
Extract of Pfaffia glomerata Ameliorates Paroxetine-Induced Sexual Dysfunction in Male Mice and the Characterization of Its Phytoconstituents by UPLC-MS

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Experimental Drugs
2.3. Preparation of Pfaffia glomerata Extract (PGE)
2.4. Determination of Saponin Content
2.5. PGE Characterization by UPLC–MS
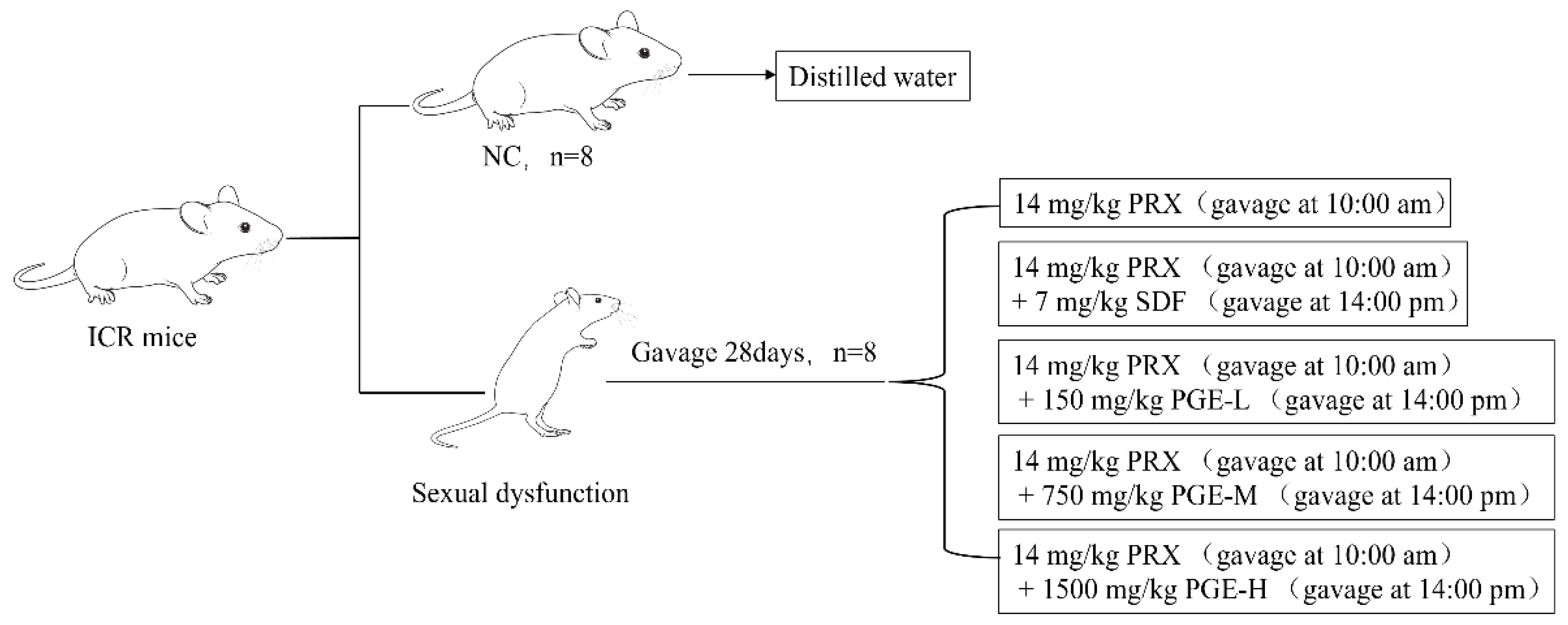
2.6. Animals and Treatment
2.7. Sexual Behavior Experiment
2.8. Organ Coefficient
2.9. Measurements of Hormones and Enzymes
2.10. Testis Histopathological and Sperm Analysis
2.11. Molecular Docking
2.12. Statistical Analysis
3. Results
3.1. Analysis of the Saponin Content of PGE
3.2. UPLC–MS Analysis
3.3. Effect of PGE on the Sexual Behavior of Male Mice
3.4. Effect of PGE on Organ Coefficient
3.5. Effects of PGE on Serum Sex Hormones
3.6. The Effect of PGE on the Activity or Content of NO, NOS, cGMP, and PDE5 in the Penile Tissue
3.7. The Effect of PGE on the Activity and Contents of LDH, ACP, ALP, and MDA in the Testicular Tissue
3.8. Histopathological Analysis
3.9. Effects of Pfaffia Glomerata on Sperm Count and Viability
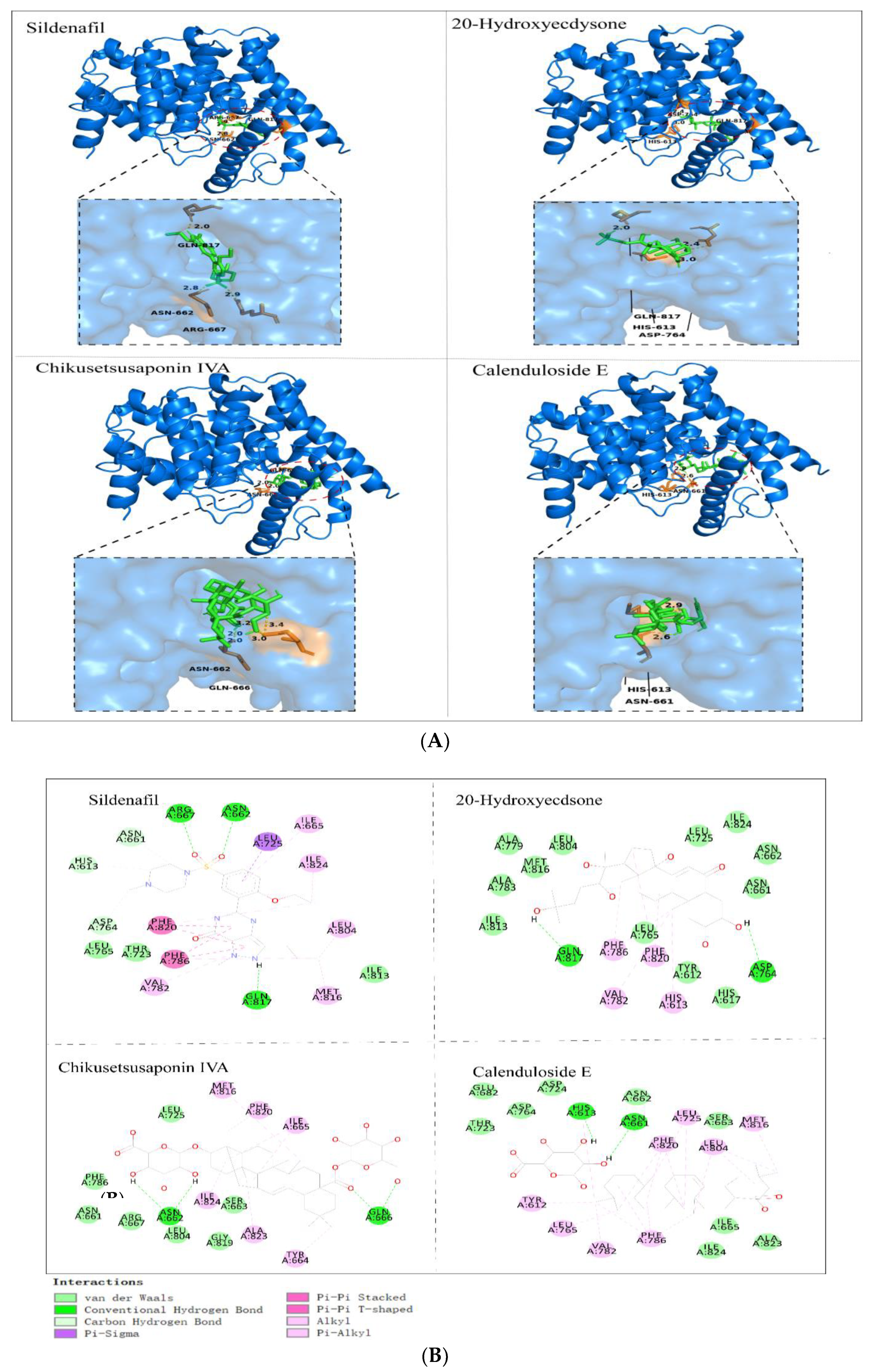
3.10. Molecular Docking Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Khalid, M.; Alqarni, M.H.; Wahab, S.; Annadurai, S.; Alamri, M.A.; Foudah, A.I.; Aljarba, T.M.; Akhtar, J.; Badruddeen; Ahmad, S. Ameliorative Sexual Behavior and Phosphodiesterase-5 Inhibitory Effects of Spondias mangifera Fruit Extract in Rodents: In Silico, In Vitro, and In Vivo Study. J. Clin. Med. 2022, 11, 3732. [Google Scholar] [CrossRef]
- Yakubu, M.T.; Afolayan, A.J. Effect of aqueous extract of Bulbine natalensis (Baker) stem on the sexual behaviour of male rats. Int. J. Androl. 2009, 32, 629–636. [Google Scholar] [CrossRef]
- Durukan, E.; Jensen, C.F.S.; Skaarup, K.G.; Skaarup, K.G.; Østergren, P.B.; Sønksen, J.; Biering-Sørensen, T.; Fode, M. Erectile Dysfunction Is Associated with Left Ventricular Diastolic Dysfunction: A Systematic Review and Meta-analysis. Eur. Urol. Focus 2023, 83, 797–798. [Google Scholar] [CrossRef]
- Pierre, C.J.; Azeez, T.A.; Rossetti, M.L.; Gordon, B.S.; La Favor, J.D. Long-term administration of resveratrol and MitoQ stimulates cavernosum antioxidant gene expression in a mouse castration model of erectile dysfunction. Life Sci. 2022, 310, 121082. [Google Scholar] [CrossRef]
- Gupta, M.; Tiwari, A.; Chandra, K.P.; Awasthi, R.; Chaudhary, S.; Gupta, N.; Agarwal, V.; Chaubey, S.K.; Ansari, S.; Pandey, A.K.; et al. Assessment of erectile dysfunction and other sexual dysfunction in men with type 2 diabetes mellitus: A multicenter observational study in North India. Clin. Epidemiol. Glob. Health 2022, 18, 101136. [Google Scholar] [CrossRef]
- Rahman, A.U.; Alam, F.; Rehman, Z.U.; Khan, M.; Ahmad, T. Effects of Mirabilis jalapa L. root extract and sildenafil on paroxetine-induced sexual dysfunction in male rats and characterization of its phytoconstituents by UPLC-MS. S. Afr. J. Bot. 2023, 152, 240–246. [Google Scholar] [CrossRef]
