
Lactiplantibacillus paraplantarum BPF2 and Pediococcus acidilactici ST6, Two Bacteriocinogenic Isolated Strains from Andalusian Spontaneous Fermented Sausages
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Indicator Bacterial Strains and Culture Conditions
2.2. Isolation of LAB Species from Dry-Fermented Sausages
2.3. Screening and Selection of Antagonist LAB Strains
2.4. Strain Identification and Characterization
2.5. Bacteriocin Production and Purification Assay
2.6. Bacteriocin Stability against Heat, pH, and Enzymes
2.7. Antagonism Assays in Cocultures
2.8. Statistical Analysis
3. Results and Discussion
3.1. Isolation of LAB Species from Dry-Fermented Sausages
3.2. Screening and Selection of Antagonist LAB Strains
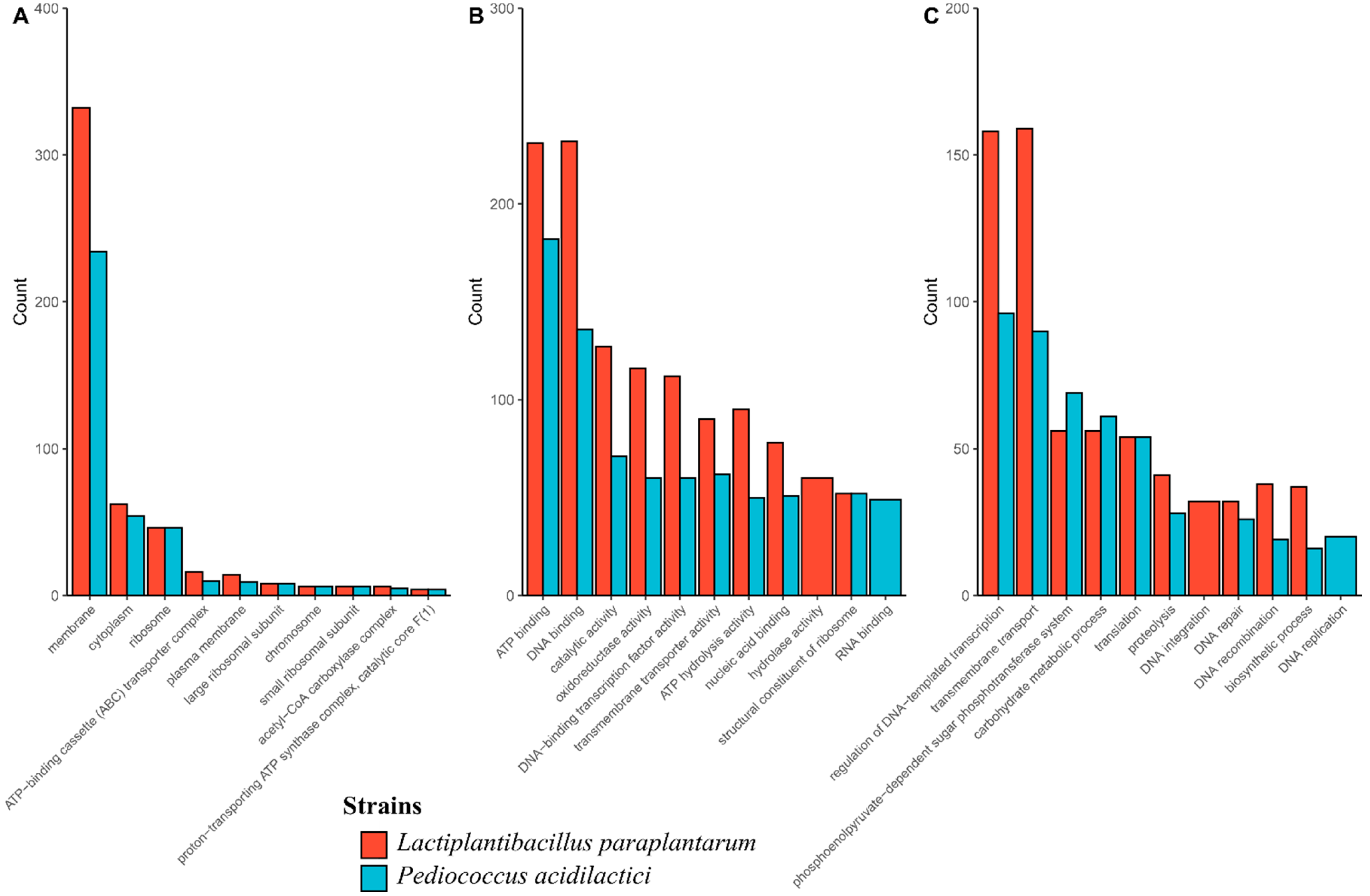
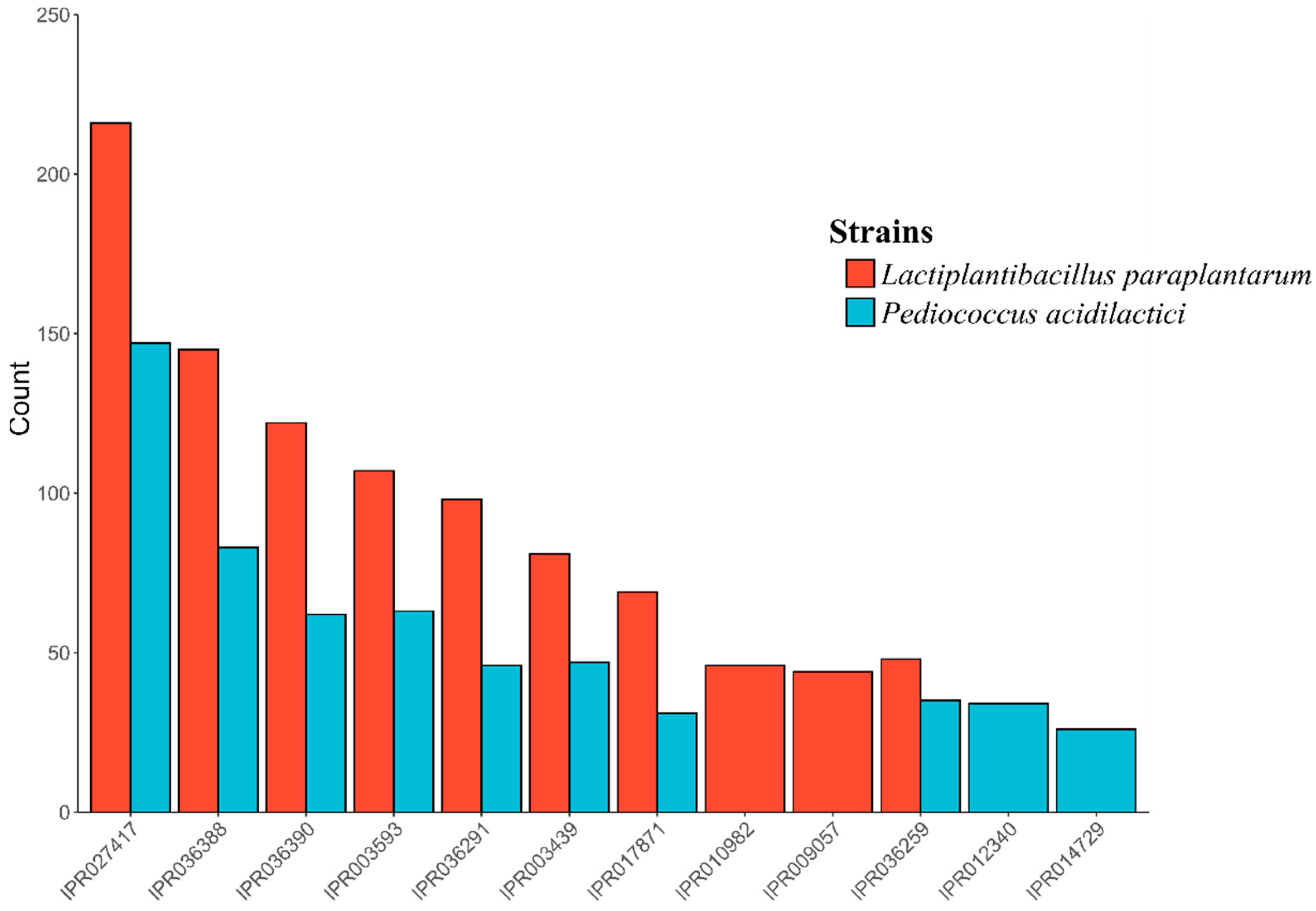
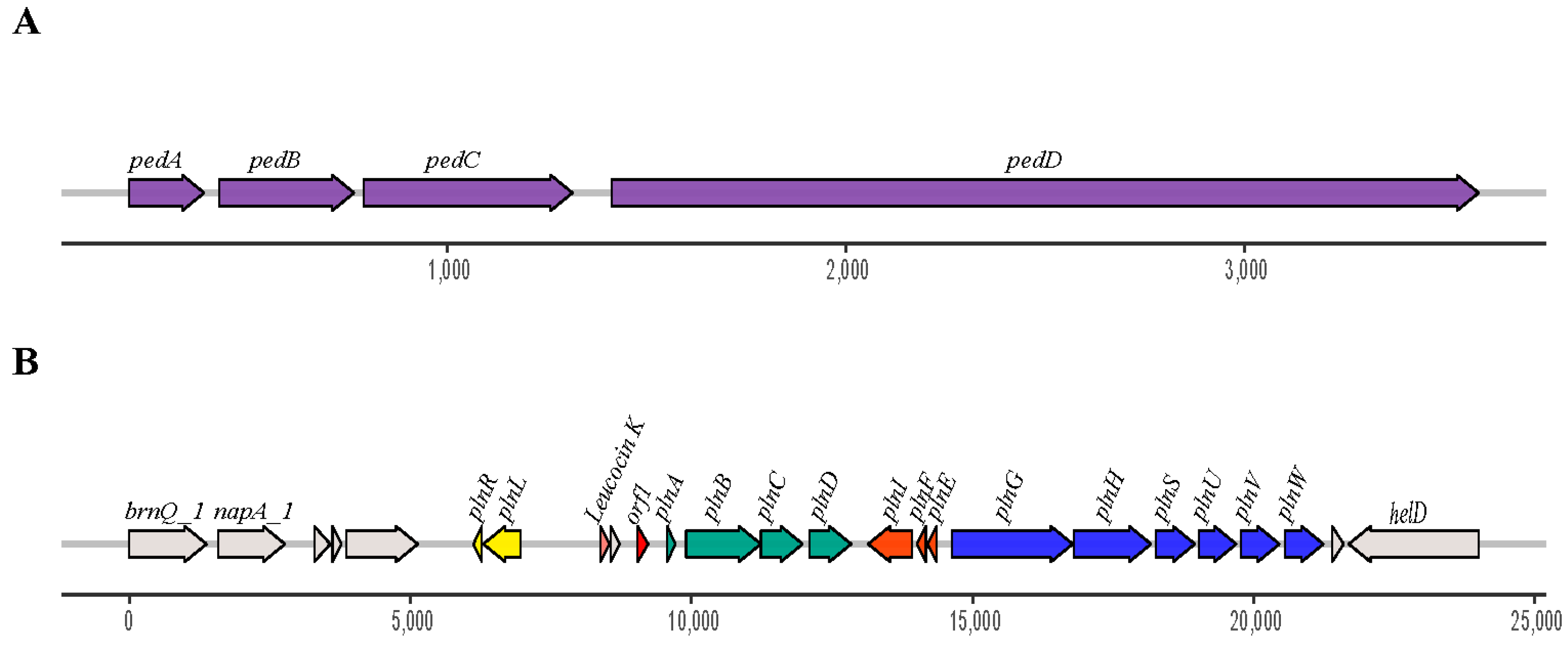
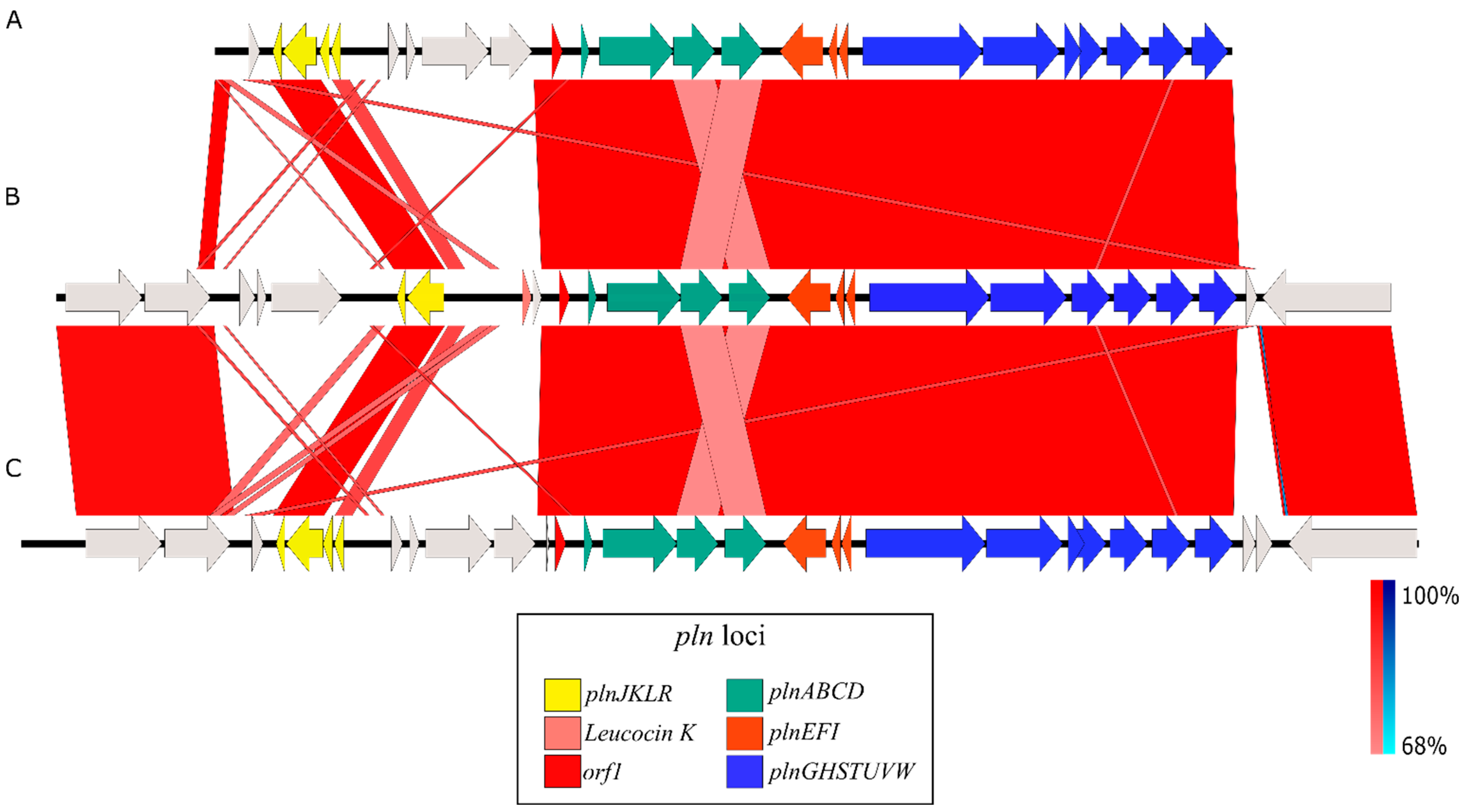
3.3. Identification and Genomic Studies
3.4. Identification of the Bacteriocins by LC-MS/MS
3.5. Effect of Temperature, pH, and Different Enzymes on Bacteriocins
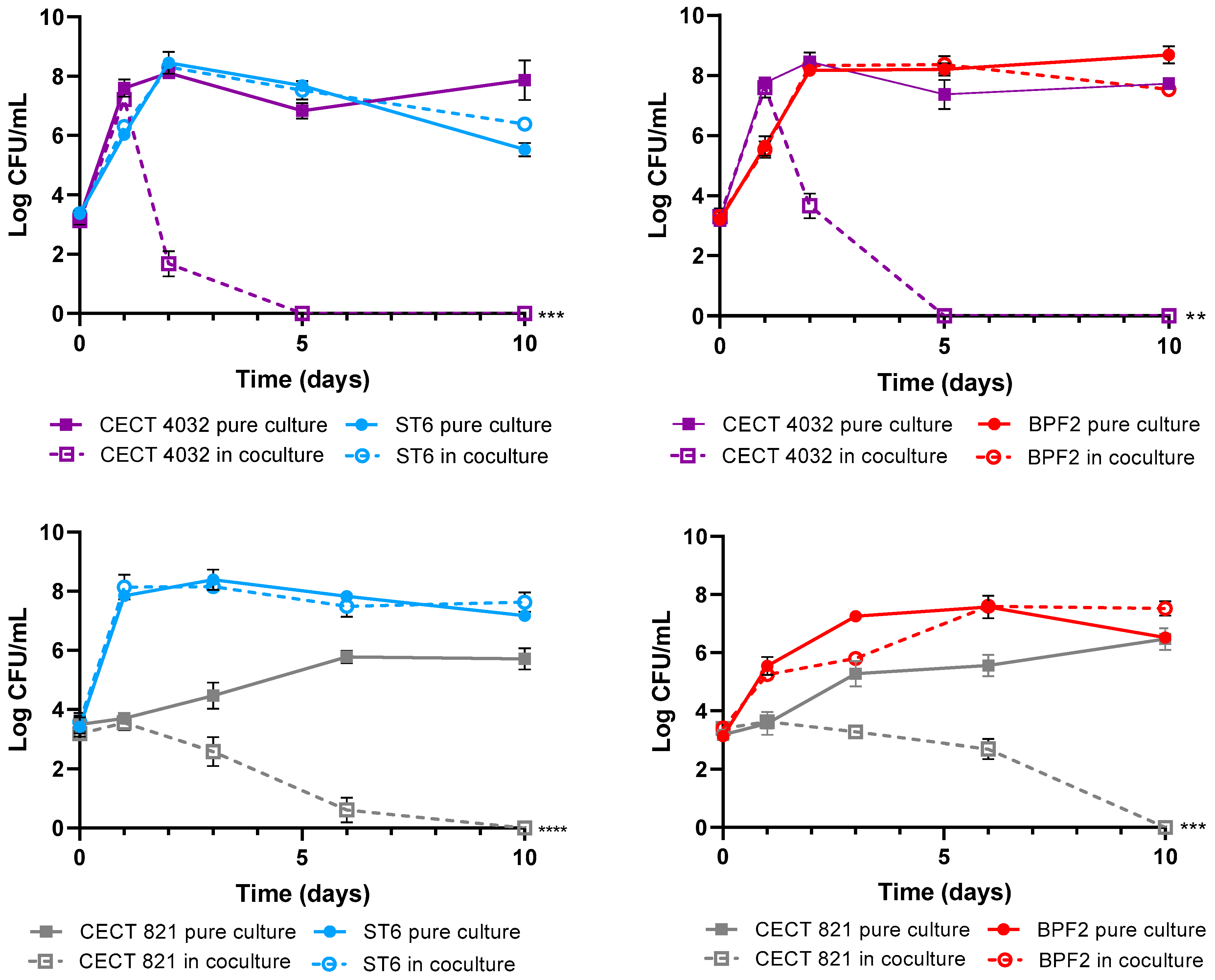
3.6. Antagonism Assays in Cocultures
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lücke, F.-K. Fermented Sausages. In Microbiology of Fermented Foods; Wood, B.J.B., Ed.; Springer: Boston, MA, USA, 1998; pp. 441–483. ISBN 978-1-4613-0309-1. [Google Scholar]
- Zagorec, M.; Champomier-Vergès, M.-C. Lactobacillus Sakei: A Starter for Sausage Fermentation, a Protective Culture for Meat Products. Microorganisms 2017, 5, 56. [Google Scholar] [CrossRef] [PubMed]
- Zeuthen, P. Historical Aspects of Meat Fermentations. In Fermented Meats; Campbell-Platt, G., Cook, P.E., Eds.; Springer: Boston, MA, USA, 1995; pp. 53–68. ISBN 978-1-4615-2163-1. [Google Scholar]
- Oliveira, M.; Ferreira, V.; Magalhães, R.; Teixeira, P. Biocontrol Strategies for Mediterranean-Style Fermented Sausages. Food Res. Int. 2018, 103, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Bungenstock, L.; Abdulmawjood, A.; Reich, F. Evaluation of antibacterial properties of lactic acid bacteria from traditionally and industrially produced fermented sausages from Germany. PLoS ONE 2020, 15, e0230345. [Google Scholar] [CrossRef]
- Leroy, F.; De Vuyst, L. Lactic Acid Bacteria as Functional Starter Cultures for the Food Fermentation Industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Drosinos, E.H.; Mataragas, M.; Xiraphi, N.; Moschonas, G.; Gaitis, F.; Metaxopoulos, J. Characterization of the Microbial Flora from a Traditional Greek Fermented Sausage. Meat Sci. 2005, 69, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Landeta, G.; Curiel, J.A.; Carrascosa, A.V.; Muñoz, R.; de las Rivas, B. Technological and Safety Properties of Lactic Acid Bacteria Isolated from Spanish Dry-Cured Sausages. Meat Sci. 2013, 95, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, S.; Cachaldora, A.; Gómez, M.; Franco, I.; Carballo, J. Monitoring the Bacterial Population Dynamics during the Ripening of Galician Chorizo, a Traditional Dry Fermented Spanish Sausage. Food Microbiol. 2013, 33, 77–84. [Google Scholar] [CrossRef]
