
Role of Anthocyanins in the Interaction between Salivary Mucins and Wine Astringent Compounds
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
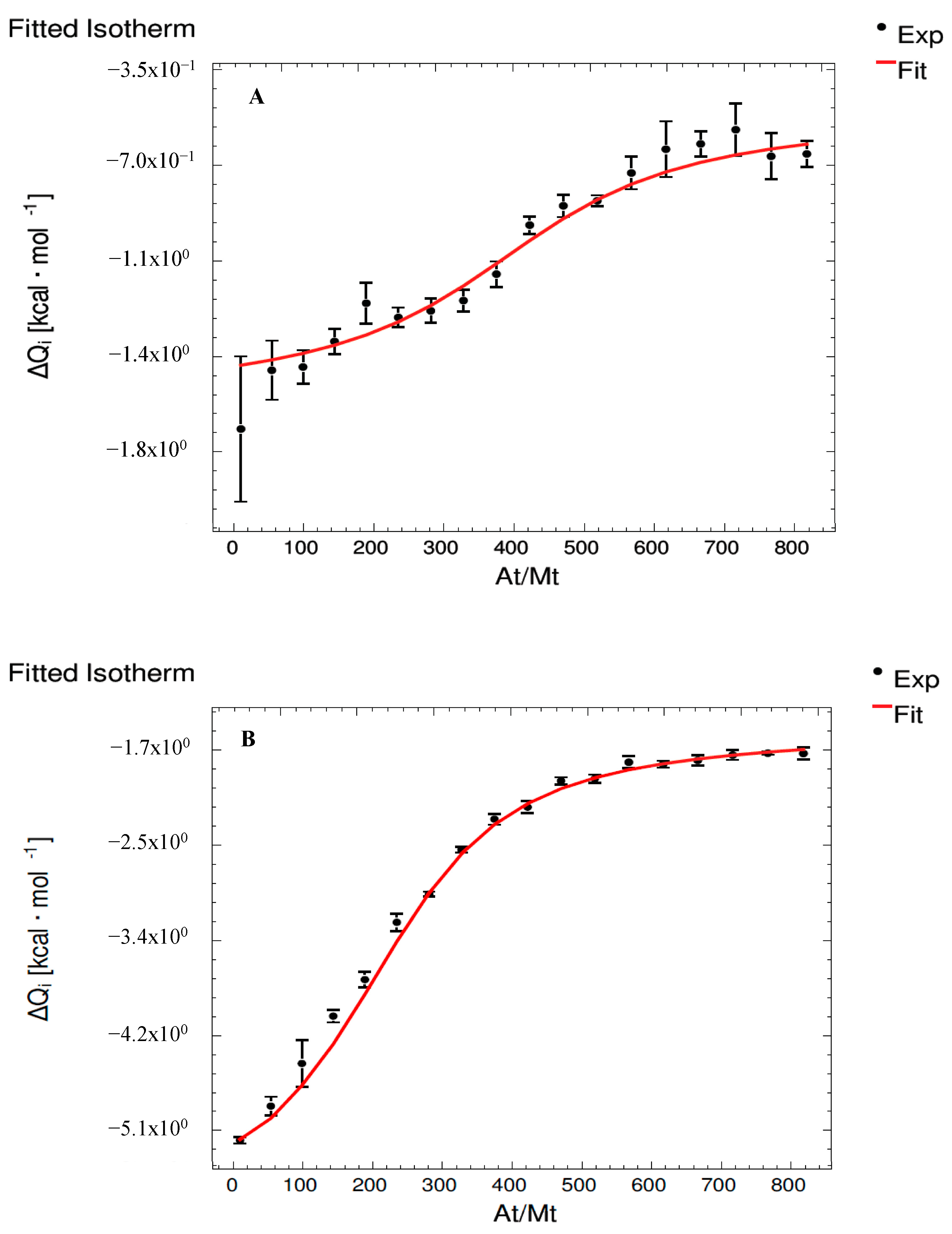
3.1. Interaction Assays Corresponding to Catechin System
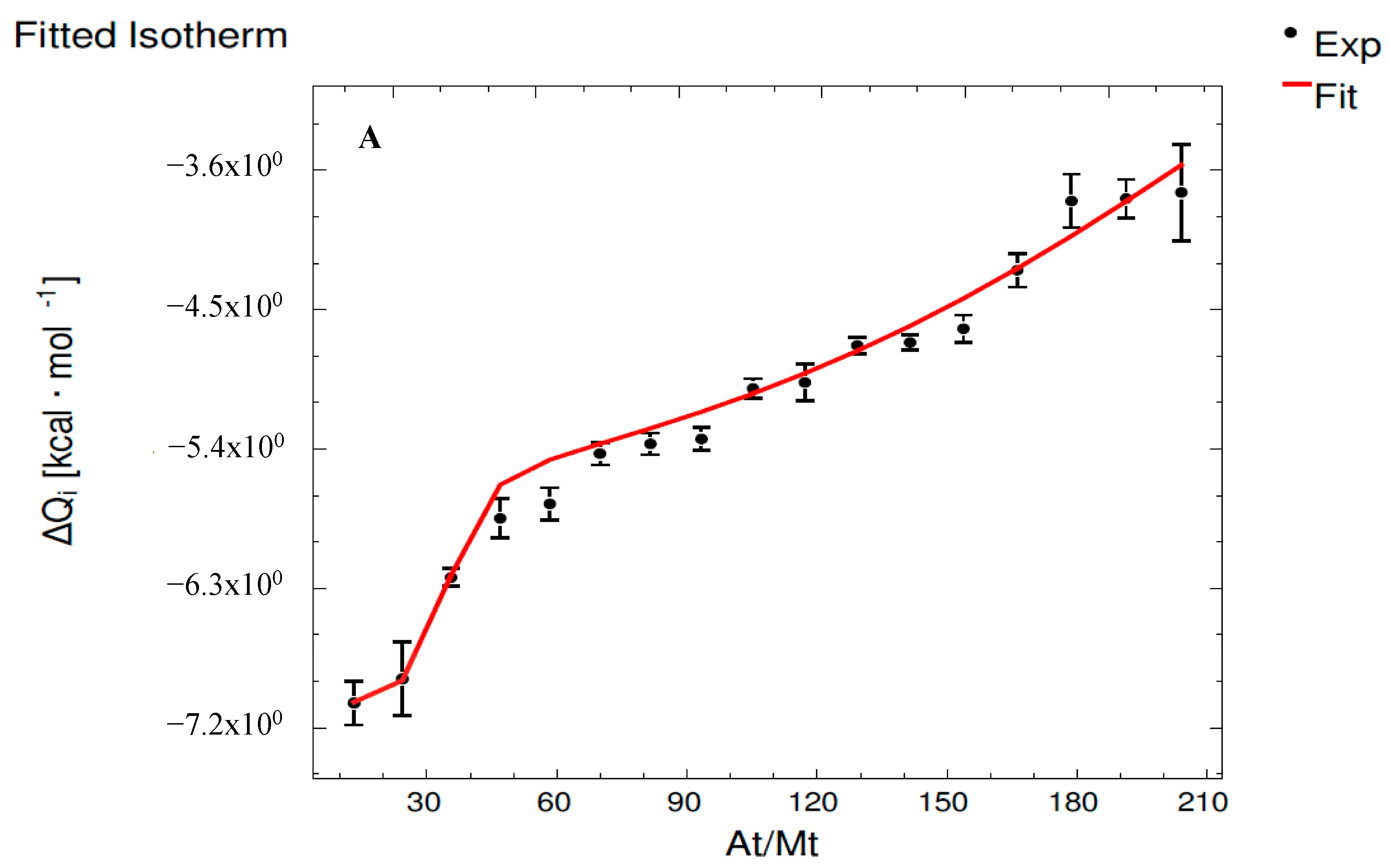
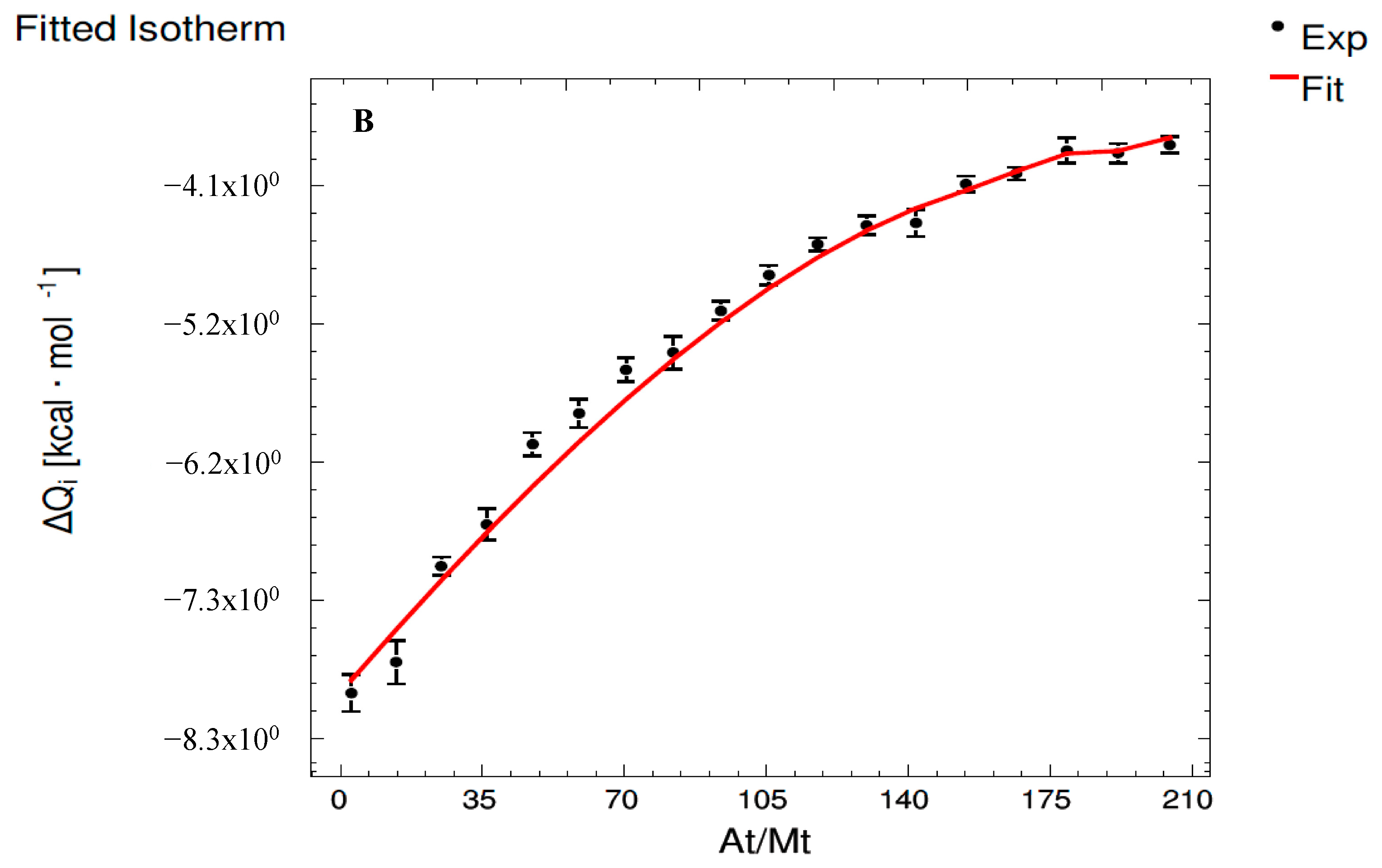
3.2. Interaction Assays Corresponding to Epicatechin System
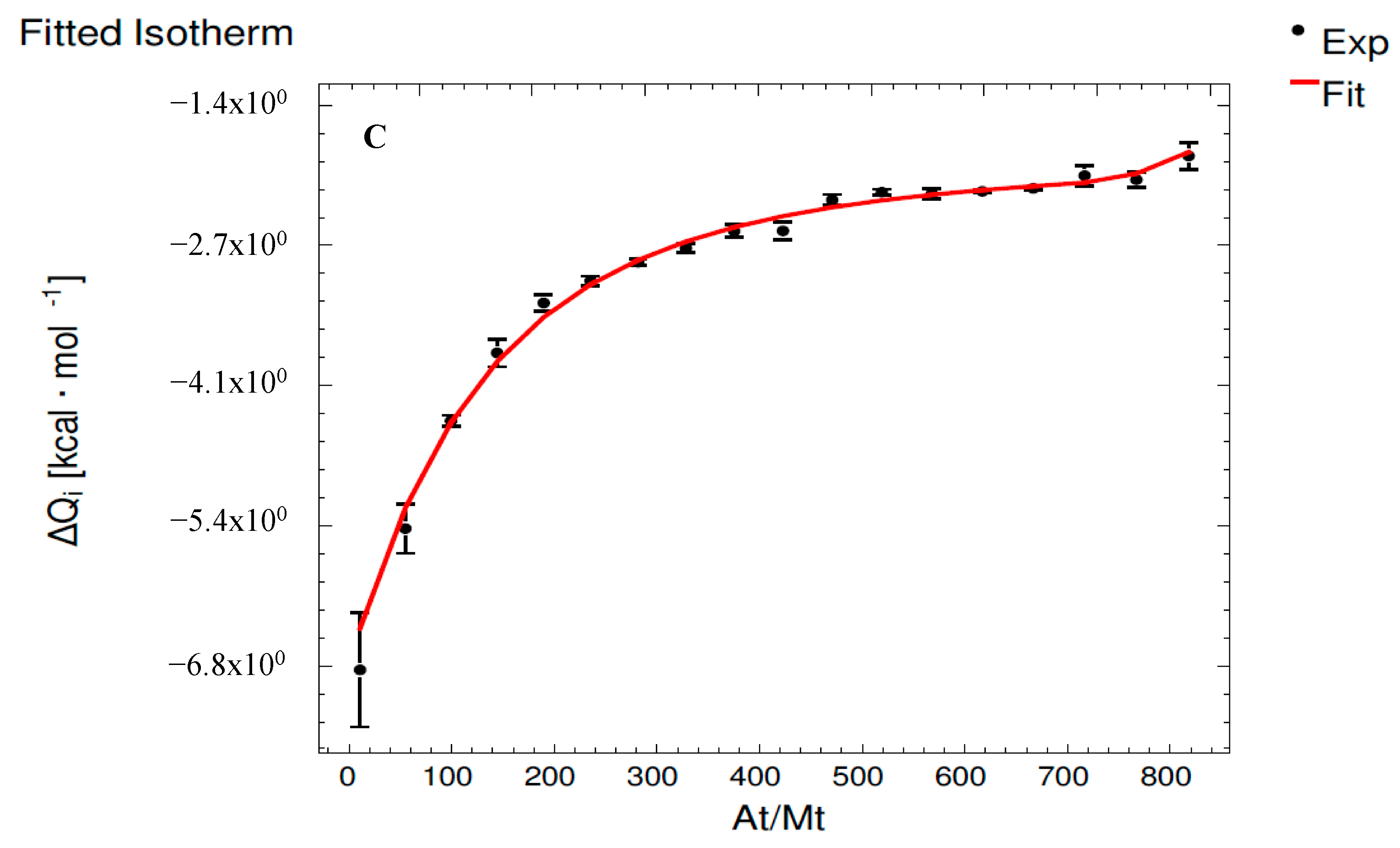
3.3. Interaction Assays Corresponding to QG System
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Mira de Orduña, R. Climate Change Associated Effects on Grape and Wine Quality and Production. Food Res. Int. 2010, 43, 1844–1855. [Google Scholar] [CrossRef]
- Boulton, R. The Copigmentation of Anthocyanins and Its Role in the Color of Red Wine: A Critical Review. Am. J. Enol. Vitic. 2001, 52, 67–87. [Google Scholar] [CrossRef]
- He, F.; Liang, N.-N.; Mu, L.; Pan, Q.-H.; Wang, J.; Reeves, M.J.; Duan, C.-Q. Anthocyanins and Their Variation in Red Wines I. Monomeric Anthocyanins and Their Color Expression. Molecules 2012, 17, 1571–1601. [Google Scholar] [CrossRef] [PubMed]
- González-Manzano, S.; Dueñas, M.; Rivas-Gonzalo, J.C.; Escribano-Bailón, M.T.; Santos-Buelga, C. Studies on the Copigmentation between Anthocyanins and Flavan-3-Ols and Their Influence in the Colour Expression of Red Wine. Food Chem. 2009, 114, 649–656. [Google Scholar] [CrossRef]
- Brouillard, R.; Wigand, M.-C.; Dangles, O.; Cheminat, A. PH and Solvent Effects on the Copigmentation Reaction of Malvin with Polyphenols, Purine and Pyrimidine Derivatives. J. Chem. Soc. Perkin Trans. 1991, 2, 1235–1241. [Google Scholar] [CrossRef]
