
Succession and Diversity of Microbial Flora during the Fermentation of Douchi and Their Effects on the Formation of Characteristic Aroma


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
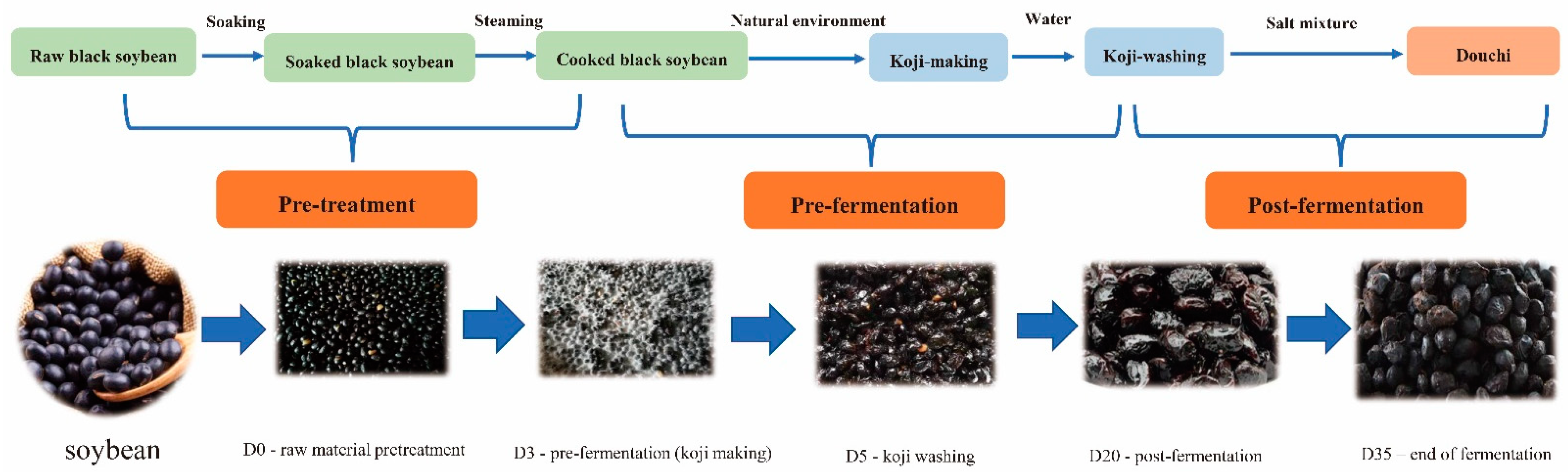
2.1. Collection of Douchi Samples
2.2. Extraction and Sequencing of DNA
2.2.1. DNA Extraction from Bacteria and Fungi
2.2.2. Amplification and Sequencing by Illumina MiSeq
2.2.3. Bioinformatics Analysis
2.3. Analysis of Volatile Compounds in Douchi
2.3.1. Isolation of Volatile Compounds from Traditional Fermented Douchi
2.3.2. Using Gas Chromatography–Mass Spectrometry to Identify and Quantify Volatile Compounds
2.4. Statistical Analysis
3. Results and Discussion
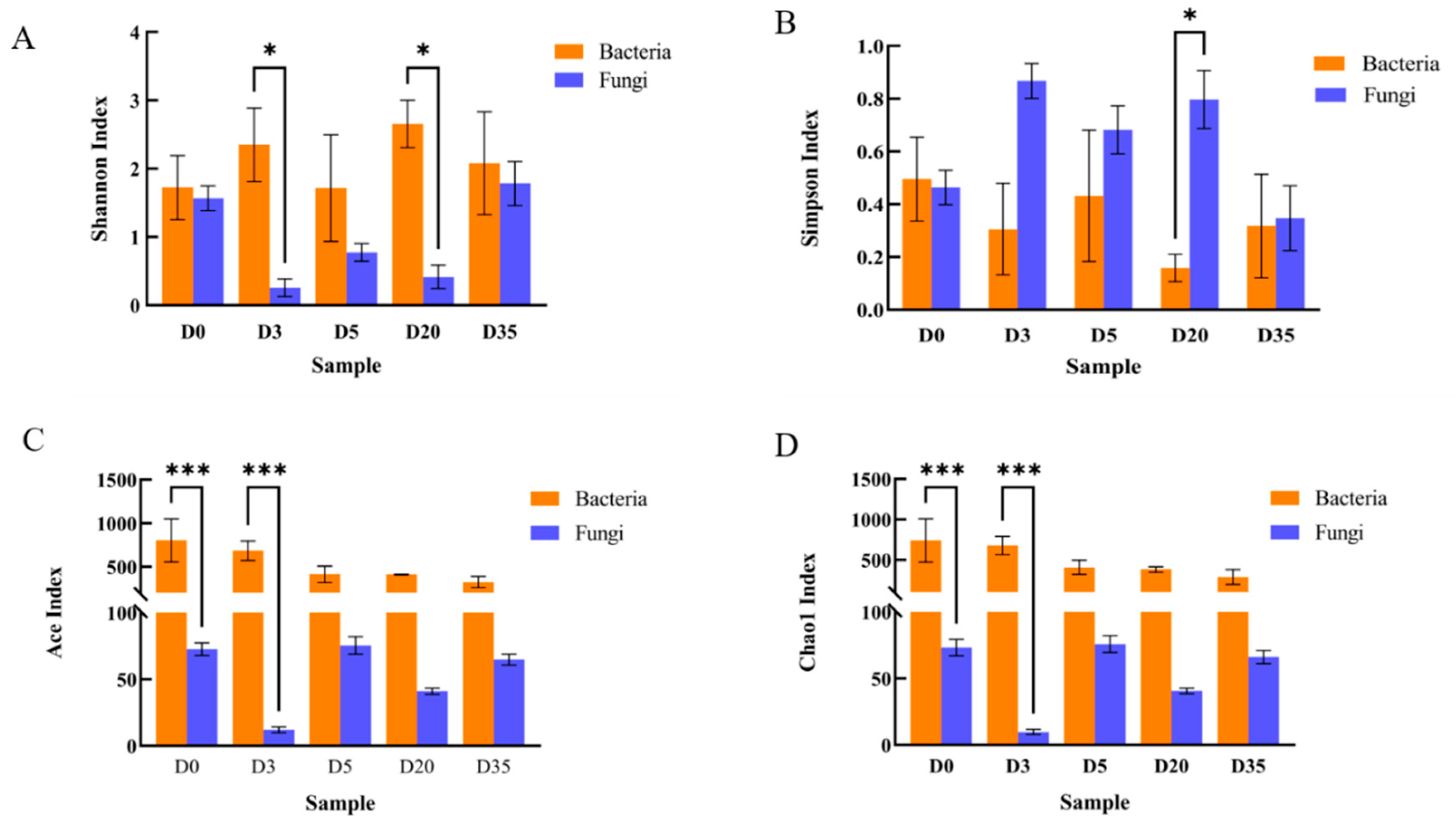
3.1. Yangfan Douchi Microbial Diversity during Different Stages of Fermentation
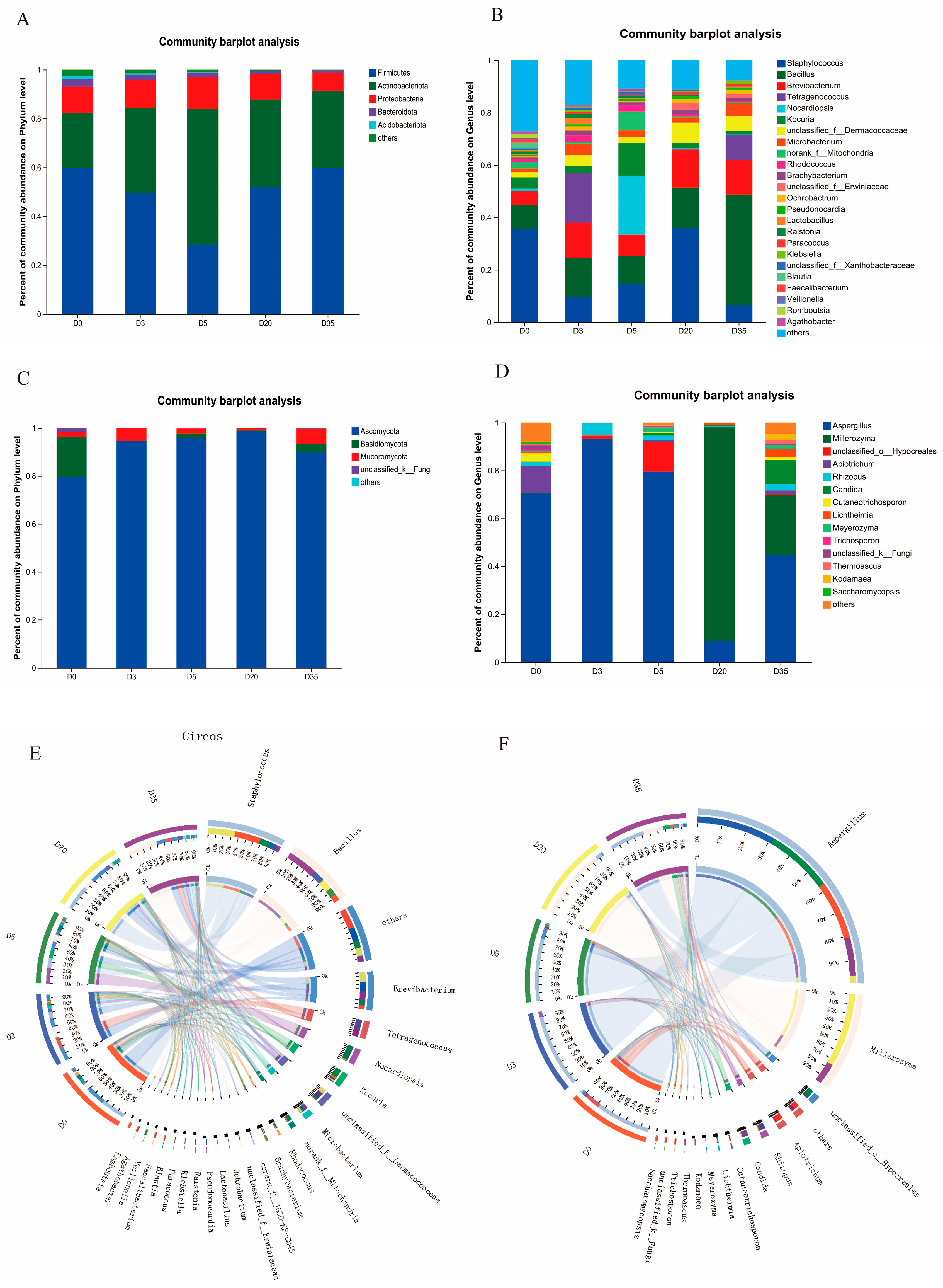
3.2. Changes in Bacterial Community Diversity and Composition in Aspergillus-Type Douchi
3.3. Changes in Fungal Community Diversity and Composition in Aspergillus-Type Douchi
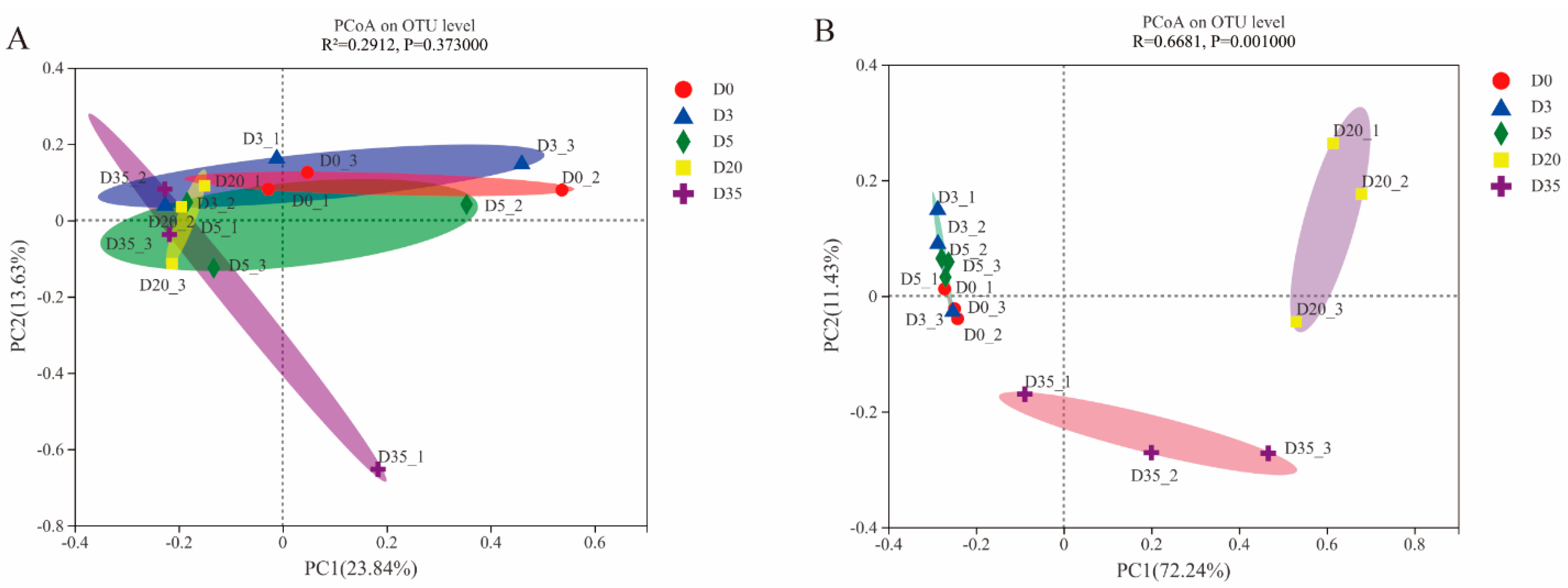
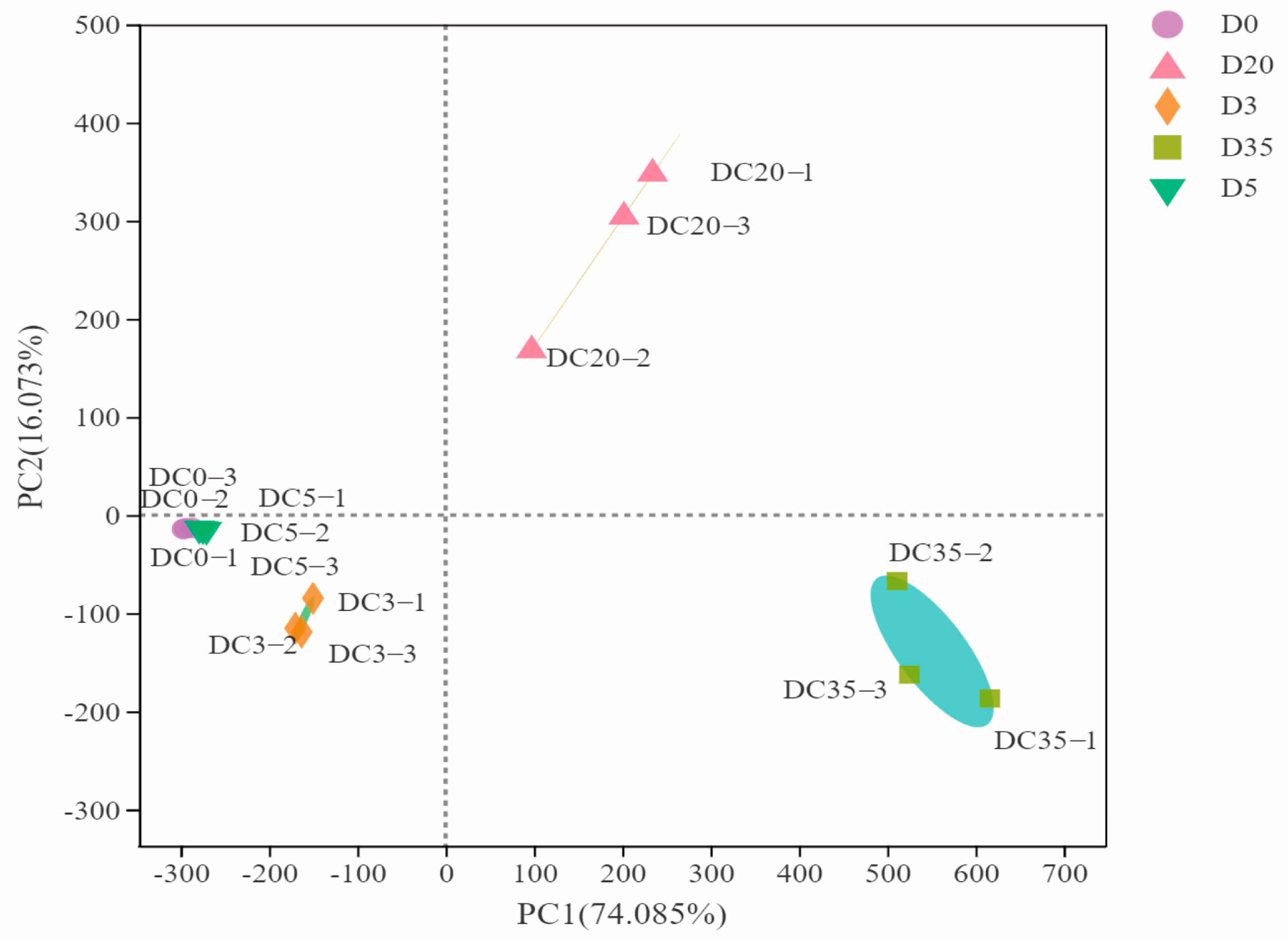
3.4. Beta Diversity of Aspergillus-Type Douchi during Fermentation Time
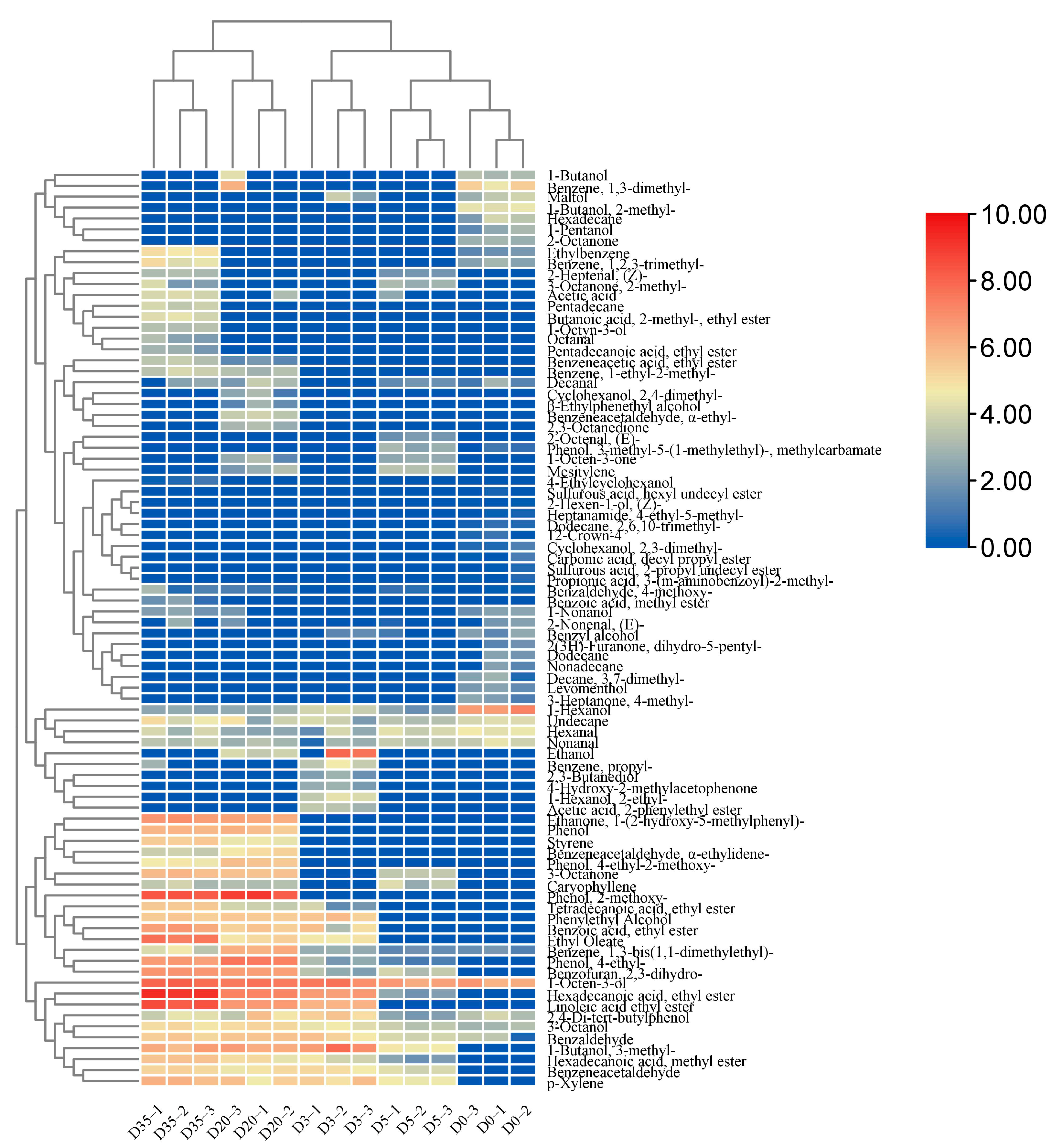
3.5. Dynamic Variations in Flavor Components during Douchi Fermentation
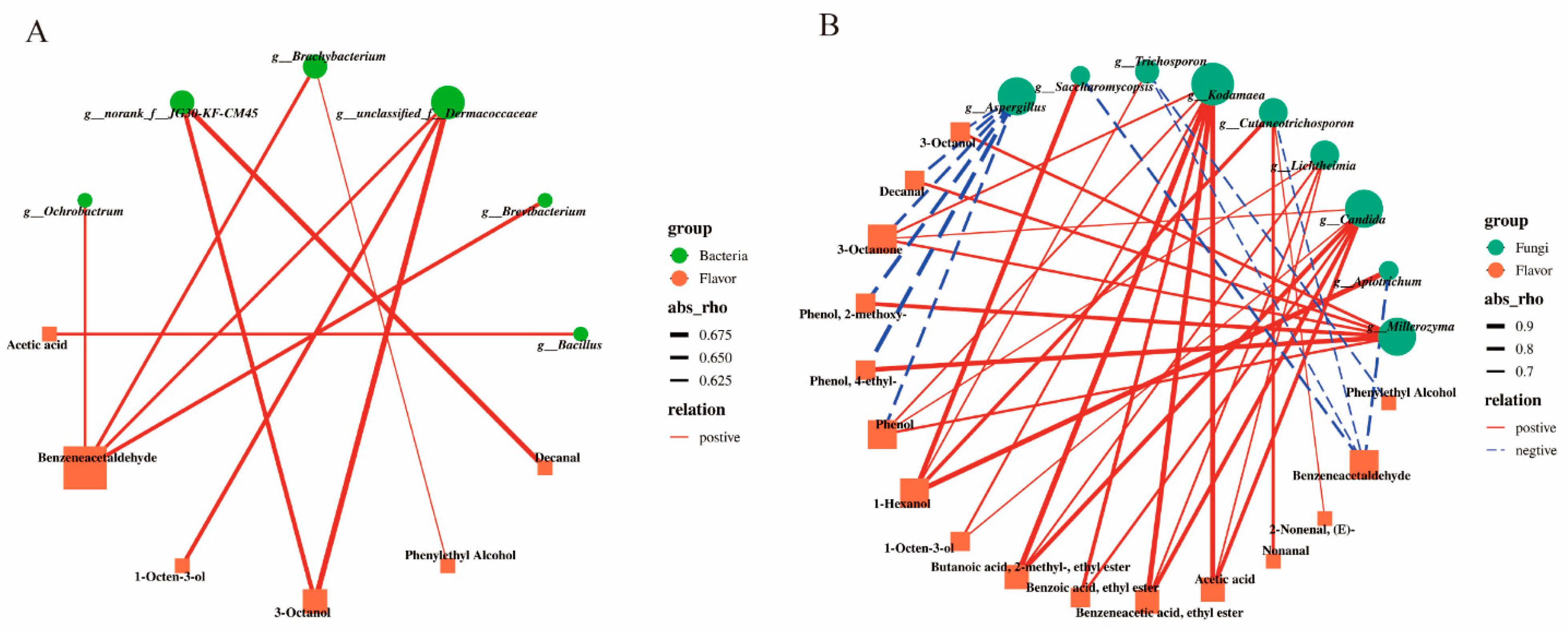
3.6. Analysis of the Correlation between the Core Microbial Communities and Volatile Flavor Compounds in Douchi