- Canpolat, S.; Ulker, N.; Yardimci, A.; Tancan, E.; Sahin, E.; Yaman, S.O.; Bulmuş, O.; Alver, A.; Ozcan, M. Irisin ameliorates male sexual dysfunction in paroxetine-treated male rats. Psychoneuroendocrinology 2022, 136, 105597. [Google Scholar] [CrossRef]
- Saikia, Q.; Hazarika, A.; Kalita, J.C. Isoliquiritigenin ameliorates paroxetine-induced sexual dysfunction in male albino mice. Reprod. Toxicol. 2023, 117, 108341. [Google Scholar] [CrossRef]
- Haripriya, V.M.; Dhamotharan, K.; Shukla, S.K.; Suvekbala, V.; Ragupathy, L.; Kumaran, A. Aphrodisiac properties of hydro-alcoholic extract of Cassia auriculata flower in male rats. Andrologia 2019, 51, e13180. [Google Scholar] [CrossRef]
- Masuku, N.P.; Unuofin, J.O.; Lebelo, S.L. Promising role of medicinal plants in the regulation and management of male erectile dysfunction. Biomed. Pharmacother. 2020, 130, 110555. [Google Scholar] [CrossRef]
- Huang, Z.; Huang, L.; Zhou, X.; Liu, H.; Chen, J.; Liu, Y.; Li, B. Chemical constituents from the roots of Pfaddia glomerata. Chin. Tradit. Pat. Med. 2021, 43, 3037–3044. [Google Scholar]
- Yang, Y.; Wang, H.; Yang, J.; Kuang, Y.; Yang, S. Chemical constituents from Pfaffia paniculate. Chin. Tradit. Herb. Drugs 2019, 50, 563–567. [Google Scholar]
- Lu, Q.; Mo, D.; He, C.; Lan, H.; Li, G.; Luo, H.; Mo, H.; Qu, D.; Xu, K. Effects of Pfaffia paniculata Extract on the Growth and Development of Silkworm (Bombyx mori). Genom. Appl. Biol. 2021, 40, 2988–2994. [Google Scholar]
- Dias, F.C.R.; Martins, A.L.P.; de Melo, F.C.S.A.; do Carmo Cupertino, M.; Gomes, M.D.L.M.; De Oliveira, J.M.; Damasceno, E.M.; Silva, J.; Otoni, W.C.; da Matta, S.L.P. Hydroalcoholic extract of Pfaffia glomerata alters the organization of the seminiferous tubules by modulating the oxidative state and the microstructural reorganization of the mice testes. J. Ethnopharmacol. 2019, 233, 179–189. [Google Scholar] [CrossRef]
- Costa, C.A.R.A.; Tanimoto, A.; Quaglio, A.E.V.; Almeida, L.D., Jr.; Severi, J.A.; Di Stasi, L.C. Anti-inflammatory effects of Brazilian ginseng (Pfaffia paniculata) on TNBS-induced intestinal inflammation: Experimental evidence. Int. Immunopharmacol. 2015, 28, 459–469. [Google Scholar] [CrossRef]
- Rahamouz-Haghighi, S.; Sharafi, A. Antiproliferative assay of suma or Brazilian ginseng (Hebanthe eriantha) methanolic extract on HCT116 and 4T1 cancer cell lines, in vitro toxicity on Artemia salina larvae, and antibacterial activity. Nat. Prod. Res. 2023, 1–5. [Google Scholar] [CrossRef]
- Leal, P.F.; Kfouri, M.B.; Alexandre, F.C.; Fagundes, F.H.; Prado, J.M.; Toyama, M.H.; Meireles, M.A.A. Brazilian Ginseng extraction via LPSE and SFE: Global yields, extraction kinetics, chemical composition and antioxidant activity. J. Supercrit. Fluids 2010, 54, 38–45. [Google Scholar] [CrossRef]
