

Phenolic Content, Amino Acids, Volatile Compounds, Antioxidant Capacity, and Their Relationship in Wild Garlic (A. ursinum L.)
 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Plant Material
2.3. Total Phenolic Content and Total Antioxidant Capacity
2.4. Amino Acids
2.5. Volatile Organic Compounds
2.6. Statistical Analysis
3. Results
3.1. Relationships among Total Phenolic Content, Amino Acids, Volatile Compounds, and Antioxidant Capacity in Both Leaves and Bulbs of Wild Garlic
3.2. Phytochemical Biodiversity of Wild Garlic Populations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sobolewska, D.; Podolak, I.; Makowska-Wąs, J. Allium Ursinum: Botanical, Phytochemical and Pharmacological Overview. Phytochem. Rev. 2015, 14, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Tomšik, A.; Pavlić, B.; Vladić, J.; Ramić, M.; Brindza, J.; Vidović, S. Optimization of Ultrasound-Assisted Extraction of Bioactive Compounds from Wild Garlic (Allium ursinum L.). Ultrason. Sonochem. 2016, 29, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, B.; Schulz, H.; Storsberg, J.; Keusgen, M. Chemical Characterization of Allium ursinum L. Depending on Harvesting Time. J. Agric. Food Chem. 2005, 53, 7288–7294. [Google Scholar] [CrossRef] [PubMed]
- Sendl, A. Allium sativum and Allium ursinum: Part 1 Chemistry, Analysis, History, Botany. Phytomedicine 1995, 1, 323–339. [Google Scholar] [CrossRef] [PubMed]
- Radulović, N.S.; Miltojević, A.B.; Stojković, M.B.; Blagojević, P.D. New Volatile Sulfur-Containing Compounds from Wild Garlic (Allium ursinum L., Liliaceae). Food Res. Int. 2015, 78, 1–10. [Google Scholar] [CrossRef]
- Gîtin, L.; Dinicə, R.; Neagu, C.; Dumitrascu, L. Sulfur Compounds Identification and Quantification from Allium spp. Fresh Leaves. J. Food Drug Anal. 2014, 22, 425–430. [Google Scholar] [CrossRef]
- Maccelli, A.; Cesa, S.; Cairone, F.; Secci, D.; Menghini, L.; Chiavarino, B.; Fornarini, S.; Crestoni, M.E.; Locatelli, M. Metabolic Profiling of Different Wild and Cultivated Allium Species Based on High-Resolution Mass Spectrometry, High-Performance Liquid Chromatography-Photodiode Array Detector, and Color Analysis. J. Mass Spectrom. 2020, 55, e4525. [Google Scholar] [CrossRef]
- Métayer, S.; Seiliez, I.; Collin, A.; Duchêne, S.; Mercier, Y.; Geraert, P.A.; Tesseraud, S. Mechanisms through Which Sulfur Amino Acids Control Protein Metabolism and Oxidative Status. J. Nutr. Biochem. 2008, 19, 207–215. [Google Scholar] [CrossRef]
- Tóth, T.; Kovarovič, J.; Bystrická, J.; Vollmannová, A.; Musilová, J.; Lenková, M. The Content of Polyphenols and Antioxidant Activity in Leaves and Flowers of Wild Garlic (Allium ursinum L.). Acta Aliment. 2018, 47, 252–258. [Google Scholar] [CrossRef]
- Bhat, R. Bioactive Compounds of Allium Species. In Bioactive Compounds in Underutilized Vegetables and Legumes; Hosakatte Niranjana Murthy, K.Y.P., Ed.; Springer Publishing: New York, NY, USA, 2020; pp. 277–295. [Google Scholar]