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, P.; Li, H.; Zhao, W.; Xiong, K.; Wen, H.; Yang, H. Dynamic analysis of physicochemical characteristics and microbial communities of aspergillus-type douchi during fermentation. Food Res. Int. 2022, 153, 110932. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Liu, X.W.; Huang, J.L.; Baloch, S.; Xu, X.; Pei, X.F. Microbial diversity and chemical analysis of shuidouchi, traditional Chinese fermented soybean. Food Res. Int. 2019, 116, 1289–1297. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yang, H.; Tu, Z.; Wang, X. High-throughput sequencing of microbial community diversity and dynamics during douchi fermentation. PLoS ONE 2016, 11, e0168166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Yang, L.; Zhang, J.; Li, H.; Tu, Z.; Wang, X. Exploring functional core bacteria in fermentation of a traditional Chinese food, Aspergillus-type douchi. PLoS ONE 2019, 14, e0226965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Li, P.; He, W.; Liao, L.; Xia, B.; Jiang, L.; Liu, Y. Analysis of microbial community and the characterization of Aspergillus flavus in Liuyang douchi during fermentation. LWT 2022, 154, 112567. [Google Scholar] [CrossRef]
- He, B.; Li, H.; Hu, Z.; Zhang, Y.; Sun, M.; Qiu, S.; Zeng, B. Difference in microbial community and taste compounds between Mucor-type and Aspergillus-type douchi during koji-Making. Food Res. Int. 2019, 121, 136–143. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, X.; Huang, Z.; Li, L.; Su, Y. Improving anti-alpha-glucosidase activity of douchi koji using a newly isolated strain of Bacillus subtilis B2. Int. J. Food Eng. 2011, 7, 1–18. [Google Scholar] [CrossRef]