- Talon, R.; Leroy, S.; Lebert, I. Microbial Ecosystems of Traditional Fermented Meat Products: The Importance of Indigenous Starters. Meat Sci. 2007, 77, 55–62. [Google Scholar] [CrossRef]
- Essid, I.; Ben Ismail, H.; Bel Hadj Ahmed, S.; Ghedamsi, R.; Hassouna, M. Characterization and Technological Properties of Staphylococcus Xylosus Strains Isolated from a Tunisian Traditional Salted Meat. Meat Sci. 2007, 77, 204–212. [Google Scholar] [CrossRef]
- Noonpakdee, W.; Santivarangkna, C.; Jumriangrit, P.; Sonomoto, K.; Panyim, S. Isolation of Nisin-Producing Lactococcus Lactis WNC 20 Strain from Nham, a Traditional Thai Fermented Sausage. Int. J. Food Microbiol. 2003, 81, 137–145. [Google Scholar] [CrossRef]
- Zhu, Y.; Yang, Q. Isolation of Antibacterial, Nitrosylmyoglobin Forming Lactic Acid Bacteria and Their Potential Use in Meat Processing. Front. Microbiol. 2020, 11, 1315. [Google Scholar] [CrossRef] [PubMed]
- Jack, R.W.; Tagg, J.R.; Ray, B. Bacteriocins of Gram-Positive Bacteria. Microbiol. Rev. 1995, 59, 171–200. [Google Scholar] [CrossRef] [PubMed]
- Gálvez, A.; López, R.L.; Abriouel, H.; Valdivia, E.; Omar, N.B. Application of Bacteriocins in the Control of Foodborne Pathogenic and Spoilage Bacteria. Crit. Rev. Biotechnol. 2008, 28, 125–152. [Google Scholar] [CrossRef]
- Heo, S.; Kim, M.G.; Kwon, M.; Lee, H.S.; Kim, G.-B. Inhibition of Clostridium Perfringens Using Bacteriophages and Bacteriocin Producing Strains. Korean J. Food Sci. Anim. Resour. 2018, 38, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Todorov, S.D.; Popov, I.; Weeks, R.; Chikindas, M.L. Use of Bacteriocins and Bacteriocinogenic Beneficial Organisms in Food Products: Benefits, Challenges, Concerns. Foods 2022, 11, 3145. [Google Scholar] [CrossRef] [PubMed]
- Nishie, M.; Nagao, J.-I.; Sonomoto, K. Antibacterial Peptides “Bacteriocins”: An Overview of Their Diverse Characteristics and Applications. Biocontrol Sci. 2012, 17, 1–16. [Google Scholar] [CrossRef]
- Juneja, V.K.; Dwivedi, H.P.; Yan, X. Novel Natural Food Antimicrobials. Annu. Rev. Food Sci. Technol. 2012, 3, 381–403. [Google Scholar] [CrossRef]
- Ribeiro, S.C.; Coelho, M.C.; Todorov, S.D.; Franco, B.D.G.M.; Dapkevicius, M.L.E.; Silva, C.C.G. Technological Properties of Bacteriocin-producing Lactic Acid Bacteria Isolated from Pico Cheese an Artisanal Cow’s Milk Cheese. J. Appl. Microbiol. 2014, 116, 573–585. [Google Scholar] [CrossRef]
- Kamiloğlu, A. Functional and Technological Characterization of Lactic Acid Bacteria Isolated from Turkish Dry-Fermented Sausage (Sucuk). Braz. J. Microbiol. 2022, 53, 959–968. [Google Scholar] [CrossRef]
- Barbieri, F.; Tabanelli, G.; Montanari, C.; Dall’Osso, N.; Šimat, V.; Smole Možina, S.; Baños, A.; Özogul, F.; Bassi, D.; Fontana, C.; et al. Mediterranean Spontaneously Fermented Sausages: Spotlight on Microbiological and Quality Features to Exploit Their Bacterial Biodiversity. Foods 2021, 10, 2691. [Google Scholar] [CrossRef]