- Gawel, R. Red Wine Astringency: A Review. Aust. J. Grape Wine Res. 1998, 4, 74–95. [Google Scholar] [CrossRef]
- Soares, S.; Brandão, E.; Guerreiro, C.; Soares, S.; Mateus, N.; de Freitas, V. Tannins in Food: Insights into the Molecular Perception of Astringency and Bitter Taste. Molecules 2020, 25, 2590. [Google Scholar] [CrossRef]
- Baxter, N.J.; Lilley, T.H.; Haslam, E.; Williamson, M.P. Multiple Interactions between Polyphenols and a Salivary Proline-Rich Protein Repeat Result in Complexation and Precipitation. Biochemistry 1997, 36, 5566–5577. [Google Scholar] [CrossRef]
- Ferrer-Gallego, R.; Soares, S.; Mateus, N.; Rivas-Gonzalo, J.; Escribano-Bailón, M.T.; De Freitas, V. New Anthocyanin-Human Salivary Protein Complexes. Langmuir 2015, 31, 8392–8401. [Google Scholar] [CrossRef]
- Soares, S.; Santos Silva, M.; García-Estévez, I.; Brandão, E.; Fonseca, F.; Ferreira-da-Silva, F.; Escribano-Bailón, M.T.; Mateus, N.; de Freitas, V. Effect of Malvidin-3-Glucoside and Epicatechin Interaction on Their Ability to Interact with Salivary Proline-Rich Proteins. Food Chem. 2019, 276, 33–42. [Google Scholar] [CrossRef]
- Huq, N.L.; Cross, K.J.; Ung, M.; Myroforidis, H.; Veith, P.D.; Chen, D.; Stanton, D.; He, H.; Ward, B.R.; Reynolds, E.C. A Review of the Salivary Proteome and Peptidome and Saliva-Derived Peptide Therapeutics. Int. J. Pept. Res. Ther. 2007, 13, 547–564. [Google Scholar] [CrossRef]