- Kovarovič, J.; Bystrická, J.; Urminská, D.; Harangozo, L.; Vollmannová, A.; Trebichalský, P.; Timoracká, M.; Carbonell-Barrachina, A.A. Evaluation and Comparison of Total Polyphenols Content and Antioxidant Activity of Wild Garlic (Allium ursinum L.) in Selected Morphological Parts. J. Microbiol. Biotechnol. Food Sci. 2019, 9, 492–495. [Google Scholar] [CrossRef]
- Luta, G.; Gherghina, E.; Balan, D.; Israel-Roming, F. Bioactive Compounds and Antioxidant Properties of Some Wild Plants with Potential Culinary Uses. Rev. Chim. 2020, 71, 179–184. [Google Scholar] [CrossRef]
- Leporatti, M.L.; Ivancheva, S. Preliminary Comparative Analysis of Medicinal Plants Used in the Traditional Medicine of Bulgaria and Italy. J. Ethnopharmacol. 2003, 87, 123–142. [Google Scholar] [CrossRef] [PubMed]
- Błażewicz-Woźniak, M.; Michowska, A. The Growth, Flowering and Chemical Composition of Leaves of Three Ecotypes of Allium ursinum L. Acta Agrobot. 2011, 64, 320. [Google Scholar] [CrossRef]
- Benkeblia, N.; Lanzotti, V. Allium Thiosulfinates: Chemistry, Biological Properties and Their Potential Utilization in Food Preservation. Food 2007, 1, 193–201. [Google Scholar]
- Pavlović, D.R.; Veljković, M.; Stojanović, N.M.; Gočmanac-Ignjatović, M.; Mihailov-Krstev, T.; Branković, S.; Sokolović, D.; Marčetić, M.; Radulović, N.; Radenković, M. Influence of Different Wild-Garlic (Allium ursinum) Extracts on the Gastrointestinal System: Spasmolytic, Antimicrobial and Antioxidant Properties. J. Pharm. Pharmacol. 2017, 69, 1208–1218. [Google Scholar] [CrossRef]
- Jivishov, E.; Keusgen, M. Can Allium Chemical Chest Be a Source of Anticancer Compounds? Phytochem. Rev. 2020, 19, 1503–1523. [Google Scholar] [CrossRef]
- Kamenetsky, R.; Khassanov, F.; Rabinowitch, H.D.; Auger, J. Garlic Biodiversity and Genetic Resources. Med. Aromat. Plant Sci. Biotechnol. 2007, 1, 1–5. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Ou, B.; Chang, T.; Huang, D.; Prior, R.L. Determination of Total Antioxidant Capacity by Oxygen Radical Absorbance Capacity (ORAC) Using Fluorescein as the Fluorescence Probe: First Action 2012.23. J. AOAC Int. 2013, 96, 1372–1376. [Google Scholar] [CrossRef] [PubMed]
- Buha, S.M.; Panchal, A.; Panchal, H.; Chambhare, R.; Kumar, S.; Jain, M.; Patel, P.R. HPLC-FLD for the Simultaneous Determination of Primary and Secondary Amino Acids from Complex Biological Sample by Pre-Column Derivatization. J. Chromatogr. Sci. 2011, 49, 118–123. [Google Scholar] [CrossRef]
- Molina-Calle, M.; Priego-Capote, F.; de Castro, M.D.L. HS–GC/MS Volatile Profile of Different Varieties of Garlic and Their Behavior under Heating. Anal. Bioanal. Chem. 2016, 408, 3843–3852. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Chong, J.; Zhou, G.; De Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the Gap between Raw Spectra and Functional Insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]
- McKey, D. Adaptive Patterns in Alkaloid Physiology. Am. Nat. 1974, 108, 305–320. [Google Scholar] [CrossRef]