- Alonso, S.; Carmen Castro, M.; Berdasco, M.; de la Banda, I.G.; Moreno-Ventas, X.; de Rojas, A.H. Isolation and Partial Characterization of Lactic Acid Bacteria from the Gut Microbiota of Marine Fishes for Potential Application as Probiotics in Aquaculture. Probiotics Antimicrob. Proteins 2019, 11, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Tagg, J.R.; McGiven, A.R. Assay System for Bacteriocins. Appl. Microbiol. 1971, 21, 943. [Google Scholar] [CrossRef] [PubMed]
- Gálvez, A.; Maqueda, M.; Valdivia, E.; Quesada, A.; Montoya, E. Characterization and Partial Purification of a Broad Spectrum Antibiotic AS-48 Produced by Streptococcus faecalis. Can. J. Microbiol. 1986, 32, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Martín-Platero, A.M.; Valdivia, E.; Maqueda, M.; Martínez-Bueno, M. Fast, Convenient, and Economical Method for Isolating Genomic DNA from Lactic Acid Bacteria Using a Modification of the Protein “Salting-out” Procedure. Anal. Biochem. 2007, 366, 102–104. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A New Generation of Protein Database Search Programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing|Journal of Computational Biology. Available online: https://www.liebertpub.com/doi/abs/10.1089/cmb.2012.0021 (accessed on 6 March 2023).
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A Greedy Algorithm for Aligning DNA Sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- InterProScan 5: Genome-Scale Protein Function Classification|Bioinformatics|Oxford Academic. Available online: https://academic.oup.com/bioinformatics/article/30/9/1236/237988?login=false (accessed on 6 March 2023).
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Res. 2023, 51, D418–D427. [Google Scholar] [CrossRef]
- Gene Ontology: Tool for the Unification of Biology|Nature Genetics. Available online: https://www.nature.com/articles/ng0500_25 (accessed on 6 March 2023).
- Binns, D.; Dimmer, E.; Huntley, R.; Barrell, D.; O’Donovan, C.; Apweiler, R. QuickGO: A Web-Based Tool for Gene Ontology Searching. Bioinformatics 2009, 25, 3045–3046. [Google Scholar] [CrossRef]
- The Gene Ontology Consortium the Gene Ontology Resource: Enriching a GOld Mine. Nucleic Acids Res. 2021, 49, D325–D334. [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Hammami, R.; Zouhir, A.; Ben Hamida, J.; Fliss, I. BACTIBASE: A New Web-Accessible Database for Bacteriocin Characterization. BMC Microbiol. 2007, 7, 89. [Google Scholar] [CrossRef]
- Abriouel, H.; Valdivia, E.; Martínez-Bueno, M.; Maqueda, M.; Gálvez, A. A Simple Method for Semi-Preparative-Scale Production and Recovery of Enterocin AS-48 Derived from Enterococcus faecalis Subsp. Liquefaciens A-48-32. J. Microbiol. Methods 2003, 55, 599–605. [Google Scholar] [CrossRef]
- Schägger, H.; von Jagow, G. Tricine-Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis for the Separation of Proteins in the Range from 1 to 100 KDa. Anal. Biochem. 1987, 166, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.; Kim, J.; Kim, W.J. Isolation and Characterization of Bacteriocin-Producing Pediococcus acidilactici HW01 from Malt and Its Potential to Control Beer Spoilage Lactic Acid Bacteria. Food Control 2017, 80, 59–66. [Google Scholar] [CrossRef]