- Dinu, V.; Gillis, R.B.; MacCalman, T.; Lim, M.; Adams, G.G.; Harding, S.E.; Fisk, I.D. Submaxillary Mucin: Its Effect on Aroma Release from Acidic Drinks and New Insight into the Effect of Aroma Compounds on Its Macromolecular Integrity. Food Biophys. 2019, 14, 278–286. [Google Scholar] [CrossRef]
- van Nieuw Amerongen, A.; Bolscher, J.G.M.; Veerman, E.C.I. Salivary Proteins: Protective and Diagnostic Value in Cariology? Caries Res. 2004, 38, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Bansil, R.; Turner, B.S. Mucin Structure, Aggregation, Physiological Functions and Biomedical Applications. Curr. Opin. Colloid. Interface Sci. 2006, 11, 164–170. [Google Scholar] [CrossRef]
- Dhanisha, S.S.; Guruvayoorappan, C.; Drishya, S.; Abeesh, P. Mucins: Structural Diversity, Biosynthesis, Its Role in Pathogenesis and as Possible Therapeutic Targets. Crit. Rev. Oncol. Hematol. 2018, 122, 98–122. [Google Scholar] [CrossRef]
- Davies, H.S.; Pudney, P.D.A.; Georgiades, P.; Waigh, T.A.; Hodson, N.W.; Ridley, C.E.; Blanch, E.W.; Thornton, D.J. Reorganisation of the Salivary Mucin Network by Dietary Components: Insights from Green Tea Polyphenols. PLoS ONE 2014, 9, e108372. [Google Scholar] [CrossRef]
- Brandão, E.; Santos Silva, M.; García-Estévez, I.; Mateus, N.; de Freitas, V.; Soares, S. Molecular Study of Mucin-Procyanidin Interaction by Fluorescence Quenching and Saturation Transfer Difference (STD)-NMR. Food Chem. 2017, 228, 427–434. [Google Scholar] [CrossRef]
- Nayak, A.; Carpenter, G.H. A Physiological Model of Tea-Induced Astringency. Physiol. Behav. 2008, 95, 290–294. [Google Scholar] [CrossRef]
- Lee, C.A.; Vickers, Z.M. Astringency of Foods May Not Be Directly Related to Salivary Lubricity. J. Food Sci. 2012, 77, 302–306. [Google Scholar] [CrossRef]