- McCall, A.C.; Fordyce, J.A. Can Optimal Defence Theory Be Used to Predict the Distribution of Plant Chemical Defences? J. Ecol. 2010, 98, 985–992. [Google Scholar] [CrossRef]
- Mallor, C.; Thomas, B. Resource Allocation and the Origin of Flavour Precursors in Onion Bulbs. J. Hortic. Sci. Biotechnol. 2008, 83, 191–198. [Google Scholar] [CrossRef]
- Suzuki, T.; Sugii, M.; Kakimoto, T. Metabolic Incorporation of L-Valine-[14C] into S-(2-Carboxypropyl) Glutathione and S-(2-Carboxypropyl) Cysteine in Garlic. Chem. Pharm. Bull. 1962, 10, 328–331. [Google Scholar] [CrossRef]
- Suzuki, T.; Sugii, M.; Kakimoto, T. New γ-Glutamyl Peptides in Garlic. Chem. Pharm. Bull. 1961, 9, 77–78. [Google Scholar] [CrossRef]
- Yoshimoto, N.; Saito, K. S-Alk(En)Ylcysteine Sulfoxides in the Genus Allium: Proposed Biosynthesis, Chemical Conversion, and Bioactivities. J. Exp. Bot. 2019, 70, 4123–4137. [Google Scholar] [CrossRef]
- Lancaster, J.E.; Shaw, M.L. Metabolism of γ-Glutamyl Peptides during Development, Storage and Sprouting of Onion Bulbs. Phytochemistry 1991, 30, 2857–2859. [Google Scholar] [CrossRef]
- Lancaster, J.E.; Shaw, M.L. γ-Glutamyl Peptides in the Biosynthesis of S-Alk(En)Yl-l-Cysteine Sulphoxides (Flavour Precursors) in Allium. Phytochemistry 1989, 28, 455–460. [Google Scholar] [CrossRef]
- Shimon, L.J.W.; Rabinkov, A.; Shin, I.; Miron, T.; Mirelman, D.; Wilchek, M.; Frolow, F. Two Structures of Alliinase from Alliium sativum L.: Apo Form and Ternary Complex with Aminoacrylate Reaction Intermediate Covalently Bound to the PLP Cofactor. J. Mol. Biol. 2007, 366, 611–625. [Google Scholar] [CrossRef] [PubMed]
- Stanisavljević, N.; Soković Bajić, S.; Jovanović, Ž.; Matić, I.; Tolinački, M.; Popović, D.; Popović, N.; Terzić-Vidojević, A.; Golić, N.; Beškoski, V.; et al. Antioxidant and Antiproliferative Activity of Allium ursinum and Their Associated Microbiota during Simulated in Vitro Digestion in the Presence of Food Matrix. Front. Microbiol. 2020, 11, 601616. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.W.; Hang, L.F.; Ali, S.; Xu, X.Y.; Liu, Y.J.; Yan, Q.Q.; Luo, Q.Y.; Li, Y.; Lin, L.J.; Li, H.X.; et al. Biosynthesis and Metabolism of Garlic Odor Compounds in Cultivated Chinese Chives (Allium tuberosum) and Wild Chinese Chives (Allium hookeri). Int. J. Mol. Sci. 2022, 23, 7013. [Google Scholar] [CrossRef] [PubMed]
- Santos-Sánchez, N.F.; Salas-Coronado, R.; Hernández-Carlos, B.; Villanueva-Cañongo, C.; Santos-Sánchez, N.F.; Salas-Coronado, R.; Hernández-Carlos, B.; Villanueva-Cañongo, C. Shikimic Acid Pathway in Biosynthesis of Phenolic Compounds. Plant Physiol. Asp. Phenolic Compd. 2019, 1, 1–15. [Google Scholar] [CrossRef]
- Xu, N.; Chen, G.; Liu, H. Antioxidative Categorization of Twenty Amino Acids Based on Experimental Evaluation. Molecules 2017, 22, 2066. [Google Scholar] [CrossRef]
- Tzin, V.; Galili, G. New Insights into the Shikimate and Aromatic Amino Acids Biosynthesis Pathways in Plants. Mol. Plant 2010, 3, 956–972. [Google Scholar] [CrossRef]
