Biotechnological Methods to Accelerate Cheddar Cheese Ripening. Crit. Rev. Biotechnol. 2006, 26, 121–143. [Google Scholar] [CrossRef] [PubMed]
- Toelstede, S.; Hofmann, T. Sensomics Mapping and Identification of the Key Bitter Metabolites in Gouda Cheese. J. Agric. Food Chem. 2008, 56, 2795–2804. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.K.; Young, N.D.; Drake, M.; Cadwallader, K.R. Production and Sensory Characterization of a Bitter Peptide from β-Casein. J. Agric. Food Chem. 2005, 53, 1185–1189. [Google Scholar] [CrossRef] [PubMed]
- Hussein, J.B.; Suleiman, A.; Ilesanmi, J.Y.O.; Sanusi, S.A. Chemical Composition and Sensory Qualities of West African Soft Cheese (Warankashi) Produced from Blends of Cow Milk and Soy Milk. Niger. J. Trop. Agric. 2016, 16, 79–89. [Google Scholar]
- Fernández-Salguero, J.; Sanjuán, E. Influence of Vegetable and Animal Rennet on Proteolysis during Ripening in Ewes’ Milk Cheese. Food Chem. 1999, 64, 177–183. [Google Scholar] [CrossRef]
- Lemieux, L.; Simard, R.E. Bitter flavour in dairy products. I. A Review of the Factors Likely to Influence its Development, Mainly in Cheese Manufacture. Le Lait 1991, 71, 599–636. [Google Scholar] [CrossRef] [Green Version]
- Rana, M.; Hoque, M.; Rahman, M.; Habib, R.; Siddiki, M. Papaya (Carica papaya) Latex—An Alternative to Rennet for Cottage Cheese Preparation. J. Adv. Vet. Anim. Res. 2017, 4, 249. [Google Scholar] [CrossRef]
- Marino, M.; de Wittenau, G.D.; Saccà, E.; Cattonaro, F.; Spadotto, A.; Innocente, N.; Radovic, S.; Piasentier, E.; Marroni, F. Metagenomic Profiles of Different Types of Italian High-Moisture Mozzarella Cheese. Food Microbiol. 2019, 79, 123–131. [Google Scholar] [CrossRef]
- Crespo, A.; Martín, A.; Ruiz-Moyano, S.; Benito, M.J.; Rufo, M.; Paniagua, J.M.; Jiménez, A. Application of Ultrasound for Quality Control of Torta Del Casar Cheese Ripening. J. Dairy Sci. 2020, 103, 8808–8821. [Google Scholar] [CrossRef]
- Hatta, W.; Sudarwanto, M.; Sudirman, I.; Malaka, R. Prevalence and Sources of Contamination of Escherichia coli and Salmonella spp. in Cow Milk Dangke, Indonesian Fresh Soft Cheese. Glob. Vet. 2013, 11, 352–356. [Google Scholar]
- Darwish, M. Influence of Plant Based Coagulant (Enzyme Extracts from Albizia and Sunflower Seeds) on Quality of Domiati Cheese. J. Food Dairy Sci. 2016, 7, 501–506. [Google Scholar] [CrossRef]
- Ferreira, I.M.P.L.V.O.; Pinho, O.; Sampaio, P. Volatile Fraction of DOP “Castelo Branco” Cheese: Influence of Breed. Food Chem. 2009, 112, 1053–1059. [Google Scholar] [CrossRef]
- Guiné, R.P.F.; Fontes, L.; Lima, M.J. Evaluation of Texture in Serra Da Estrela Cheese Manufactured in Different Dairies. Open Agric. 2019, 4, 475–486. [Google Scholar] [CrossRef]
- Araújo-Rodrigues, H.; Tavaria, F.K.; dos Santos, M.T.P.G.; Alvarenga, N.; Pintado, M.M. A Review on Microbiological and Technological Aspects of Serpa PDO Cheese: An Ovine Raw Milk Cheese. Int. Dairy J. 2020, 100, 104561. [Google Scholar] [CrossRef]
- Cardinali, F.; Ferrocino, I.; Milanović, V.; Belleggia, L.; Corvaglia, M.R.; Garofalo, C.; Foligni, R.; Mannozzi, C.; Mozzon, M.; Cocolin, L. Microbial Communities and Volatile Profile of Queijo de Azeitão PDO Cheese, a Traditional Mediterranean Thistle-Curdled Cheese from Portugal. Food Res. Int. 2021, 147, 110537. [Google Scholar] [CrossRef]
- Carbonell, M.; Nuñez, M.; Fernández-García, E. Evolution of the Volatile Components of Ewe Raw Milk La Serena Cheese during Ripening. Correlation with Flavour Characteristics. Le Lait 2002, 82, 683–698. [Google Scholar] [CrossRef] [Green Version]