- Frazier, R.A.; Deaville, E.R.; Green, R.J.; Stringano, E.; Willoughby, I.; Plant, J.; Mueller-Harvey, I. Interactions of Tea Tannins and Condensed Tannins with Proteins. J. Pharm. Biomed. Anal. 2010, 51, 490–495. [Google Scholar] [CrossRef]
- McRae, J.M.; Falconer, R.J.; Kennedy, J.A. Thermodynamics of Grape and Wine Tannin Interaction with Polyproline: Implications for Red Wine Astringency. J. Agric. Food Chem. 2010, 58, 12510–12518. [Google Scholar] [CrossRef] [PubMed]
- Poncet-Legrand, C.; Gautier, C.; Cheynier, V.; Imberty, A. Interactions between Flavan-3-Ols and Poly(L-Proline) Studied by Isothermal Titration Calorimetry: Effect of the Tannin Structure. J. Agric. Food Chem. 2007, 55, 9235–9240. [Google Scholar] [CrossRef] [PubMed]
- De Freitas, V.; Mateus, N. Protein/Polyphenol Interactions: Past and Present Contributions. Mechanisms of Astringency Perception. Curr. Org. Chem. 2012, 16, 724–746. [Google Scholar] [CrossRef]
- Cala, O.; Pinaud, N.; Simon, C.; Fouquet, E.; Laguerre, M.; Dufourc, E.J.; Pianet, I. NMR and Molecular Modeling of Wine Tannins Binding to Saliva Proteins: Revisiting Astringency from Molecular and Colloidal Prospects. FASEB J. 2010, 24, 4281–4290. [Google Scholar] [CrossRef]
- Ramos-Pineda, A.M.; García-Estévez, I.; Brás, N.F.; Martín Del Valle, E.M.; Dueñas, M.; Escribano Bailón, M.T. Molecular Approach to the Synergistic Effect on Astringency Elicited by Mixtures of Flavanols. J. Agric. Food Chem. 2017, 65, 6425–6433. [Google Scholar] [CrossRef]