- Al Jitan, S.; Alkhoori, S.A.; Yousef, L.F. Phenolic Acids From Plants: Extraction and Application to Human Health. Stud. Nat. Prod. Chem. 2018, 58, 389–417. [Google Scholar] [CrossRef]
- Jones, D.A.; Jones, J.D.G. The Role of Leucine-Rich Repeat Proteins in Plant Defences. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 1997; Volume 24, pp. 89–167. [Google Scholar] [CrossRef]
- Li, Y.; Li, S.; Du, R.; Wang, J.; Li, H.; Xie, D.; Yan, J. Isoleucine Enhances Plant Resistance Against Botrytis Cinerea via Jasmonate Signaling Pathway. Front. Plant Sci. 2021, 12, 1738. [Google Scholar] [CrossRef]
- Muthuramalingam, P.; Krishnan, S.R.; Pandian, S.; Mareeswaran, N.; Aruni, W.; Pandian, S.K.; Ramesh, M. Global Analysis of Threonine Metabolism Genes Unravel Key Players in Rice to Improve the Abiotic Stress Tolerance. Sci. Rep. 2018, 8, 9270. [Google Scholar] [CrossRef] [PubMed]
- Atmaca, G. Antioxidant Effects of Sulfur-Containing Amino Acids. Yonsei Med. J. 2009, 45, 776–788. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, B.L.; Edrada-Ebel, R.; Da Costa, F.B. Effect of the Environment on the Secondary Metabolic Profile of Tithonia Diversifolia: A Model for Environmental Metabolomics of Plants. Sci. Rep. 2016, 6, 29265. [Google Scholar] [CrossRef] [PubMed]
- Chapin, F.S.; Bloom, A.J.; Field, C.B.; Waring, R.H. Plant Responses to Multiple Environmental Factors. Bioscience 1987, 37, 49–57. [Google Scholar] [CrossRef]
- Lachowicz, S.; Kolniak-Ostek, J.; Oszmiański, J.; Wiśniewski, R. Comparison of Phenolic Content and Antioxidant Capacity of Bear Garlic (Allium ursinum L.) in Different Maturity Stages. J. Food Process. Preserv. 2017, 41, e12921. [Google Scholar] [CrossRef]
- Heimler, D.; Romani, A.; Ieri, F. Plant Polyphenol Content, Soil Fertilization and Agricultural Management: A Review. Eur. Food Res. Technol. 2017, 243, 1107–1115. [Google Scholar] [CrossRef]
- Mahmutović, O.; Tahirović, I.; Copra, A.; Memić, M.; Ibragić, S.; Karić, L. Correlation of Total Secondary Sulfur Compounds, Total Phenols and Antioxidant Capacity in the Ramsons and Garlic. Br. J. Pharm. Res. 2014, 4, 2662–2669. [Google Scholar] [CrossRef]
- Mihaylova, D.S.; Lante, A.; Tinello, F.; Krastanov, A.I. Study on the Antioxidant and Antimicrobial Activities of Allium ursinum L. Pressurised-Liquid Extract. Nat. Prod. Res. 2014, 28, 2000–2005. [Google Scholar] [CrossRef]
- Lachowicz, S.; Oszmiański, J.; Wiśniewski, R. Determination of Triterpenoids, Carotenoids, Chlorophylls, and Antioxidant Capacity in Allium ursinum L. at Different Times of Harvesting and Anatomical Parts. Eur. Food Res. Technol. 2018, 244, 1269–1280. [Google Scholar] [CrossRef]
- Vuković, S.; Popović-Djordjević, J.B.; Kostić, A.; Pantelić, N.D.; Srećković, N.; Akram, M.; Laila, U.; Katanić Stanković, J.S. Allium Species in the Balkan Region—Major Metabolites, Antioxidant and Antimicrobial Properties. Horticulturae 2023, 9, 408. [Google Scholar] [CrossRef]