- Zhao, R.; Lu, Y.; Ran, J.; Li, G.; Lei, S.; Zhu, Y.; Xu, B. Purification and Characterization of Bacteriocin Produced by Lactobacillus Rhamnosus Zrx01. Food Biosci. 2020, 38, 100754. [Google Scholar] [CrossRef]
- Bizani, D.; Brandelli, A. Characterization of a Bacteriocin Produced by a Newly Isolated Bacillus Sp. Strain 8 A. J. Appl. Microbiol. 2002, 93, 512–519. [Google Scholar] [CrossRef]
- Zamfir, M.; Callewaert, R.; Cornea, P.C.; Savu, L.; Vatafu, I.; De Vuyst, L. Purification and Characterization of a Bacteriocin Produced by Lactobacillus Acidophilus IBB 801. J. Appl. Microbiol. 1999, 87, 923–931. [Google Scholar] [CrossRef]
- Baños, A.; Ariza, J.J.; Nuñez, C.; Gil-Martínez, L.; García-López, J.D.; Martínez-Bueno, M.; Valdivia, E. Effects of Enterococcus Faecalis UGRA10 and the Enterocin AS-48 against the Fish Pathogen Lactococcus Garvieae. Studies In Vitro and In Vivo. Food Microbiol. 2019, 77, 69–77. [Google Scholar] [CrossRef]
- Ayala, D.I.; Cook, P.W.; Franco, J.G.; Bugarel, M.; Kottapalli, K.R.; Loneragan, G.H.; Brashears, M.M.; Nightingale, K.K. A Systematic Approach to Identify and Characterize the Effectiveness and Safety of Novel Probiotic Strains to Control Foodborne Pathogens. Front. Microbiol. 2019, 10, 1108. [Google Scholar] [CrossRef] [PubMed]
- Erdoğmuş, S.F.; Erişmiş, U.C.; Uğuz, C. Isolation and Identification of Lactic Acid Bacteria from Fermented Meat Products and Evaluation of Their Antimicrobial Effect. Czech J. Food Sci. 2021, 39, 289–296. [Google Scholar] [CrossRef]
- Comi, G.; Urso, R.; Iacumin, L.; Rantsiou, K.; Cattaneo, P.; Cantoni, C.; Cocolin, L. Characterisation of Naturally Fermented Sausages Produced in the North East of Italy. Meat Sci. 2005, 69, 381–392. [Google Scholar] [CrossRef]
- Ren, D.; Zhu, J.; Gong, S.; Liu, H.; Yu, H. Antimicrobial Characteristics of Lactic Acid Bacteria Isolated from Homemade Fermented Foods. BioMed Res. Int. 2018, 2018, 5416725. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.-C.; Oh, J.K.; Kim, S.H.; Oh, S.; Kang, D.-K. Isolation and Characterization of an Anti-Listerial Bacteriocin from Leuconostoc Lactis SD501. Korean J. Food Sci. Anim. Resour. 2018, 38, 1008–1018. [Google Scholar] [CrossRef] [PubMed]
- Essid, I.; Hassouna, M. Effect of Inoculation of Selected Staphylococcus Xylosus and Lactobacillus Plantarum Strains on Biochemical, Microbiological and Textural Characteristics of a Tunisian Dry Fermented Sausage. Food Control 2013, 32, 707–714. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, P.; Xie, Y.; Wang, X. Co-Fermentation with Lactobacillus Curvatus LAB26 and Pediococcus Pentosaceus SWU73571 for Improving Quality and Safety of Sour Meat. Meat Sci. 2020, 170, 108240. [Google Scholar] [CrossRef] [PubMed]
- Mao, B.; Yin, R.; Li, X.; Cui, S.; Zhang, H.; Zhao, J.; Chen, W. Comparative Genomic Analysis of Lactiplantibacillus Plantarum Isolated from Different Niches. Genes 2021, 12, 241. [Google Scholar] [CrossRef]