- Soares, S.; García-Estévez, I.; Ferrer-Galego, R.; Brás, N.F.; Brandão, E.; Silva, M.; Teixeira, N.; Fonseca, F.; Sousa, S.F.; Ferreira-da-Silva, F.; et al. Study of Human Salivary Proline-Rich Proteins Interaction with Food Tannins. Food Chem. 2018, 243, 175–185. [Google Scholar] [CrossRef]
- Mao, M.; Li, K.; Liao, M.; Chen, F.; Hu, X.; Ma, L.; Ji, J. Study on the Interactions Between Oral Mucin and Cyanidin 3-O-Glucoside: The Effect of Oxidized Quinone. Food Bioprocess Technol. 2023. [Google Scholar] [CrossRef]
- García-Estévez, I.; Alcalde-Eon, C.; Escribano-Bailón, M.T. Flavanol Quantification of Grapes via Multiple Reaction Monitoring Mass Spectrometry. Application to Differentiation among Clones of Vitis vinifera L. Cv. Rufete Grapes. J. Agric. Food Chem. 2017, 65, 6359–6368. [Google Scholar] [CrossRef]
- García-Estévez, I.; Jacquet, R.; Alcalde-Eon, C.; Rivas-Gonzalo, J.C.; Escribano-Bailón, M.T.; Quideau, S. Hemisynthesis and Structural and Chromatic Characterization of Delphinidin 3-O-Glucoside−Vescalagin Hybrid Pigment. J. Agric. Food Chem. 2013, 61, 11560–11568. [Google Scholar] [CrossRef]
- Watrelot, A.A.; Renard, C.M.G.C.; Le Bourvellec, C. Comparison of Microcalorimetry and Haze Formation to Quantify the Association of B-Type Procyanidins to Poly-L-Proline and Bovine Serum Albumin. LWT—Food Sci. Technol. 2015, 63, 376–382. [Google Scholar] [CrossRef]
- McRae, J.M.; Ziora, Z.M.; Kassara, S.; Cooper, M.A.; Smith, P.A. Ethanol Concentration Influences the Mechanisms of Wine Tannin Interactions with Poly(L-proline) in Model Wine. J. Agric. Food Chem. 2015, 63, 4345–4352. [Google Scholar] [CrossRef] [PubMed]