- Jones, M.G.; Hughes, J.; Tregova, A.; Milne, J.; Tomsett, A.B.; Collin, H.A. Biosynthesis of the Flavour Precursors of Onion and Garlic. J. Exp. Bot. 2004, 55, 1903–1918. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, A.; Mikhova, B.; Najdenski, H.; Tsvetkova, I.; Kostova, I. Chemical Composition and Antimicrobial Activity of Wild Garlic Allium ursinum of Bulgarian Origin. Nat. Prod. Commun. 2009, 4, 1059–1062. [Google Scholar] [CrossRef]
- Scheffler, L.; Sharapa, C.; Amar, T.; Buettner, A. Identification and Quantification of Volatile Ramson-Derived Metabolites in Humans. Front. Chem. 2018, 6, 410. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Harnly, J.M. Free Amino Acid and Cysteine Sulfoxide Composition of 11 Garlic (Allium sativum L.) Cultivars by Gas Chromatography with Flame Ionization and Mass Selective Detection. J. Agric. Food Chem. 2005, 53, 9100–9104. [Google Scholar] [CrossRef] [PubMed]
- Krivokapić, M.; Bradić, J.; Petković, A.; Popović, M. Phytochemical and Pharmacological Properties of Allium ursinum. Serb. J. Exp. Clin. Res. 2018, 22, 357–362. [Google Scholar] [CrossRef]
- Fekkes, D. State-of-the-Art of High-Performance Liquid Chromatographic Analysis of Amino Acids in Physiological Samples. J. Chromatogr. B Biomed. Sci. Appl. 1996, 682, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Vranova, V.; Zahradnickova, H.; Janous, D.; Skene, K.R.; Matharu, A.S.; Rejsek, K.; Formanek, P. The Significance of D-Amino Acids in Soil, Fate and Utilization by Microbes and Plants: Review and Identification of Knowledge Gaps. Plant Soil 2012, 354, 21–39. [Google Scholar] [CrossRef]
- Raab, T.K.; Lipson, D.A.; Monson, R.K. Soil amino acid utilization among species of the cyperaceae: Plant and soil processes. Ecology 1999, 80, 2408–2419. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of Proline under Changing Environments. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Reuter, H.D. Allium sativum and Allium ursinum: Part 2 Pharmacology and Medicinal Application. Phytomedicine 1995, 2, 73–91. [Google Scholar] [CrossRef]




{kind=link}
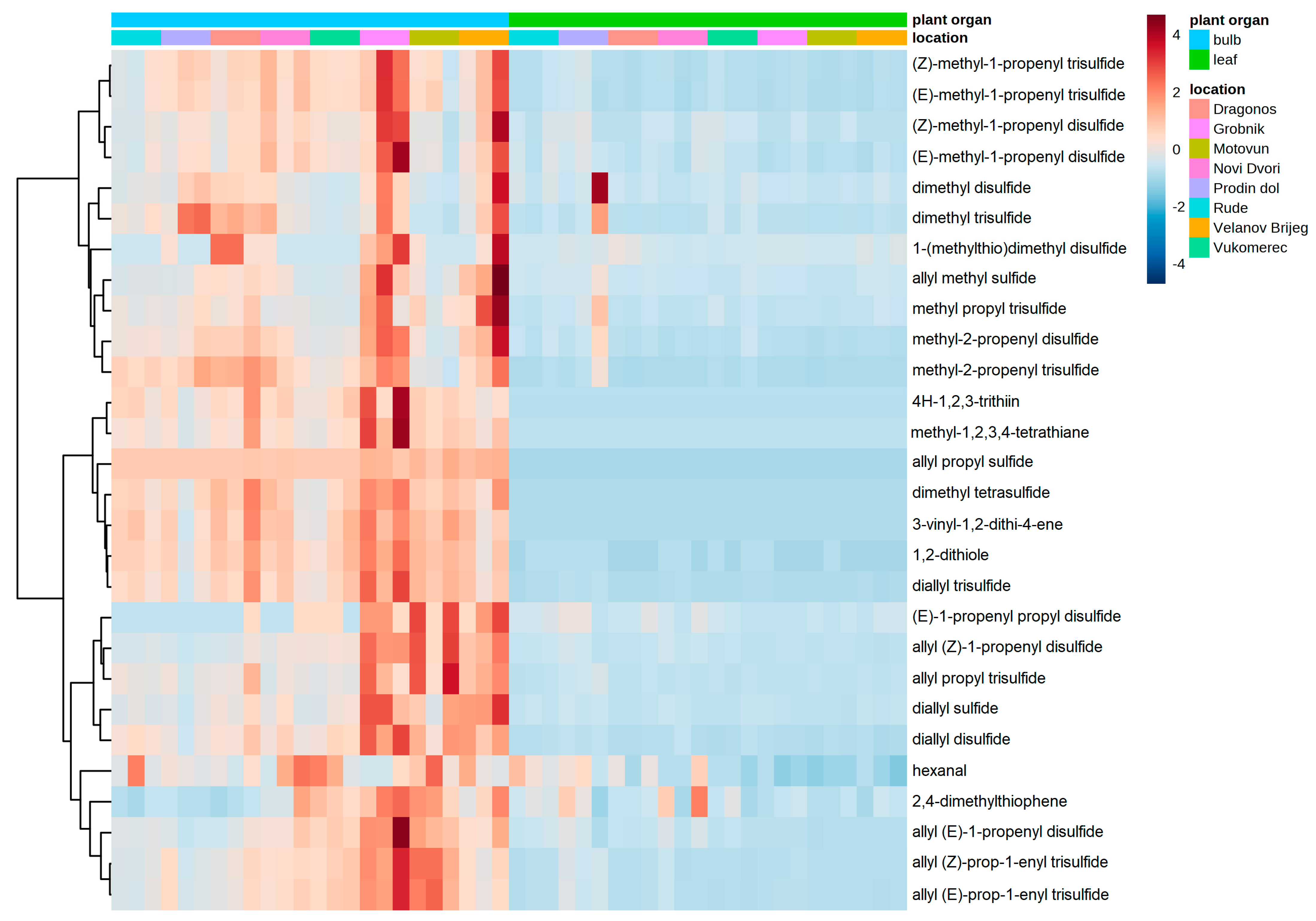{kind=link}
{kind=link}
| Compound | Aspartic Acid | Glutamic Acid | Histidine | Glycine | Threonine | Arginine | Tyrosine | Valine | Methionine | Phenyl-Alanine | Isoleucine | Leucine | Lysine | Cystine |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total phenolic content | −0.27 | −0.35 | −0.25 | −0.18 | 0.58 | 0.23 | 0.15 | 0.01 | 0.44 | 0.29 | 0.36 | 0.30 | 0.41 | 0.02 |
| Allyl methyl sulfide | −0.41 | −0.42 | −0.09 | −0.17 | 0.30 | −0.06 | −0.23 | −0.11 | 0.15 | −0.07 | −0.07 | −0.08 | −0.01 | −0.37 |
| Hexanal | −0.36 | −0.28 | −0.02 | 0.05 | 0.42 | 0.09 | −0.04 | 0.19 | 0.40 | 0.28 | 0.26 | 0.19 | 0.14 | −0.01 |
| Diallyl sulfide | −0.80 | −0.77 | −0.36 | −0.40 | 0.74 | 0.29 | 0.06 | 0.08 | 0.73 | 0.42 | 0.47 | 0.41 | 0.46 | −0.21 |
| Allyl propyl sulfide | −0.54 | −0.55 | −0.20 | −0.30 | 0.32 | −0.11 | −0.38 | −0.23 | 0.17 | −0.13 | −0.13 | −0.15 | −0.08 | −0.51 |
