
Exploring the Interactions of Soybean 7S Globulin with Gallic Acid, Chlorogenic Acid and (−)-Epigallocatechin Gallate

Abstract
:
1. Introduction
2. Materials and Methods
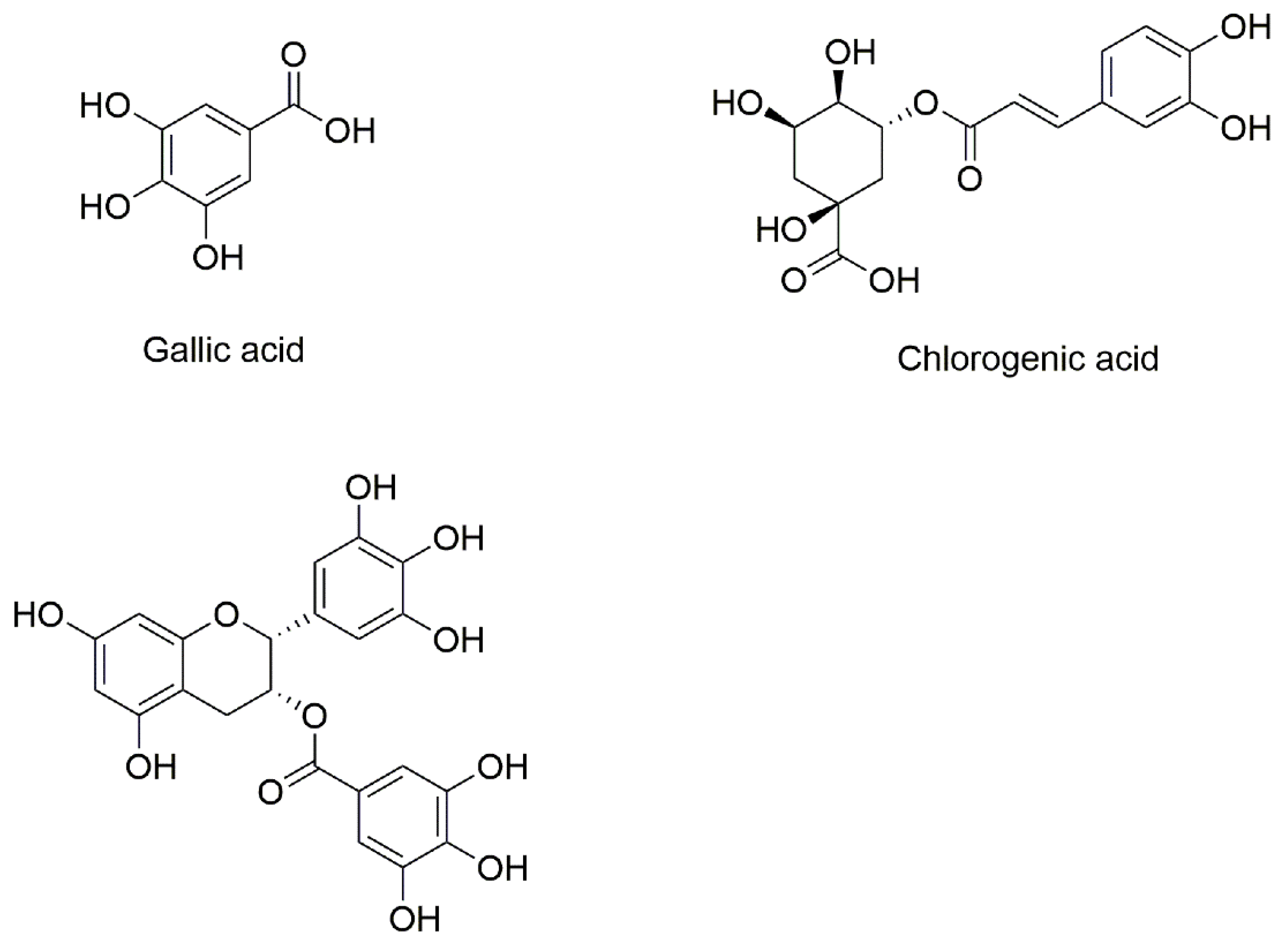