- Ranjan, R.; Pandit, R.J.; Duggirala, S.M.; Joshi, C.G.; Sharma, S.; Patil, N.V. Genome Sequencing of Pediococcus acidilactici (NRCC1), a Novel Isolate from Dromedary Camel (Camelus Dromedarius) Rumen Fluid. Ann. Microbiol. 2018, 68, 103–110. [Google Scholar] [CrossRef]
- Li, Z.; Song, Q.; Wang, M.; Ren, J.; Liu, S.; Zhao, S. Comparative Genomics Analysis of Pediococcus acidilactici Species. J. Microbiol. 2021, 59, 573–583. [Google Scholar] [CrossRef]
- Surachat, K.; Deachamag, P.; Kantachote, D.; Wonglapsuwan, M.; Jeenkeawpiam, K.; Chukamnerd, A. In Silico Comparative Genomics Analysis of Lactiplantibacillus Plantarum DW12, a Potential Gamma-Aminobutyric Acid (GABA)-Producing Strain. Microbiol. Res. 2021, 251, 126833. [Google Scholar] [CrossRef]
- Yetiman, A.E.; Keskin, A.; Darendeli, B.N.; Kotil, S.E.; Ortakci, F.; Dogan, M. Characterization of Genomic, Physiological, and Probiotic Features Lactiplantibacillus Plantarum DY46 Strain Isolated from Traditional Lactic Acid Fermented Shalgam Beverage. Food Biosci. 2022, 46, 101499. [Google Scholar] [CrossRef]
- Diep, D.B.; Håvarstein, L.S.; Nes, I.F. Characterization of the Locus Responsible for the Bacteriocin Production in Lactobacillus Plantarum C11. J. Bacteriol. 1996, 178, 4472–4483. [Google Scholar] [CrossRef]
- Kleerebezem, M.; Boekhorst, J.; van Kranenburg, R.; Molenaar, D.; Kuipers, O.P.; Leer, R.; Tarchini, R.; Peters, S.A.; Sandbrink, H.M.; Fiers, M.W.E.J.; et al. Complete Genome Sequence of Lactobacillus Plantarum WCFS1. Proc. Natl. Acad. Sci. USA 2003, 100, 1990–1995. [Google Scholar] [CrossRef]
- Diep, D.B.; Straume, D.; Kjos, M.; Torres, C.; Nes, I.F. An Overview of the Mosaic Bacteriocin Pln Loci from Lactobacillus Plantarum. Peptides 2009, 30, 1562–1574. [Google Scholar] [CrossRef]
- Nie, R.; Zhu, Z.; Qi, Y.; Wang, Z.; Sun, H.; Liu, G. Bacteriocin Production Enhancing Mechanism of Lactiplantibacillus paraplantarum RX-8 Response to Wickerhamomyces Anomalus Y-5 by Transcriptomic and Proteomic Analyses. Front. Microbiol. 2023, 14, 1111516. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.J.; Hwang, J.; Kang, W.K.; Ahn, J.-P.; Kim, H.J. Administration of the Probiotic Lactiplantibacillus paraplantarum Is Effective in Controlling Hyperphosphatemia in 5/6 Nephrectomy Rat Model. Life Sci. 2022, 306, 120856. [Google Scholar] [CrossRef] [PubMed]
- Skrzypczak, K.; Gustaw, K.; Jabłońska-Ryś, E.; Sławińska, A.; Gustaw, W.; Winiarczyk, S. Spontaneously Fermented Fruiting Bodies of Agaricus Bisporus as a Valuable Source of New Isolates of Lactic Acid Bacteria with Functional Potential. Foods 2020, 9, 1631. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zeng, J.; Wang, C.; Wang, J. Structural Basis of Pore Formation in the Mannose Phosphotransferase System by Pediocin PA-1. Appl. Environ. Microbiol. 2022, 88, e01992-21. [Google Scholar] [CrossRef] [PubMed]
- Balandin, S.V.; Sheremeteva, E.V.; Ovchinnikova, T.V. Pediocin-like Antimicrobial Peptides of Bacteria. Biochem. Mosc. 2019, 84, 464–478. [Google Scholar] [CrossRef]