| 2,4-dimethylthiophene | −0.31 | −0.28 | −0.18 | −0.25 | 0.26 | −0.17 | −0.45 | −0.34 | −0.07 | −0.30 | −0.27 | −0.31 | −0.29 | −0.58 |
| Dimethyl disulfide | −0.11 | −0.19 | 0.26 | 0.01 | 0.22 | 0.08 | 0.14 | 0.22 | 0.16 | 0.12 | 0.12 | 0.11 | 0.13 | −0.03 |
| Methyl-2-propenyl disulfide | −0.52 | −0.52 | −0.13 | −0.24 | 0.44 | 0.05 | −0.09 | 0.02 | 0.36 | 0.12 | 0.13 | 0.11 | 0.17 | −0.29 |
| (Z)-methyl-1-propenyl disulfide | −0.47 | −0.45 | −0.21 | −0.23 | 0.44 | 0.00 | −0.20 | −0.10 | 0.24 | −0.01 | 0.02 | −0.01 | 0.06 | −0.39 |
| (E)-methyl-1-propenyl disulfide | −0.49 | −0.46 | −0.21 | −0.25 | 0.45 | 0.03 | −0.16 | −0.07 | 0.28 | 0.04 | 0.08 | 0.04 | 0.09 | −0.37 |
| 1,2-dithiole | −0.72 | −0.70 | −0.33 | −0.43 | 0.57 | 0.14 | −0.09 | 0.03 | 0.60 | 0.30 | 0.32 | 0.27 | 0.30 | −0.26 |
| Diallyl disulfide | −0.66 | −0.67 | −0.28 | −0.39 | 0.44 | −0.04 | −0.30 | −0.15 | 0.37 | 0.05 | 0.06 | 0.02 | 0.05 | −0.44 |
| Allyl (Z)-1-propenyl disulfide | −0.54 | −0.55 | −0.24 | −0.35 | 0.28 | −0.13 | −0.46 | −0.34 | 0.09 | −0.21 | −0.19 | −0.23 | −0.19 | −0.60 |
| Allyl (E)-1-propenyl disulfide | −0.54 | −0.52 | −0.27 | −0.35 | 0.37 | −0.11 | −0.38 | −0.25 | 0.18 | −0.11 | −0.09 | −0.13 | −0.12 | −0.52 |
| (E)-1-propenyl propyl disulfide | −0.45 | −0.43 | −0.15 | −0.25 | 0.24 | −0.15 | −0.45 | −0.32 | −0.01 | −0.26 | −0.25 | −0.28 | −0.24 | −0.58 |
| 1-(methylthio)dimethyl disulfide | −0.39 | −0.38 | −0.16 | −0.22 | 0.24 | 0.00 | −0.17 | −0.11 | 0.15 | −0.05 | −0.04 | −0.03 | 0.00 | −0.27 |
| Dimethyl trisulfide | −0.36 | −0.34 | −0.01 | −0.16 | 0.42 | 0.40 | 0.33 | 0.26 | 0.47 | 0.38 | 0.42 | 0.41 | 0.47 | 0.02 |
| Methyl-2-propenyl trisulfide | −0.62 | −0.59 | −0.20 | −0.31 | 0.59 | 0.27 | 0.15 | 0.20 | 0.64 | 0.42 | 0.44 | 0.41 | 0.46 | −0.10 |
| Methyl propyl trisulfide | −0.40 | −0.43 | −0.04 | −0.17 | 0.34 | −0.02 | −0.13 | −0.03 | 0.20 | 0.00 | 0.01 | 0.00 | 0.06 | −0.32 |
| (Z)-methyl-1-propenyl trisulfide | −0.61 | −0.56 | −0.26 | −0.33 | 0.57 | 0.15 | −0.08 | −0.05 | 0.42 | 0.14 | 0.20 | 0.15 | 0.22 | −0.36 |
| (E)-methyl-1-propenyl trisulfide | −0.62 | −0.58 | −0.27 | −0.34 | 0.57 | 0.13 | −0.11 | −0.08 | 0.40 | 0.11 | 0.17 | 0.12 | 0.19 | −0.39 |
| 3-vinyl-1,2-dithi-4-ene | −0.74 | −0.72 | −0.34 | −0.42 | 0.55 | 0.14 | −0.11 | 0.01 | 0.60 | 0.30 | 0.31 | 0.27 | 0.29 | −0.25 |
| 4H-1,2,3-trithiin | −0.64 | −0.63 | −0.34 | −0.40 | 0.45 | 0.07 | −0.14 | −0.03 | 0.49 | 0.22 | 0.23 | 0.20 | 0.21 | −0.26 |
| Dimethyl tetrasulfide | −0.62 | −0.61 | −0.32 | −0.41 | 0.37 | −0.01 | −0.25 | −0.13 | 0.36 | 0.08 | 0.09 | 0.06 | 0.08 | −0.38 |
| Diallyl trisulfide | 0.47 | 0.49 | 0.74 | 0.21 | −0.24 | −0.21 | 0.12 | 0.38 | −0.22 | 0.01 | −0.08 | −0.10 | −0.22 | 0.16 |
| Allyl propyl trisulfide | −0.54 | −0.56 | −0.21 | −0.34 | 0.26 | −0.03 | −0.33 | −0.22 | 0.19 | −0.07 | −0.06 | −0.09 | −0.06 | −0.48 |
| Allyl (Z)-prop-1-enyl trisulfide | −0.60 | −0.59 | −0.25 | −0.41 | 0.40 | −0.03 | −0.31 | −0.23 | 0.26 | −0.03 | 0.01 | −0.06 | −0.03 | −0.49 |