- Xie, M.; Wu, J.; An, F.; Yue, X.; Tao, D.; Wu, R. An integrated metagenomic/metaproteomic investigation of microbiota in dajiang-meju, a traditional fermented soybean product in northeast China. Food Res. Int. 2019, 115, 414–424. [Google Scholar] [CrossRef]
- Jeong, D.W.; Jeong, K.; Lee, H.; Kim, C.T.; Lee, J.H. Effects of enterococcus faecium and staphylococcus succinus starters on the production of volatile compounds during doenjang fermentation. LWT 2019, 122, 108996. [Google Scholar] [CrossRef]
- Devanthi, P.; Gkatzionis, K. Soy sauce fermentation: Microorganisms, aroma formation, and process modification. Food Res. Int. 2019, 120, 364–374. [Google Scholar] [CrossRef]
- Dong, M.H.; Chun, B.H.; Kim, H.M.; Che, O.J. Characterization and correlation of microbial communities and metabolite and volatile compounds in doenjang fermentation. Food Res. Int. 2021, 148, 110645. [Google Scholar]
- Jing, C.; Cheng, Y.Q.; Yamaki, K.; Li, L.T. Anti-α-glucosidase activity of Chinese traditionally fermented soybean (douchi). Food Chem. 2007, 103, 1091–1096. [Google Scholar]
- Lu, X.; Bin, D.; Xu, B.J. A systematic, comparative study on the beneficial health components and antioxidant activities of commercially fermented soy products marketed in China. Food Chem. 2015, 174, 202–213. [Google Scholar]