- Arike, L.; Peil, L. Spectral Counting Label-Free Proteomics. In Shotgun Proteomics; Martins-de-Souza, D., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2014; Volume 1156, pp. 213–222. ISBN 978-1-4939-0684-0. [Google Scholar]
- Stiles, M.E. Bacteriocins Produced by Leuconostoc Species. J. Dairy Sci. 1994, 77, 2718–2724. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.M.; Martínez, M.I.; Kok, J. Pediocin PA-1, a Wide-Spectrum Bacteriocin from Lactic Acid Bacteria. Crit. Rev. Food Sci. Nutr. 2002, 42, 91–121. [Google Scholar] [CrossRef] [PubMed]
- Schved, F.; Lalazar, A.; Henis, Y.; Juven, B.J. Purification, Partial Characterization and Plasmid-Linkage of Pediocin SJ-1, a Bacteriocin Produced by Pediococcus acidilactici. J. Appl. Bacteriol. 1993, 74, 67–77. [Google Scholar] [CrossRef]
- Niamah, A.K. Structure, Mode of Action and Application of Pediocin Natural Antimicrobial Food Preservative: A Review. Basrah J. Agric. Sci. 2018, 31, 59–69. [Google Scholar] [CrossRef]
- Shi, F.; Wang, Y.; Li, Y.; Wang, X. Mode of Action of Leucocin K7 Produced by Leuconostoc Mesenteroides K7 against Listeria Monocytogenes and Its Potential in Milk Preservation. Biotechnol. Lett. 2016, 38, 1551–1557. [Google Scholar] [CrossRef]
- Fugaban, J.I.I.; Vazquez Bucheli, J.E.; Park, Y.J.; Suh, D.H.; Jung, E.S.; Franco, B.D.G.D.M.; Ivanova, I.V.; Holzapfel, W.H.; Todorov, S.D. Antimicrobial Properties of Pediococcus acidilactici and Pediococcus Pentosaceus Isolated from Silage. J. Appl. Microbiol. 2022, 132, 311–330. [Google Scholar] [CrossRef]
- Li, Z.; Li, F.; Kharazian, Z.A.; Guo, X. Effect of Inoculating Two Bacteriocin-Producing Lactiplantibacillus plantarum Strains at Ensiling on In Vitro Rumen Fermentation and Methane Emissions of Alfalfa Silage with Two Dry Matter Contents. Animals 2023, 13, 384. [Google Scholar] [CrossRef]
- Knipe, H.; Temperton, B.; Lange, A.; Bass, D.; Tyler, C.R. Probiotics and Competitive Exclusion of Pathogens in Shrimp Aquaculture. Rev. Aquac. 2021, 13, 324–352. [Google Scholar] [CrossRef]
- Nieto-Lozano, J.C.; Reguera-Useros, J.I.; Peláez-Martínez, M.D.C.; Sacristán-Pérez-Minayo, G.; Gutiérrez-Fernández, Á.J.; de la Torre, A.H. The Effect of the Pediocin PA-1 Produced by Pediococcus acidilactici against Listeria Monocytogenes and Clostridium Perfringens in Spanish Dry-Fermented Sausages and Frankfurters. Food Control 2010, 21, 679–685. [Google Scholar] [CrossRef]
- Nieto-Lozano, J.C.; Reguera-Useros, J.I.; Peláez-Martínez, M.D.C.; Hardisson de la Torre, A. Effect of a Bacteriocin Produced by Pediococcus acidilactici against Listeria Monocytogenes and Clostridium Perfringens on Spanish Raw Meat. Meat Sci. 2006, 72, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Luchansky, J.B.; Glass, K.A.; Harsono, K.D.; Degnan, A.J.; Faith, N.G.; Cauvin, B.; Baccus-Taylor, G.; Arihara, K.; Bater, B.; Maurer, A.J. Genomic Analysis of Pediococcus Starter Cultures Used to Control Listeria Monocytogenes in Turkey Summer Sausage. Appl. Environ. Microbiol. 1992, 58, 3053–3059. [Google Scholar] [CrossRef] [PubMed]