| Allyl (E)-prop-1-enyl trisulfide | −0.60 | −0.59 | −0.25 | −0.40 | 0.41 | −0.02 | −0.29 | −0.22 | 0.26 | −0.02 | 0.02 | −0.05 | −0.02 | −0.48 |
| Methyl-1,2,3,4-tetrathiane | −0.74 | −0.71 | −0.34 | −0.41 | 0.56 | 0.16 | −0.10 | 0.00 | 0.58 | 0.28 | 0.30 | 0.26 | 0.30 | −0.27 |
| Compounds | DPPH | FRAP | ORAC |
|---|---|---|---|
| Total phenolic content | 0.37 | 0.47 | −0.30 |
| Amino acids | |||
| Aspartic acid | −0.37 | −0.23 | 0.68 |
| Glutamic acid | −0.28 | −0.15 | 0.67 |
| Histidine | −0.20 | −0.13 | 0.26 |
| Glycine | −0.11 | −0.05 | 0.34 |
| Threonine | 0.46 | 0.47 | −0.66 |
| Arginine | 0.55 | 0.69 | −0.12 |
| Tyrosine | 0.66 | 0.72 | 0.05 |
| Valine | 0.59 | 0.56 | −0.01 |
| Methionine | 0.88 | 0.75 | −0.59 |
| Phenylalanine | 0.86 | 0.78 | −0.30 |
| Isoleucine | 0.87 | 0.84 | −0.34 |
| Leucine | 0.89 | 0.82 | −0.28 |
| Lysine | 0.84 | 0.84 | −0.32 |
| Cystine | 0.57 | 0.56 | 0.27 |
| Volatile organic compounds | |||
| Allyl methyl sulfide | −0.01 | −0.15 | −0.55 |
| Hexanal | 0.24 | 0.16 | −0.58 |
| Diallyl sulfide | 0.48 | 0.32 | −0.93 |
| Allyl propyl sulfide | −0.09 | −0.25 | −0.69 |
| 2,4-dimethylthiophene | −0.31 | −0.40 | −0.52 |
| Dimethyl disulfide | 0.13 | 0.04 | −0.32 |
| Methyl-2-propenyl disulfide | 0.20 | 0.02 | −0.68 |
| (Z)-methyl-1-propenyl disulfide | 0.07 | −0.08 | −0.60 |
| (E)-methyl-1-propenyl disulfide | 0.12 | −0.04 | −0.62 |
| 1,2-dithiole | 0.38 | 0.17 | −0.88 |
| Diallyl disulfide | 0.10 | −0.12 | −0.83 |
| Allyl (Z)-1-propenyl disulfide | −0.19 | −0.38 | −0.73 |
| Allyl (E)-1-propenyl disulfide | −0.08 | −0.27 | −0.72 |
| (E)-1-propenyl propyl disulfide | −0.28 | −0.41 | −0.63 |
| 1-(methylthio)dimethyl disulfide | 0.10 | −0.07 | −0.45 |
| Dimethyl trisulfide | 0.47 | 0.37 | −0.48 |
| Methyl-2-propenyl trisulfide | 0.53 | 0.35 | −0.74 |
| Methyl propyl trisulfide | 0.01 | −0.09 | −0.56 |
| (Z)-methyl-1-propenyl trisulfide | 0.24 | 0.09 | −0.75 |
| (E)-methyl-1-propenyl trisulfide | 0.20 | 0.06 | −0.77 |
| 3-vinyl-1,2-dithi-4-ene | 0.39 | 0.18 | −0.87 |
| 4H-1,2,3-trithiin | 0.29 | 0.08 | −0.76 |
| Dimethyl tetrasulfide | 0.15 | −0.06 | −0.74 |
| Diallyl trisulfide | −0.19 | −0.21 | 0.05 |
| Allyl propyl trisulfide | −0.06 | −0.22 | −0.71 |
| Allyl (Z)-prop-1-enyl trisulfide | 0.00 | −0.18 | −0.80 |
| Allyl (E)-prop-1-enyl trisulfide | 0.01 | −0.17 | −0.80 |
| Methyl-1,2,3,4-tetrathiane | 0.39 | 0.17 | −0.86 |
| Plant Organ | TPC mg GAE/g DW | DPPH | FRAP | ORAC nmol TE/g DW |
|---|---|---|---|---|
| µmol TE/g DW | ||||
| Bulb | 11.2 ± 0.3 | 9.80 ± 1.10 | 11.8 ± 1.0 | 87.2 ± 8.5 |
| Leaf | 10.4 ± 0.2 | 5.00 ± 0.20 | 8.80 ± 0.10 | 293 ± 10 |
| p-value | *** | *** | *** | *** |
| Location | ||||
| Rude | 10.1 ± 0.4 c 1 | 10.8 ± 2.4 a | 11.3 ± 1.3 c | 212 ± 43 ab |
| Prodin dol | 10.7 ± 0.3 bc | 10.0 ± 2.3 a | 13.9 ± 2.2 a | 195 ± 25 a–c |
| Vukomerec | 12.0 ± 0.7 a | 6.80 ± 1.10 b | 11.5 ± 1.3 bc | 229 ± 58 a |