- Zhao, F.; Hou, X.; Wang, C. Discussion on foreign Chinese medicine properties of Hebanthe eriantha (Poir) Pwdersen. Glob. Tradit. Chin. Med. 2022, 15, 925–932. [Google Scholar]
- Vardanega, R.; Carvalho, P.I.; Albarelli, J.Q.; Santos, D.T.; Meireles, M.A.A. Techno-economic evaluation of obtaining Brazilian ginseng extracts in potential production scenarios. Food Bioprod. Process. 2017, 101, 45–55. [Google Scholar] [CrossRef]
- Dias, F.C.R.; Gomes, M.D.L.M.; de Melo, F.C.S.A.; Menezes, T.P.; Martins, A.L.; do Carmo Cupertino, M.; Otoni, W.C.; da Matta, S.L.P. Pfaffia glomerata hydroalcoholic extract stimulates penile tissue in adult Swiss mice. J. Ethnopharmacol. 2020, 261, 113182. [Google Scholar] [CrossRef]
- Fu, Y.; Zhang, H.; Fu, Q. Comparison of Ultrasonic Extraction and Antioxidant Activity of Three Quinoa Saponins. J. Chin. Cereals Oils 2020, 35, 40–47. [Google Scholar]
- Luo, S. A Study on Extraction of Effective Constituents of Pfaffia glomerata Planted in Guangxi and Its Anti-Tumor Effects. Master’s Thesis, Guangxi Medical University, Nanning, China, 2014; pp. 27–28. [Google Scholar]
- Wei, G.; Wei, M.; Chen, Q.; Xu, C.; Xu, J.; Huang, X. Optimization of the extraction and enrichment process and evatuation of the free radical scavenging activity of triterpene sapogenins from Pfaffia glomerata. J. Int. Pharm. Res. 2019, 46, 855–861. [Google Scholar]
- Zhang, C. The Production of Total Sapindus-Saponins from Sapindus Mukurossi Gaertn. Master’s Thesis, Jiangnan University, Wuxi, China, 2012; pp. 12–13. [Google Scholar]
- Wei, G.; Li, F.; Huang, C.; Xu, C. Content Determination of Oleanolic Acid-Type Saponins in Pfaffia paniculata by Colorimetry. China Pharm. 2019, 28, 23–25. [Google Scholar]
- Xu, H.J.; Shi, X.W.; Ji, X.; Du, Y.F.; Zhu, H.; Zhang, L.T. A rapid method for simultaneous determination of triterpenoid saponins in Pulsatilla turczaninovii using microwave-assisted extraction and high performance liquid chromatography–tandem mass spectrometry. Food Chem. 2012, 135, 251–258. [Google Scholar] [CrossRef]
- Chen, L.H.; Zhang, Y.B.; Yang, X.W.; Xu, J.; Wang, Z.J.; Sun, Y.Z.; Xu, W.; Wang, Y.P. Application of UPLC-Triple TOF-MS/MS metabolomics strategy to reveal the dynamic changes of triterpenoid saponins during the decocting process of Asian ginseng and American ginseng. Food Chem. 2023, 424, 136425. [Google Scholar] [CrossRef]
- Mroczek, A.; Kapusta, I.; Stochmal, A.; Janiszowska, W. MS/MS and UPLC-MS profiling of triterpenoid saponins from leaves and roots of four red beet (Beta vulgaris L.) cultivars. Phytochem. Lett. 2019, 30, 333–337. [Google Scholar] [CrossRef]
- Ademosun, A.O.; Adebayo, A.A.; Oboh, G. Anogeissus leiocarpus attenuates paroxetine-induced erectile dysfunction in male rats via enhanced sexual behavior, nitric oxide level and antioxidant status. Biomed. Pharmacother. 2019, 111, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, L.; Yan, C.; Wang, Z.; Sui, W.; Yang, P.; Wang, H.; Li, R.; Li, Y.; Lian, F.; et al. Effects of Small Molecule Polypeptide of Oyster on Sexual Function in Male Mice. Genom. Appl. Biol. 2019, 38, 109–116. [Google Scholar]