- Frazier, R.A.; Papadopoulou, A.; Green, R.J. Isothermal Titration Calorimetry Study of Epicatechin Binding to Serum Albumin. J. Pharm. Biomed. Anal. 2006, 41, 1602–1605. [Google Scholar] [CrossRef] [PubMed]
- Manjón, E.; Brás, N.F.; García-Estévez, I.; Escribano-Bailón, M.T. Cell Wall Mannoproteins from Yeast Affect Salivary Protein-Flavanol Interactions through Different Molecular Mechanisms. J. Agric. Food Chem. 2020, 68, 13459–13468. [Google Scholar] [CrossRef]
- De Freitas, V.; Carvalho, E.; Mateus, N. Study of Carbohydrate Influence on Protein-Tannin Aggregation by Nephelometry. Food Chem. 2003, 81, 503–509. [Google Scholar] [CrossRef]
- Ferrer-Gallego, R.; Quijada-Morín, N.; Brás, N.F.; Gomes, P.; de Freitas, V.; Rivas-Gonzalo, J.C.; Escribano-Bailón, M.T. Characterization of Sensory Properties of Flavanols—A Molecular Dynamic Approach. Chem. Senses 2015, 40, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Torres-Rochera, B.; Manjón, E.; Brás, N.F.; Escribano-Bailón, M.T.; García-Estévez, I. Supramolecular Study of the Interactions between Malvidin-3-O-Glucoside and Wine Phenolic Compounds: Influence on Color. J. Agric. Food Chem. 2023. [Google Scholar] [CrossRef] [PubMed]
- Ferrer-Gallego, R.; Brás, N.F.; García-Estévez, I.; Mateus, N.; Rivas-Gonzalo, J.C.; De Freitas, V.; Escribano-Bailón, M.T. Effect of Flavonols on Wine Astringency and Their Interaction with Human Saliva. Food Chem. 2016, 209, 358–364. [Google Scholar] [CrossRef]