| Motovun | 11.0 ± 0.3 bc | 4.00 ± 0.70 c | 6.90 ± 0.90 e | 165 ± 62 cd |
| Velanov Brijeg | 10.7 ± 0.3 bc | 4.30 ± 0.40 c | 8.80 ± 0.60 d | 165 ± 46 cd |
| Dragonoš | 10.2 ± 0.5 c | 10.0 ± 2.0 a | 12.5 ± 1.7 b | 222 ± 45 a |
| Novi Dvori | 11.5 ± 0.4 ab | 7.80 ± 1.70 b | 10.6 ± 1.2 c | 178 ± 47 b–d |
| Grobnik | 10.3 ± 0.5 c | 5.20 ± 0.30 c | 6.80 ± 0.80 e | 152 ± 51 d |
| p-value | ** | *** | *** | *** |
| Location | Bulb | |||
| Rude | 10.7 ± 0.2 c–f | 15.9 ± 1.1 a | 14.0 ± 0.8 c | 117 ± 4 e–g |
| Prodin dol | 11.4 ± 0.3 bc | 15.0 ± 1.1 a | 18.8 ± 0.2 a | 146 ± 13 e |
| Vukomerec | 13.2 ± 0.6 a | 9.28 ± 0.63 c | 14.2 ± 0.8 c | 100 ± 8 e–g |
| Motovun | 10.7 ± 0.3 c–f | 3.15 ± 1.19 e | 4.91 ± 0.35 f | 26.1 ± 2.7 h |
| Velanov Brijeg | 10.4 ± 0.5 c–g | 3.90 ± 0.74 de | 7.78 ± 0.87 e | 66.2 ± 20.8 gh |
| Dragonoš | 10.9 ± 0.2 c–e | 14.4 ± 0.5 a | 16.2 ± 0.8 b | 120 ± 5 ef |
| Novi Dvori | 12.4 ± 0.0 ab | 11.4 ± 0.4 b | 13.0 ± 1.0 c | 72.9 ± 3.9 f–h |
| Grobnik | 9.85 ± 0.80 e–g | 4.85 ± 0.62 de | 5.17 ± 0.36 f | 46.3 ± 3.5 h |
| Leaf | ||||
| Rude | 9.48 ± 0.68 fg | 5.61 ± 0.38 d | 8.61 ± 0.32 de | 306 ± 20 bc |
| Prodin dol | 10.0 ± 0.0 d–g | 4.97 ± 0.15 de | 8.92 ± 0.13 de | 245 ± 24 d |
| Vukomerec | 10.6 ± 0.2 c–f | 4.35 ± 0.19 de | 8.67 ± 0.02 de | 358 ± 17 a |
| Motovun | 11.2 ± 0.6 b–d | 4.92 ± 0.36 de | 8.83 ± 0.30 de | 304 ± 5 bc |
| Velanov Brijeg | 10.9 ± 0.1 c–e | 4.76 ± 0.41 de | 9.76 ± 0.32 d | 264 ± 25 cd |
| Dragonoš | 9.36 ± 0.60 g | 5.49 ± 0.22 d | 8.74 ± 0.12 de | 323 ± 5 ab |
| Novi Dvori | 10.5 ± 0.1 c–g | 4.18 ± 0.97 de | 8.21 ± 0.24 de | 283 ± 9 b–d |
| Grobnik | 10.7 ± 0.5 c–f | 5.61 ± 0.16 d | 8.45 ± 0.18 de | 257 ± 47 cd |
| p-value | ** | *** | *** | ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovačević, T.K.; Major, N.; Sivec, M.; Horvat, D.; Krpan, M.; Hruškar, M.; Ban, D.; Išić, N.; Goreta Ban, S. Phenolic Content, Amino Acids, Volatile Compounds, Antioxidant Capacity, and Their Relationship in Wild Garlic (A. ursinum L.). Foods 2023, 12, 2110. https://doi.org/10.3390/foods12112110
Kovačević TK, Major N, Sivec M, Horvat D, Krpan M, Hruškar M, Ban D, Išić N, Goreta Ban S. Phenolic Content, Amino Acids, Volatile Compounds, Antioxidant Capacity, and Their Relationship in Wild Garlic (A. ursinum L.). Foods. 2023; 12(11):2110. https://doi.org/10.3390/foods12112110
Chicago/Turabian StyleKovačević, Tvrtko Karlo, Nikola Major, Marta Sivec, Dijana Horvat, Marina Krpan, Mirjana Hruškar, Dean Ban, Nina Išić, and Smiljana Goreta Ban. 2023. "Phenolic Content, Amino Acids, Volatile Compounds, Antioxidant Capacity, and Their Relationship in Wild Garlic (A. ursinum L.)" Foods 12, no. 11: 2110. https://doi.org/10.3390/foods12112110
APA StyleKovačević, T. K., Major, N., Sivec, M., Horvat, D., Krpan, M., Hruškar, M., Ban, D., Išić, N., & Goreta Ban, S. (2023). Phenolic Content, Amino Acids, Volatile Compounds, Antioxidant Capacity, and Their Relationship in Wild Garlic (A. ursinum L.). Foods, 12(11), 2110. https://doi.org/10.3390/foods12112110