- Peng, X.; Li, X.; Shi, X.; Guo, S. Evaluation of the aroma quality of Chinese traditional soy paste during storage based on principal component analysis. Food Chem. 2014, 151, 532–538. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Cai, W.; Wang, Y.; Ni, H.; Liu, Z.; Liu, J.; Zhong, J.; Hou, Q.; Shan, C.; Yang, X.; Guo, Z. Diversity of microbiota, microbial functions, and flavor in different types of low-temperature daqu. Food Res. Int. 2021, 150, 110734. [Google Scholar] [CrossRef]
- Liu, P.; Xiang, Q.; Sun, W.; Wang, X.; Lin, J.; Che, Z.; Ma, P. Correlation between microbial communities and key flavors during post-fermentation of Pixian broad bean paste. Food Res. Int. 2020, 137, 109513. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lu, Y.; Zhao, A.; Wu, Y.; Zhang, Y.; Yang, X. Quantitative Analyses for several nutrients and volatile components during fermentation of soybean by Bacillus subtilis natto. Food Chem. 2022, 374, 131725. [Google Scholar] [CrossRef]
- Yan, Z.Z.; Xiang, N.L.; Yan, Q.J.; Hong, J.H.; Hong, Z.Y.; Xiao, J.T.; Yun, G.L. Characterization and correlation of dominant bacteria and volatile compounds in post-fermentation process of Ba-bao Douchi. Food Res. Int. 2022, 160, 111688. [Google Scholar]