- Komora, N.; Maciel, C.; Amaral, R.A.; Fernandes, R.; Castro, S.M.; Saraiva, J.A.; Teixeira, P. Innovative Hurdle System towards Listeria Monocytogenes Inactivation in a Fermented Meat Sausage Model—High Pressure Processing Assisted by Bacteriophage P100 and Bacteriocinogenic Pediococcus acidilactici. Food Res. Int. 2021, 148, 110628. [Google Scholar] [CrossRef] [PubMed]
- Bin Lee, H.; Kim, K.H.; Kang, G.A.; Lee, K.-G.; Kang, S.-S. Antibiofilm, AntiAdhesive and Anti-Invasive Activities of Bacterial Lysates Extracted from Pediococcus acidilactici against Listeria Monocytogenes. Food 2022, 11, 2948. [Google Scholar] [CrossRef]
- Talukdar, P.K.; Udompijitkul, P.; Hossain, A.; Sarker, M.R. Inactivation Strategies for Clostridium Perfringens Spores and Vegetative Cells. Appl. Environ. Microbiol. 2016, 83, e02731-16. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | Reference | Isolation |
|---|---|---|
| Listeria monocytogenes | DSM 112142 | Minced meat |
| Listeria monocytogenes | CECT 4032 | Associated with a case of meningitis after eating soft cheese |
| Staphylococcus aureus | CECT 239 | Human lesion |
| Escherichia coli | CECT 516 | Human feces |
| Clostridium perfringens | CECT 821 | Sheep |
| LAB Strain | L. monocytogenes DSM 112142 | S. aureus CECT 520 | LAB Strain | L. monocytogenes DSM 112142 | S. aureus CECT 520 |
|---|---|---|---|---|---|
| SE1 | + | - | CPN1.1 * | +++ | + |
| SE2 | ++ | - | CPN1.3 | + | + |
| SA01 | ++ | + | CPN2.1 | + | + |
| SE3.5 | + | + | CPN2.2 | ++ | - |
| SA4 | ++ | - | CPN2.5 | + | - |
| SA4.7 | ++ | - | BPF1 * | ++ | ++ |
| SA4.7 | ++ | + | BPF2 * | +++ | ++ |
| SA5.2 | + | - | SBE13.6 | + | + |
| SA5.5 | ++ | - | SBE 15.1 | ++ | - |
| SA6.1 | ++ | - | SBE 15.3 | + | - |
| SA6.3 | ++ | + | CT1.1 | ++ | - |
| SA6.4 | + | + | CT1.4 | ++ | + |
| SB6.4 | ++ | - | CT1.4 | + | - |
| SB6.4 | ++ | + | CT1.5 | ++ | + |
| SB6.5 | + | + | CCU1.1 | + | - |
| SB7.3 | + | + | CCU1.14 | - | + |
| SB8.1 | ++ | - | CCU2.2 | +++ | - |
| SB8.7 | + | + | CCH2 * | +++ | ++ |
| SB8.6 | - | + | CCH4 | + | + |
| SB9.7 | ++ | + | CCH5 | ++ | + |
| CHE1.2 | ++ | + | CCH7 | ++ | + |
| CHE 1.4 | + | + | CCH9 * | +++ | + |
| CHE 2.1 | + | - | CCH11 * | +++ | - |
| CHE 1.3 | ++ | + | CT4.2 | + | - |
| CCA1.2 | - | + | ST5 | ++ | + |
| CCA1.2 II | + | + | CD2 | ++ | - |
| CCA 2.2 | - | + | CD4 | ++ | - |
| SCT7 * | +++ | ++ | SCT1 * | +++ | ++ |
| SCT9 * | +++ | ++ | SCT3 | + | - |
| ST2 | ++ | - | ST7 * | +++ | + |
| ST9 * | +++ | + | ST6 * | +++ | ++ |
| LAB Strain | L. monocytogenes DSM 112142 | S. aureus CECT 520 | C. perfringens CECT 821 |
|---|---|---|---|
| CPN1.1 | 15 ± 0.5 | 8 ± 0.5 | 10 ± 0.5 |
| BPF1 | 16 ± 1 | 9 ± 1 | 11 ± 0.5 |
| BPF2 | 24 ± 0.5 | 16 ± 0.4 | 18 ± 0.5 |
| CCH2 | 18 ± 0.6 | 8 ± 0.5 | 12 ± 1 |
| CCH9 | 19 ± 1 | 8 ± 1 | 10 ± 0.5 |
| CCH11 | 16 ± 0.5 | 10 ± 0.5 | 12 ± 1 |
| SCT7 | 19 ± 1 | 12 ± 1 | 11 ± 0.5 |
| SCT9 | 20 ± 0.4 | 14 ± 0.5 | 13 ± 0.5 |
| ST9 | 18 ± 1 | 6 ± 0.4 | 10 ± 1 |