- Guerreiro, C.; Brandão, E.; de Jesus, M.; Gonçalves, L.; Pérez-Gregório, R.; Mateus, N.; de Freitas, V.; Soares, S. New Insights into the Oral Interactions of Different Families of Phenolic Compounds: Deepening the Astringency Mouthfeels. Food Chem. 2022, 375, 131642. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Set 1 | MC | MMvC (C) | MMvC (Mv) | MMv2000 |
| K1 (M−1) | (1.92 ± 0.21) × 109 b | (5.11 ± 0.21) × 107 c | (1.97 ± 0.08) × 107 c | (3.22 ± 0.03) × 1010 a |
| ΔH1 (cal·mol−1) | (−1.02 ± 0.02) × 103 b | (1.00 ± 0.06) × 105 a | (−4.03 ± 0.13) × 105 c | (−6.01 ± 0.01) × 103 b |
| ΔG1 (cal·mol−1) | (−1.26 ± 0.13) × 104 a,b | (−1.05 ± 0.04) × 104 a | (−9.93 ± 0.40) × 103 a | (−1.43 ± 0.01) × 104 b |
| −T·ΔS1 (cal·mol−1) | (−1.16 ± 0.14) × 104 b | (−1.10 ± 0.06) × 105 c | (3.93 ± 0.13) × 105 a | (−8.30 ± 0.14) × 103 b |
| Set 2 | MC | MMvC (C) | MMvC (Mv) | MMv2000 |
| K2 (M−1) | (6.09 ± 0.08) × 107 b | (1.00 ± 0.01) × 107 c | (1.02 ± 0.02) × 107 c | (8.92 ± 0.20) × 108 a |
| ΔH2 (cal·mol−1) | (−1.55 ± 0.10) × 102 b | (−1.13 ± 0.01) × 105 d | (9.10 ± 0.01) × 104 a | (−1.36 ± 0.01) × 103 c |
| ΔG2 (cal·mol−1) | (−1.06 ± 0.01) × 104 b | (−9.53 ± 0.09) × 103 a | (−9.54 ± 0.19) × 103 a | (−1.22 ± 0.03) × 104 c |
| −T·ΔS2 (cal·mol−1) | (−1.04 ± 0.01) × 104 b | (1.04 ± 0.01) × 105 a | (−1.01 ± 0.01) × 105 c | (−1.08 ± 0.03) × 104 b |
| Set 1 | ME | MMvE (E) | MMvE (Mv) | MMv500 |
| K1 (M−1) | - | (9.29 ± 0.35) × 108 a | (6.97 ± 0.15) × 108 b | (6.20 ± 1.70) × 107 c |
| ΔH1 (cal·mol−1) | - | (2.03 ± 0.10) × 105 a | (1.37 ± 0.10) × 105 b | (−6.61 ± 0.07) × 103 c |
| ΔG1 (cal·mol−1) | - | (−1.22 ± 0.04) × 104 a | (−1.20 ± 0.03) × 104 a | (−1.06 ± 0.29) × 104 a |