- Zhang, X.; Peng, Z.; Zheng, H.; Zhang, C.; Lin, H.; Qin, X. The potential protective effect and possible mechanism of peptides from oyster (Crassostrea hongkongensis) hydrolysate on triptolide-induced testis injury in male mice. Mar. Drugs 2021, 19, 566. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.; Cheng, Y. Effect of Astragalus membranaceus polysaccharide on the serum cytokine levels and spermatogenesis of mice. Int. J. Biol. Macromol. 2019, 140, 771–774. [Google Scholar] [CrossRef]
- Oghbaei, H.; Hamidian, G.; Alipour, M.R.; Alipour, S.; Keyhanmanesh, R. The effect of prolonged dietary sodium nitrate treatment on the hypothalamus-pituitary-gonadal axis and testicular structure and function in streptozotocin-induced diabetic male rats. Food Funct. 2020, 11, 2451–2465. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Y.; Huai, Q.; Cai, J.; Zoraghi, R.; Francis, S.H.; Corbin, J.D.; Robinson, H.; Xin, Z.; Lin, G.; et al. Multiple conformations of phosphodiesterase-5: Implications for enzyme function and drug development. J. Biol. Chem. 2006, 281, 21469–21479. [Google Scholar] [CrossRef]
- Zhu, C.; Peng, H.; Yin, C.; Li, Z.; Li, J.; Chen, K.; Xiao, L.; Nie, J.; Liu, Y. Mechanism of Damnacanthi Officinari Radix on promoting testosterone secretion based on network pharmacology and Leydig cells model. Chin. Tradit. Herb. Drugs 2022, 53, 7430–7439. [Google Scholar]
- Bitencourt, R.G.; Queiroga, C.L.; Junior, Í.M.; Cabral, F.A. Fractionated extraction of saponins from Brazilian ginseng by sequential process using supercritical CO2, ethanol and water. J. Supercrit. Fluids 2014, 92, 272–281. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, F.; Yan, D.; Tang, L.; Xu, L.; Huang, L.; Li, B. Based on multi-index optimization of Pfaffia glomerata compound extraction process and its anti-fatigue research. J. Chin. Med. Mater. 2022, 45, 2452–2456. [Google Scholar]
- Oshima, M.; Gu, Y. Pfaffia paniculata induced changes in plasma esadiol-17 beta, progesterone and testosterone levels in mice. J. Reprod. Dev. 2003, 49, 175–180. [Google Scholar] [CrossRef]
- Hoshino, T.; Narukawa, Y.; Haishima, Y.; Goda, Y.; Kiuchi, F. Two new sulfated oleanan saponins from Achyranthes root. J. Nat. Med. 2013, 67, 386–389. [Google Scholar] [CrossRef]
- Fu, J. Qualitative Analysis of Triterpenoid Saponins from Achyranthes Bidentata and Preliminary Study on Metabolism In Vitro and In Vivo. Master’s Thesis, Anhui University of Traditional Chinese Medicine, Hefei, China, 2019. [Google Scholar]
- Nakamura, S.; Chen, G.; Nakashima, S.; Matsuda, H.; Pei, Y.; Yoshikawa, M. Brazilian natural medicines. IV. New noroleanane-type triterpene and ecdysterone-type sterol glycosides and melanogenesis inhibitors from the roots of Pfaffia glomerata. Chem. Pharm. Bull. 2010, 58, 690–695. [Google Scholar] [CrossRef]