- Cardinali, F.; Taccari, M.; Milanović, V.; Osimani, A.; Polverigiani, S.; Garofalo, C.; Foligni, R.; Mozzon, M.; Zitti, S.; Raffaelli, N. Yeast and Mould Dynamics in Caciofiore della Sibilla Cheese Coagulated with an Aqueous Extract of Carlina acanthifolia All. Yeast 2016, 33, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Carrascosa, C.; Millán, R.; Saavedra, P.; Raposo, A.; Saraiva, A.; Sanjuán, E. A Study of Vegetable (Thistle) Rennet in the Production of Flor de Guía Cheese. Biomed. Biopharm. Res. J. 2020, 17, 1–13. [Google Scholar] [CrossRef]
- Martínez-Ruiz, N.D.R.; Enriquez, S.F.; Vázquez-Nájera, R.E.; López-Díaz, J.A. Microbiological Quality of Asadero Cheese Manufactured with a Plant Based Coagulant from Solanum elaeagnifolium. Nutr. Food Sci. 2013, 4, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Sanjuán, E.; Millán, R.; Saavedra, P.; Carmona, M.A.; Gómez, R.; Fernández-Salguero, J. Influence of Animal and Vegetable Rennet on the Physicochemical Characteristics of Los Pedroches Cheese during Ripening. Food Chem. 2002, 78, 281–289. [Google Scholar] [CrossRef]
- Dias, J.M.; Lage, P.; Alvarenga, N.; Garcia, J.; Borrega, J.; Santos, M.T.; Lampreia, C.; Coelho, L.; Pássaro, J.; Martins, J.; et al. Impact of Environmental Conditions on the Ripening of Queijo de Évora PDO Cheese. J. Food Sci. Technol. 2021, 58, 3942–3952. [Google Scholar] [CrossRef] [PubMed]
- Dias, J.; Lage, P.; Garrido, A.; Machado, E.; Conceição, C.; Gomes, S.; Martins, A.; Paulino, A.; Duarte, M.F.; Alvarenga, N. Evaluation of Gas Holes in “Queijo de Nisa” PDO Cheese Using Computer Vision. J. Food Sci. Technol. 2021, 58, 1072–1080. [Google Scholar] [CrossRef]
- Beigomi, M.; Mohammadifar, M.A.; Hashemi, M.; Rohani, M.G.; Senthil, K.; Valizadeh, M. Biochemical and Rheological Characterization of a Protease from Fruits of Withania coagulans with a Milk-Clotting Activity. Food Sci. Biotechnol. 2014, 23, 1805–1813. [Google Scholar] [CrossRef]
- Tejada, L.; Abellán, A.; Cayuela, J.M.; Martínez-Cacha, A.; Fernández-Salguero, J. Proteolysis in Goats’ Milk Cheese Made with Calf Rennet and Plant Coagulant. Int. Dairy J. 2008, 18, 139–146. [Google Scholar] [CrossRef]
- Colombo, M.L.; Fernández, A.; Cimino, C.V.; Liggieri, C.; Bruno, M.; Faro, C.; Veríssimo, P.C.; Vairo-Cavalli, S. Miniature Cheeses Made with Blends of Chymosin and a Vegetable Rennet from Flowers of Silybum marianum: Enzymatic Characterization of the Flower-Coagulant Peptidase. Food Chem. 2018, 266, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Bruno, M.A.; Lazza, C.M.; Errasti, M.E.; López, L.M.I.; Caffini, N.O.; Pardo, M.F. Milk Clotting and Proteolytic Activity of an Enzyme Preparation from Bromelia hieronymi Fruits. Food Sci. Technol. 2010, 43, 695–701. [Google Scholar] [CrossRef]
- Barracosa, P.; Simões, I.; Martins, A.P.; Barros, M.; Pires, E. Biochemical Diversity of Cardoon Flowers (Cynara cardunculus L.): Predicting PDO Mediterranean Cheese Textures. Food Biosci. 2021, 39, 100805. [Google Scholar] [CrossRef]
- Folgado, A.; Abranches, R. Plant Aspartic Proteases for Industrial Applications: Thistle Get Better. Plants 2020, 9, 147. [Google Scholar] [CrossRef] [Green Version]
- Correia, P.; Vítor, A.; Tenreiro, M.; Correia, A.C.; Madanelo, J.; Guiné, R. Effect of Different Thistle Flower Ecotypes as Milk-clotting in Serra da Estrela Cheese. Nutr. Food Sci. 2016, 46, 458–475. [Google Scholar] [CrossRef]
- Christensen, J.E.; Dudley, E.G.; Pederson, J.A.; Steele, J.L. Christensen. Peptidases and Amino Acid Catabolism in Lactic Acid Bacteria. Antonie Van Leeuwenhoek 1999, 76, 217–246. [Google Scholar] [CrossRef]