- Kim, T.; Lee, J.-H.; Park, M.-H.; Kim, H.-Y. Analysis of bacterial and fungal communities in Japanese-and Chinese-fermented soybean pastes using nested pcr-dgge. Curr. Microbiol. 2009, 60, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, W.; Li, L.; Chen, S.; Wu, Y.; Qi, B. Microbial community changes induced by a newly isolated salt-tolerant Tetragenococcus muriaticus improve the volatile flavor formation in low-salt fish sauce. Food Res. Int. 2022, 156, 111153. [Google Scholar] [CrossRef]
- Qi, Q.; Huang, J.; Zhou, R.; Jin, Y.; Wu, C. Abating biogenic amines and improving the flavor profile of cantonese soy sauce via co-culturing Tetragenococcus halophilus and Zygosaccharomyces rouxii. Food Microbiol. 2022, 106, 104056. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.; González-Guerra, A.; Vázquez, L.; Fernández-López, R.; Flórez, A.B.; de la Cruz, F.; Mayo, B. Isolation and phenotypic and genomic characterization of Tetragenococcus spp. from two spanish traditional blue-veined cheeses made of raw milk. Int. J. Food Microbiol. 2022, 371, 109670. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Jiang, S.; Xiong, S.; Wang, M.; Zhu, D.; Wei, H. Application of denaturing gradient gel electrophoresis to microbial diversity analysis in Chinese douchi. J. Sci. Food Agric. 2012, 92, 2171–2176. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, C.; Du, H.; Chen, C.; Xue, Q.; Hu, Y. Effect of starter cultures on dynamics succession of microbial communities, physicochemical parameters, enzyme activities, tastes and volatile flavor compounds during sufu fermentation. Food Chem. Adv. 2022, 1, 100057. [Google Scholar] [CrossRef]