- Ridout, C.L.; Price, K.R.; Parkin, G.; Dijoux, M.G.; Lavaud, C. Saponins from sugar beet and the floc problem. J. Agric. Food Chem. 1994, 42, 279–282. [Google Scholar] [CrossRef]
- Sun, H. The Effect and Mechanism of Sterol in Hippocampus Kuda on Mice Reproductive System. Master’s Thesis, Zhejiang Gongshang University, Hangzhou, China, 2015. [Google Scholar]
- Wu, Y.; Xiao, Y.; Yue, Y.; Zhong, K.; Zhao, Y.; Gao, H. A deep insight into mechanism for inclusion of 2R, 3R-dihydromyricetin with cyclodextrins and the effect of complexation on antioxidant and lipid-lowering activities. Food Hydrocoll. 2020, 103, 105718. [Google Scholar] [CrossRef]
- Wu, Z.; Xu, J.; Ruan, J.; Chen, J.; Li, X.; Yu, Y.; Xie, X.; Tang, J.; Zhang, D.; Li, H. Probing the mechanism of interaction between capsaicin and myofibrillar proteins through multispectral, molecular docking, and molecular dynamics simulation methods. Food Chem. X 2023, 18, 100734. [Google Scholar] [CrossRef] [PubMed]
- Giatti, S.; Di Domizio, A.; Diviccaro, S.; Cioffi, L.; Marmorini, I.; Falvo, E.; Caruso, D.; Contini, A.; Melcangi, R.C. Identification of a novel off-target of paroxetine: Possible role in sexual dysfunction induced by this SSRI antidepressant drug. J. Mol. Struct. 2022, 1268, 133690. [Google Scholar] [CrossRef]
- Oosting, R.S.; Chan, J.S.; Olivier, B.; Banerjee, P. Vilazodone does not inhibit sexual behavior in male rats in contrast to paroxetine: A role for 5-HT1A receptors. Neuropharmacology 2016, 107, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Yakubu, M.T.; Atoyebi, A.R. Brysocarpus coccineus (Schum & Thonn) root reinstates sexual competence and testicular function in paroxetine-induced sexual dysfunction in male Wistar rats. Andrologia 2018, 50, e12980. [Google Scholar]
- Bolnick, J.M.; Kilburn, B.A.; Bolnick, A.D.; Diamond, M.P.; Singh, M.; Hertz, M.; Dai, J.; Armant, D.R. Sildenafil stimulates human trophoblast invasion through nitric oxide and guanosine 3′, 5′-cyclic monophosphate signaling. Fertil. Steril. 2015, 103, 1587–1595.e2. [Google Scholar] [CrossRef]
- Cui, K. Human Tissue Kallikrein 1 Protects against the Development of Erectile Dysfunction in a Rat Model of Hyperhomocysteinemia. Ph.D. Thesis, Huazhong University of Science and Technology, Wuhan, China, 2019. [Google Scholar]
- Arora, S.; Surakiatchanukul, T.; Arora, T.; Cagini, C.; Lupidi, M.; Chhablani, J. Sildenafil in ophthalmology: An update. Surv. Ophthalmol. 2022, 67, 463–487. [Google Scholar] [CrossRef]
- Teixeira, T.M.; Boeff, D.D.; de Oliveira Carvalho, L.; Ritter, M.R.; Konrath, E.L. The traditional use of native 21razilian plants for male sexual dysfunction: Evidence from ethnomedicinal applications, animal models, and possible mechanisms of action. J. Ethnopharmacol. 2023, 318, 116876. [Google Scholar] [CrossRef]