- Navidghasemizad, S.; Takala, T.M.; Alatossava, T.; Saris, P.E. Proline Iminopeptidase PepI Overexpressing Lactobacillus casei as an Adjunct Starter in Edam Cheese. Bioengineered 2013, 4, 408–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agboola, S. Cheesmaking from Ultrafiltered Milk Using Plant Rennet. Aust. J. Dairy Technol. 2002, 57, 143. [Google Scholar]
- Katsaros, G.I.; Tavantzis, G.; Taoukis, P.S. Production of Novel Dairy Products Using Actinidin and High Pressure as Enzyme Activity Regulator. Innov. Food Sci. Emerg. Technol. 2010, 11, 47–51. [Google Scholar] [CrossRef]
- McSweeney, P.L.H. Cheese Problems Solved; Elsevier: Amsterdam, The Netherlands, 2007; ISBN 9781845690601. [Google Scholar]

| Plant Extract | MCA | PA | MCA/PA | Reference |
|---|---|---|---|---|
| Calotropis gigantea | 450 (U/mL) | 86.45 (U/mL) | 5.21 | [68] |
| Zingiber officinale | 314 (unit/mg) | 0.19 (unit/mg) | 1653.00 | [52] |
| Silybum marianum | 0.083 (RU/mL) | 0.128 (EA/mL) | 0.65 | [77] |
| Balanites aegyptiaca | 2.43 (MCU/mL) | 4.96 (MCU/mL) | 0.49 | [70] |
| Cynara scolymus | 147.65 (MCU/mg) | 5.45 (Ucas/mg) | 27.1 | [78] |
| Bromelia pinguin | 2.59 (U/mg) | 2.0 (U/mg) | 1.29 | [69] |
| Morinda citrifolia | 238.8 (U/mL) | 8.86 (U/mg) | 27.00 | [55] |
| Vallesia glabra | 0.20 (U/mL) | 19.04 (U/mL) | 1.00 | [67] |
| Ficus johannis | 21.88 (U/mL) | 0.339 (IU/mL) | 64.54 | [74] |
| Solanum elaeagnifolium | 4347.00 (U/mL) | 1.3 (U-Gly/mg) | 3343.00 | [75] |
| Actinidia chinensis | 2.7 (U/mg) | 0.55 (U/mg) | 5.00 | [58] |
| Animal/microbial enzymes | ||||
| Calf chymosin | 551.00 (SU/mg) | 2.28 (U/mg) | 243.20 | [79] |
| R. miehei | 756.00 (SU/mg) | 14.74 (U/mg) | 51.31 | [79] |
| Origin | Peptide | Q-Value | Reference |
|---|---|---|---|
| αS1-CN (f11–14) | Leu-Pro-Gln-Glu | 1367 | [84] |
| αS1-CN (f1–7) | Arg-Pro-Lys-His-Pro-Ile-Lys | 1771 | [84] |
| αS1-CN (f191–197) | Lys-Pro-Trp-Ile-Gln-Pro-Lys | 2010 | [84] |
| β-CN (f73–76) | Ile-Pro-Pro-Leu | 2658 | [87] |
| β-CN (f60–68) | Tyr-Pro-Phe-Pro-Gly-Pro-Ile-His-Asn | 1871 | [87] |
| β-CN (f8–16) | Val-Pro-Gly-Glu-Ile-Val-Glu-Ser-Leu | 1390 | [84] |
| β-CN (f200–206) | Val-Arg-Gly-Pro-Phe-Pro | 1718 | [87] |
| β-CN (f193–209) | Tyr-Gln-Glu-Pro-Val-Leu-Gly-Pro-Val-Arg-Gly-Pro-Phe-Pro-Ile-Ile-Val | 1839 | [88] |
| Cheese Type | Name | Milk Type | Plant Coagulant Source | Reference |
|---|---|---|---|---|
| Soft | Torta del Casar | Ewe | Cynara cardunculus | [94] |
| Dangke | Buffalo | Carica papaya | [95] | |
| Domiati | Buffalo | Heliantus hannuus | [96] | |
| Warankashi | Cow and Soymilk | Calotropis procera or Carica papaya | [89] | |
| Semi-soft | Castelo Branco, Serra da Estrela, Serpa, Aizeitão, La Serena, Caciofiore dei Sibillini | Ewe | Cynara cardunculus | [97,98,99,100,101,102] |
| Flor de Guía, Mestiço de Tolosa | Ewe and Goat | Cynara cardunculus | [22,103] | |
| Asadero | Cow | Solanum elaeagnifolium | [104] | |
| Semi-hard | Los Pedroches, Évora Nisa, | Ewe | Cynara cardunculus | [105,106,107] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nicosia, F.D.; Puglisi, I.; Pino, A.; Caggia, C.; Randazzo, C.L. Plant Milk-Clotting Enzymes for Cheesemaking. Foods 2022, 11, 871. https://doi.org/10.3390/foods11060871
Nicosia FD, Puglisi I, Pino A, Caggia C, Randazzo CL. Plant Milk-Clotting Enzymes for Cheesemaking. Foods. 2022; 11(6):871. https://doi.org/10.3390/foods11060871
Chicago/Turabian StyleNicosia, Fabrizio Domenico, Ivana Puglisi, Alessandra Pino, Cinzia Caggia, and Cinzia Lucia Randazzo. 2022. "Plant Milk-Clotting Enzymes for Cheesemaking" Foods 11, no. 6: 871. https://doi.org/10.3390/foods11060871
APA StyleNicosia, F. D., Puglisi, I., Pino, A., Caggia, C., & Randazzo, C. L. (2022). Plant Milk-Clotting Enzymes for Cheesemaking. Foods, 11(6), 871. https://doi.org/10.3390/foods11060871