| SCT1 | 19 ± 0.5 | 9 ± 0.5 | 9 ± 1 |
| ST7 | 20 ± 0.5 | 10 ± 0.4 | 12 ± 0.5 |
| ST6 | 24 ± 0.4 | 13 ± 0.4 | 15 ± 0.5 |
| LAB Strain | Protein-Coding DNA Sequences (CDS) | tRNA Genes | rRNA Operons Predicted |
|---|---|---|---|
| P. acidilactici ST6 | 1892 | 55 | 2 |
| Lpb. paraplantarum BPF2 | 3326 | 73 | 3 |
| Genome | Total Assigned GO Terms | GO Terms Molecular Function | GO Terms Biological Processes | GO Terms Cellular Components |
|---|---|---|---|---|
| P. acidilactici ST6 | 3683 GO terms in 1290 CDS (75%) | 2063 (56%) | 1199 (32.6%) | 421 (11.4%) |
| Lpb. paraplantarum BPF2 | 5108 GO terms in 1909 CDS (67%) | 2890 (56.9%) | 1647 (32.2%) | 553 (10.8%) |
| Leucocin K from Lpb. paraplantarum BPF2 | Pediocin-PA from P. acidilactici ST6 | ||
|---|---|---|---|
| Treatments | Bacteriocin Activity a | Treatments | Bacteriocin Activity a |
| Control (cell-free filtered) | 24 ± 0.2 | Control (cell-free filtered) | 24 ± 0.0 |
| Heat | Heat | ||
| 60 °C for 30 min | 24 ± 0.2 | 60 °C for 30 min | 24 ± 1 |
| 80 °C for 30 min | 23 ± 0.5 | 80 °C for 30 min | 24 ± 0.5 |
| 100 °C for 30 min | 19 ± 1 | 100 °C for 30 min | 17 ± 0.5 |
| 121 °C for 15 min | - | 121 °C for 15 min | - |
| pH | pH | ||
| 2 | 24 ± 0.5 | 2 | 24 ± 0.5 |
| 3 | 24 ± 1 | 3 | 24 ± 1 |
| 5 | 24 ± 1 | 5 | 24 ± 1 |
| 7 | 24 ± 0.5 | 7 | 24 ± 0.5 |
| 9 | 24 ± 0.2 | 9 | 24 ± 0.5 |
| 10 | 24 ± 0.5 | 10 | 23 ± 0.2 |
| 11 | - | 11 | - |
| Enzymes | Enzymes | ||
| Trypsin | - | Trypsin | - |
| Papain | - | Papain | - |
| Proteinase K | - | Proteinase K | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-López, J.D.; Teso-Pérez, C.; Martín-Platero, A.M.; Peralta-Sánchez, J.M.; Fonollá-Joya, J.; Martínez-Bueno, M.; Baños, A. Lactiplantibacillus paraplantarum BPF2 and Pediococcus acidilactici ST6, Two Bacteriocinogenic Isolated Strains from Andalusian Spontaneous Fermented Sausages. Foods 2023, 12, 2445. https://doi.org/10.3390/foods12132445
García-López JD, Teso-Pérez C, Martín-Platero AM, Peralta-Sánchez JM, Fonollá-Joya J, Martínez-Bueno M, Baños A. Lactiplantibacillus paraplantarum BPF2 and Pediococcus acidilactici ST6, Two Bacteriocinogenic Isolated Strains from Andalusian Spontaneous Fermented Sausages. Foods. 2023; 12(13):2445. https://doi.org/10.3390/foods12132445
Chicago/Turabian StyleGarcía-López, José David, Claudia Teso-Pérez, Antonio Manuel Martín-Platero, Juan Manuel Peralta-Sánchez, Juristo Fonollá-Joya, Manuel Martínez-Bueno, and Alberto Baños. 2023. "Lactiplantibacillus paraplantarum BPF2 and Pediococcus acidilactici ST6, Two Bacteriocinogenic Isolated Strains from Andalusian Spontaneous Fermented Sausages" Foods 12, no. 13: 2445. https://doi.org/10.3390/foods12132445
APA StyleGarcía-López, J. D., Teso-Pérez, C., Martín-Platero, A. M., Peralta-Sánchez, J. M., Fonollá-Joya, J., Martínez-Bueno, M., & Baños, A. (2023). Lactiplantibacillus paraplantarum BPF2 and Pediococcus acidilactici ST6, Two Bacteriocinogenic Isolated Strains from Andalusian Spontaneous Fermented Sausages. Foods, 12(13), 2445. https://doi.org/10.3390/foods12132445