- Yakubu, M.T.; Olutoye, A.F. Aphrodisiac activity of aqueous extract of Anthonotha macrophylla P. Beauv. Leaves in female Wistar rats. J. Integr. Med. 2016, 14, 400–408. [Google Scholar] [CrossRef]
- Chen, J.B.; Li, M.J.; Chen, L.X.; Sun, Y.S. Effects of Raphani Semen on anti-fatigue and pharmacokinetics of Panax ginseng. Chin. Herb. Med. 2019, 11, 308–313. [Google Scholar] [CrossRef]
- Corona, G.; Maggi, M. The role of testosterone in male sexual function. Rev. Endocr. Metab. Disord. 2022, 23, 1159–1172. [Google Scholar] [CrossRef]
- Yakubu, M.T.; Jimoh, R.O. Carpolobia lutea roots restore sexual arousal and performance in paroxetine-induced sexually impaired male rats. Rev. Int. Androl. 2014, 12, 90–99. [Google Scholar] [CrossRef]
- Koyuncu, H.; Serefoglu, E.C.; Ozdemir, A.T.; Hellstrom, W.J. Deleterious effects of selective serotonin reuptake inhibitor treatment on semen parameters in patients with lifelong premature ejaculation. Int. J. Impot. Res. 2012, 24, 171–173. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, Z.; Dang, Y.; Liu, J.; Wu, S.; Wang, Y.; Li, Z. Study on effects of apigenin on activity of testicular marker enzymes in mice. J. Med. Pest Control 2013, 11, 1217–1221. [Google Scholar]
- Zheng, H. Effects of Enzymatic Hydrolysates from Chlamys nobilis and Its Gonad in the Sexual Function on Hemi-Castrated Male Rats. Master’s Thesis, Guangdong Ocean University, Zhanjiang, China, 2021. [Google Scholar]
- Tang, X.; Olatunji, O.J.; Zhou, Y.; Hou, X. In vitro and in vivo aphrodisiac properties of the seed extract from Allium tuberosum on corpus cavernosum smooth muscle relaxation and sexual behavior parameters in male Wistar rats. BMC Complement. Altern. Med. 2017, 17, 510. [Google Scholar] [CrossRef]
- Yakubu, M.T.; Jimoh, R.O. Aqueous extract of Carpolobia lutea root ameliorates paroxetine-induced anti-androgenic activity in male rats. Middle East Fertil. Soc. J. 2015, 20, 192–197. [Google Scholar] [CrossRef]
- Ma, N.; Zhang, C.; Chen, Q.; Liu, Z.; Han, G.; Liu, J.; Yuan, D.; Zhao, H. Protective Effects and Mechanism of Oleanolic Acid on Testicular DNA Damage in Natural Aging Rats. Nat. Prod. Res. Dev. 2018, 30, 767–772+782. [Google Scholar]
- Zhao, H.; Liu, J.; Song, L.; Liu, Z.; Han, G.; Yuan, D.; Wang, T.; Dun, Y.; Zhou, Z.; Liu, Z.; et al. Oleanolic acid rejuvenates testicular function through attenuating germ cell DNA damage and apoptosis via deactivation of NF-κB, p53 and p38 signalling pathways. J. Pharm. Pharmacol. 2017, 69, 295–304. [Google Scholar] [CrossRef]
- Corona, G.; Rastrelli, G.; Vignozzi, L.; Maggi, M. Androgens and male sexual function. Best Pract. Res. Clin. Endocrinol. Metab. 2022, 36, 101615. [Google Scholar] [CrossRef]
- Chu, F.; Zhang, W.; Guo, W.; Wang, Z.; Yang, Y.; Zhang, X.; Fang, K.; Yan, M.; Wang, P.; Lei, H. Oleanolic Acid-Amino Acids Derivatives: Design, Synthesis, and Hepatoprotective Evaluation In Vitro and In Vivo. Molecules 2018, 23, 322. [Google Scholar]