- Henryk, J.; Małgorzata, M.; Alexandra, G.; Maciej, K. Determination of compounds responsible for tempeh aroma. Food Chem. 2007, 141, 459–465. [Google Scholar]
- Yao, D.; Xu, L.; Wu, M.; Wang, X.; Zhu, L.; Wang, C. Effects of microbial community succession on flavor compounds and physicochemical properties during cs sufu fermentation. LWT 2021, 152, 112313. [Google Scholar] [CrossRef]
- Mi, R.; Chen, X.; Xiong, S.; Qi, B.; Li, J.; Qiao, X.; Chen, W.; Qu, C.; Wang, S. Predominant yeasts in Chinese dong fermented pork (nanx wudl) and their aroma-producing properties in fermented sausage condition. Food Sci. Hum. Wellness. 2021, 10, 231–240. [Google Scholar] [CrossRef]
- Jiang, X.; Peng, D.; Zhang, W.; Duan, M.; Ruan, Z.; Huang, S.; Zhou, S.; Fang, Q. Effect of aroma-producing yeasts in high-salt liquid-state fermentation soy sauce and the biosynthesis pathways of the dominant Esters. Food Chem. 2021, 344, 128681. [Google Scholar] [CrossRef]
- Aryuman, P.; Lertsiri, S.; Visessanguan, W.; Niamsiri, N.; Bhumiratana, A.; Assavanig, A. Glutaminase-producing Meyerozyma (pichia) guilliermondii isolated from Thai soy sauce fermentation. Int. J. Food Microbiol. 2015, 192, 7–12. [Google Scholar] [CrossRef]
- Inoue, Y.; Kato, S.; Saikusa, M.; Suzuki, C.; Otsubo, Y.; Tanaka, Y.; Watanabe, H.; Hayase, F. Analysis of the cooked aroma and odorants that contribute to umami aftertaste of soy miso (Japanese soybean paste). Food Chem. 2016, 213, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Xu, E.; Long, J.; Wu, Z.; Li, H.; Wang, F.; Xu, X.; Jin, Z.; Jiao, A. Characterization of volatile flavor compounds in Chinese rice wine fermented from enzymatic extruded rice. J. Food Sci. 2015, 80, C1476–C1489. [Google Scholar] [CrossRef] [PubMed]
- Chung, H. Volatile flavor components in red fermented soybean (glycine max) curds. J. Agric. Food Chem. 2000, 48, 1803–1809. [Google Scholar] [CrossRef]
- Zang, J.; Xu, Y.; Xia, W.; Regenstein, J.M.; Yu, D.; Yang, F.; Jiang, Q. Correlations between microbiota succession and flavor formation during fermentation of Chinese low-salt fermented common carp (Cyprinus Carpio L.) inoculated with mixed starter cultures. Food Microbiol. 2020, 90, 103487. [Google Scholar] [CrossRef]
- Zhang, X.; Wei, J.; Zhao, S.; Jia, H.; Yuan, Y. Flavor differences between commercial and traditional soybean paste. LWT 2021, 142, 111052. [Google Scholar] [CrossRef]
- Yue, X.; Li, M.; Liu, Y.; Zhang, X.; Zheng, Y. Microbial diversity and function of soybean paste in east asia: What we know and what we don’t. Curr. Opin. Food Sci. 2021, 37, 145–152. [Google Scholar] [CrossRef]
- Ling, H.Z.; Shi, H.L.; Chen, X.C.; Cheng, K. Detection of the microbial diversity and flavour components of northeastern Chinese soybean paste during storage. Food Chem. 2022, 374, 131686. [Google Scholar] [CrossRef]