| −T·ΔS1 (cal·mol−1) | - | (−2.16 ± 0.10) × 105 c | (−1.49 ± 0.10) × 105 b | (−4.01 ± 2.98) × 103 a |
| Set 2 | ME | MMvE (E) | MMvE (Mv) | MMv500 |
| K2 (M−1) | - | (6.06 ± 0.14) × 108 a | (3.16 ± 0.10) × 108 b | (9.49 ± 0.76) × 104 c |
| ΔH2 (cal·mol−1) | - | (−2.76 ± 0.03) × 105 b | (−3.57 ± 0.18) × 105 c | (−5.66 ± 0.12) × 103 a |
| ΔG2 (cal·mol−1) | - | (−1.20 ± 0.03) × 104 b | (−1.16 ± 0.04) × 104 b | (−6.78 ± 0.54) × 103 a |
| −T·ΔS2 (cal·mol−1) | - | (2.64 ± 0.03) × 105 b | (3.46 ± 0.18) × 105 a | (−1.12 ± 0.66) × 103 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Rochera, B.; Manjón, E.; Escribano-Bailón, M.T.; García-Estévez, I. Role of Anthocyanins in the Interaction between Salivary Mucins and Wine Astringent Compounds. Foods 2023, 12, 3623. https://doi.org/10.3390/foods12193623
Torres-Rochera B, Manjón E, Escribano-Bailón MT, García-Estévez I. Role of Anthocyanins in the Interaction between Salivary Mucins and Wine Astringent Compounds. Foods. 2023; 12(19):3623. https://doi.org/10.3390/foods12193623
Chicago/Turabian StyleTorres-Rochera, Bárbara, Elvira Manjón, María Teresa Escribano-Bailón, and Ignacio García-Estévez. 2023. "Role of Anthocyanins in the Interaction between Salivary Mucins and Wine Astringent Compounds" Foods 12, no. 19: 3623. https://doi.org/10.3390/foods12193623
APA StyleTorres-Rochera, B., Manjón, E., Escribano-Bailón, M. T., & García-Estévez, I. (2023). Role of Anthocyanins in the Interaction between Salivary Mucins and Wine Astringent Compounds. Foods, 12(19), 3623. https://doi.org/10.3390/foods12193623