- Qu, Z.; Liu, H.; Zheng, P.; Yao, C.; Wang, Y. Study on pharmacological activity of oleanolic acid saponins. Chin. Tradit. Pat. Med. 2012, 34, 1143–1147. [Google Scholar]
- Zhao, J.; Chen, Z.; Tan, Z.; Li, A.; Xi, Y. Protective effect of oleanolic acid liposomes on cisplatin-induced oligoasthenospermia in mice. Natl. J. Androl. 2022, 28, 970–975. [Google Scholar]
- El-Gaafarawi, I.; Hassan, M.; Fouad, G.; El-Komey, F. Toxic effects of paroxetine on sexual and reproductive functions of rats. Egypt. J. Hosp. Med. 2005, 21, 16–32. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | tR/min | [M-H]− | [M+COOH]− | Formula | Fragmentation Ions | Inferred Compounds | Reference |
|---|---|---|---|---|---|---|---|
| 1 | 0.82 | 479.3025 | C27H44O7 | 319.1981; 159.1127 | 20-Hydroxyecdysone | # | |
| 2 | 4.17 | 1080.3845 | C47H70O23S | 955.4741; 793.4250; 631.3748 | Sulfachyranthoside B | [39] | |
| 3 | 4.45 | 777.3987 | C41H62O14 | 631.3800; 455.3551 | 3-O-β-D-Glc-(1→2)-[[2-carboxy-1-(carboxymethyl)-2-hydroxyethyl-(1→3)]-β-D-GluA oleanolic acid | [40] | |
| 4 | 4.72 | 925.4672 | C46H70O19 | 793.4254; 631.3765; 455.3594 | Achyranthoside E | [40] | |
| 5 | 4.82 | 793.4303 | C42H66O14 | 631.3814; 569.3831; 455.3549 | Chikusetsusaponin IVA | # | |
| 6 | 5.16 | 631.3459 | C35H52O10 | 455.3197; 384.9140; 146.9766 | Pfaffiaglycoside B | [41] | |
| 7 | 5.71 | 793.4293 | C42H66O14 | 631.3812; 569.3831; 455.3578 | Zingibroside R1 | [40] | |
| 8 | 7.08 | 955.4397 | C48H76O19 | 793.4292; 631.3809; 569.3834; 455.3556 | Achyranthoside C | [41] | |
| 9 | 8.04 | 631.3823 | C36H56O9 | 455.3552; 146.9765 | Calenduloside E | [42] |
| Item | Heart | Thymus | Lien | Liver | Ren | Penis | Testis |
|---|---|---|---|---|---|---|---|
| CN | 0.546 ± 0.031 | 0.081 ± 0.026 | 0.31 ± 0.017 | 4.442 ± 0.250 | 1.535 ± 0.125 | 0.126 ± 0.008 | 0.769 ± 0.056 |
| PRX | 0.544 ± 0.035 | 0.081 ± 0.013 | 0.307 ± 0.065 | 4.267 ± 0.350 | 1.512 ± 0.076 | 0.120 ± 0.005 | 0.677 ± 0.042 ** |
| PRX + SDF | 0.565 ± 0.034 | 0.081 ± 0.024 | 0.304 ± 0.033 | 4.441 ± 0.239 | 1.515 ± 0.071 | 0.124 ± 0.011 | 0.738 ± 0.042 # |
| PRX + PGE-L | 0.536 ± 0.033 | 0.081 ± 0.024 | 0.302 ± 0.044 | 4.314 ± 0.65 | 1.530 ± 0.035 | 0.123 ± 0.011 | 0.737 ± 0.056 # |
| PRX + PGE-M | 0.555 ± 0.038 | 0.086 ± 0014 | 0.303 ± 0.018 | 4.445 ± 0.295 | 1.536 ± 0.075 | 0.125 ± 0.009 | 0.760 ± 0.051 ## |
| PRX + PGE-H | 0.557 ± 0.043 | 0.082 ± 0.017 | 0.302 ± 0.031 | 4.361 ± 0.493 | 1.528 ± 0.059 | 0.124 ± 0.013 | 0.757 ± 0.046 ## |
| Ligands | Binding Energy Score (kcal/mol) | H-Bond Interaction | Interactions |
|---|---|---|---|
| Sildenafil | −9.0 | GLN A:817, ARG A:667, ASN A:662 | HIS A:613, ASN A:661, LEU A:725, ILE A:824, LEU A:804, ILE A:813, MET A:816, VAI A:782, PHE A:786; THR A:723, LEU A:765, ASP A:764, PHE A:820 |