- Yao, D.; Ma, L.; Wu, M.; Wang, X.; Xu, L.; Yu, Q.; Wang, C. Effect of microbial communities on the quality characteristics of northeast soybean paste: Correlation between microorganisms and metabolites. LWT 2022, 164, 113648. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, M.; Xie, N.; Huang, M.; Feng, Y. Community structure of yeast in fermented soy sauce and screening of functional yeast with potential to enhance the soy sauce flavor. Int. J. Food Microbiol. 2022, 370, 109652. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, H.; Xu, J.; Wang, R.; Liu, X.; Peng, X.; Guo, S. Succession and Diversity of Microbial Flora during the Fermentation of Douchi and Their Effects on the Formation of Characteristic Aroma. Foods 2023, 12, 329. https://doi.org/10.3390/foods12020329
Zhao H, Xu J, Wang R, Liu X, Peng X, Guo S. Succession and Diversity of Microbial Flora during the Fermentation of Douchi and Their Effects on the Formation of Characteristic Aroma. Foods. 2023; 12(2):329. https://doi.org/10.3390/foods12020329
Chicago/Turabian StyleZhao, Huiyan, Jingting Xu, Ruican Wang, Xinran Liu, Xingyun Peng, and Shuntang Guo. 2023. "Succession and Diversity of Microbial Flora during the Fermentation of Douchi and Their Effects on the Formation of Characteristic Aroma" Foods 12, no. 2: 329. https://doi.org/10.3390/foods12020329
APA StyleZhao, H., Xu, J., Wang, R., Liu, X., Peng, X., & Guo, S. (2023). Succession and Diversity of Microbial Flora during the Fermentation of Douchi and Their Effects on the Formation of Characteristic Aroma. Foods, 12(2), 329. https://doi.org/10.3390/foods12020329