| 20-Hydroxyecdysone | −8.3 | GLN A:817, ASP A:764 | ILE A:813, ALA A:783, ALA A:779, MET A:816, LEU A:804, LEU A:725, ILE A:824, ASN A:662, ASN A:661, HIS A:617, TYR A:612, LEU A:765 |
| Sulfachyranthoside B | −8.5 | GLN A:666, SER A:663 | TYR A:664, THR A:802, ASN A:662, LEU A:804, LEU A:725, ILE A:824, PHE A:820, MET A:816, ALA A:823, GLY A:819, SER A:815 |
| Achyranthoside E | −9.1 | GLN A:817, SER A:815 | GLY A:819, MET A:816, ILE A:665, ILE A:824, PHE A:820, ILE A:813, LEU A:804, PHE A:786, LEU A:765, VAL A:782, GLN A:775, ALA A:767, ILE A:778, ILE A:768, ALA A:779 TYR A:612 HIS A:613, SER A:663 ASP A:803, LYS A:812 |
| Chikusetsusaponin IVA | −8.6 | ASN A:662, GLN A:666 | LEU A:725, MET A:816, PHE A:820, ILE A:665, TYR A:664, ALA A:823, SER A:663, ILE A:824, GLY A:819, LEU A:804, ARG A:667, ASN A:661, PHE A:786 |
| Pfaffiaglycoside B | −10.3 | HIS A:613 | ALA A:823; GLY A:819, ILE A:665, ILE A:824, PHE A:820, PHE A:786; LEU A:765, VAL A:782, TYR A:612, THR A:723, ASP A:764, ASP A:724, GLU A:682, HIS A:657, ASN A:661, LEU A:725, LEU A:804, MET A:816 |
| Zingibroside R1 | −10.7 | VAL A:660, ARH A:667, ASN A:662, HIS A:613, GLU A:682 | ILE A:689, SER A:679, MET A:681, ASP A:764, THR A:723, ASP A:724, ILE A:665, SER A:663, LEU A:725, MET A:816, ALA A:824, LEU A:804, PHE A:820, VAL A:782, PHE A:786, ASN A:661 |
| Achyranthoside C | −8.8 | GLN A:817, TYR A:612, ALA A:823 | GLY A:819, ASN A:662, LEU A:725, SER A:663, ASN A:661, PHE A:786, VAL A:782, GLN A:775, ILE A:786, ALA A:779, ALA A:767, HIS A:617, LEU A:765, LEU A:804, PHE A:820, MET A:816, ASP A:764, ILE A:665, ILE A:824, GLN A:666, ILE A:729 |
| Calenduloside E | −10.0 | ASN A:662, HIS A:613 | THR A:723, GLU A:682, ASP A:764, ASP A:724, ASN A:662, PHE A:820, LEU A:725, SER A:663, MET A:816, LEU A:804, ALA A:823, ILE A:824, ILE A:665, PHE A:786, VAL 782, LEU A:765, TYR A:612 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Q.; Wu, H.; Qin, X. Extract of Pfaffia glomerata Ameliorates Paroxetine-Induced Sexual Dysfunction in Male Mice and the Characterization of Its Phytoconstituents by UPLC-MS. Foods 2023, 12, 3236. https://doi.org/10.3390/foods12173236
Huang Q, Wu H, Qin X. Extract of Pfaffia glomerata Ameliorates Paroxetine-Induced Sexual Dysfunction in Male Mice and the Characterization of Its Phytoconstituents by UPLC-MS. Foods. 2023; 12(17):3236. https://doi.org/10.3390/foods12173236
Chicago/Turabian StyleHuang, Qianqian, Haiying Wu, and Xiaoming Qin. 2023. "Extract of Pfaffia glomerata Ameliorates Paroxetine-Induced Sexual Dysfunction in Male Mice and the Characterization of Its Phytoconstituents by UPLC-MS" Foods 12, no. 17: 3236. https://doi.org/10.3390/foods12173236
APA StyleHuang, Q., Wu, H., & Qin, X. (2023). Extract of Pfaffia glomerata Ameliorates Paroxetine-Induced Sexual Dysfunction in Male Mice and the Characterization of Its Phytoconstituents by UPLC-MS. Foods, 12(17), 3236. https://doi.org/10.3390/foods12173236