2.1. Materials
2.2. Preparation of 7S Globulin
2.3. Fluorescence Spectroscopy
2.4. CD Analysis
2.5. Surface Hydrophobicity
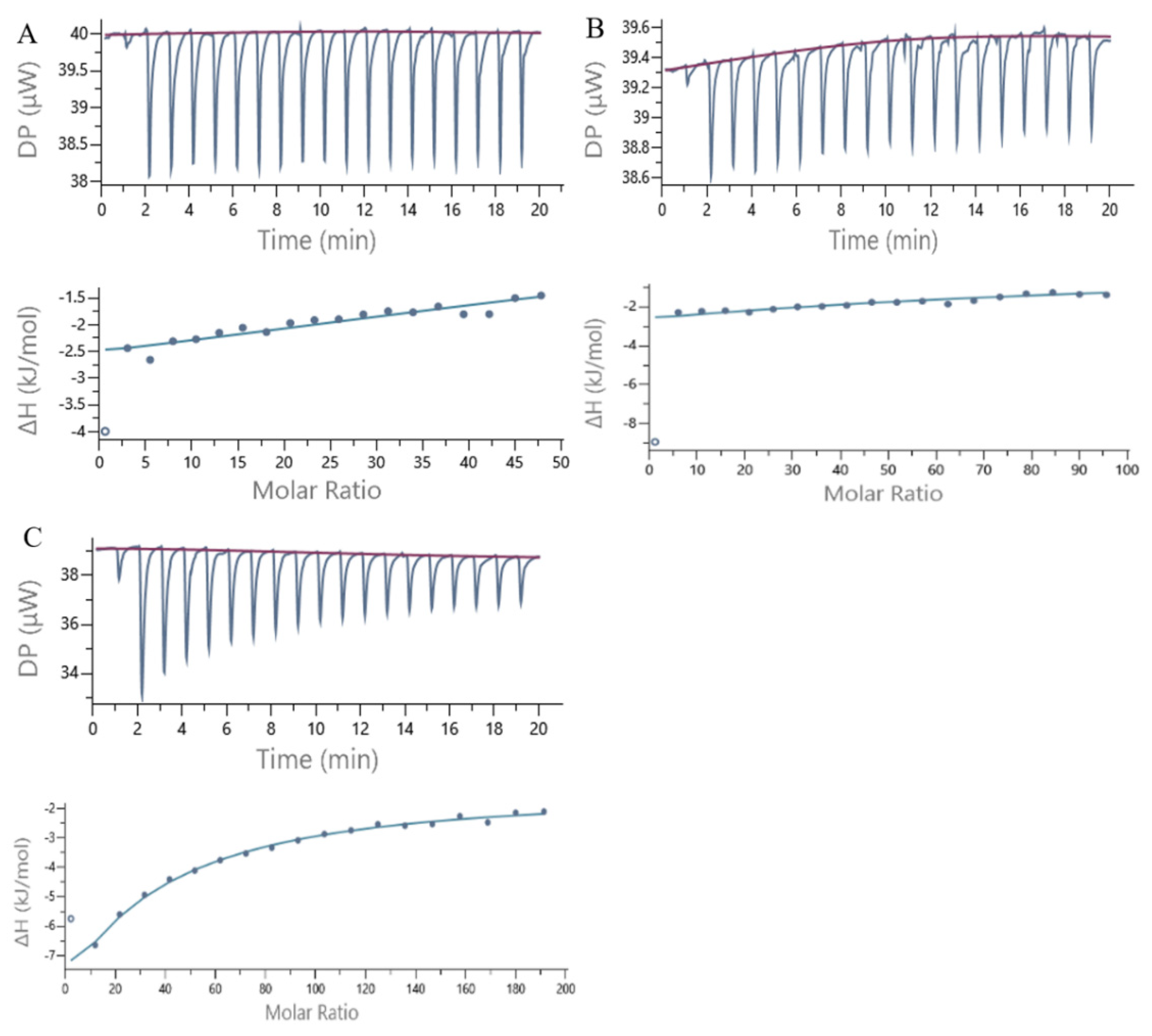
2.6. ITC Measurements
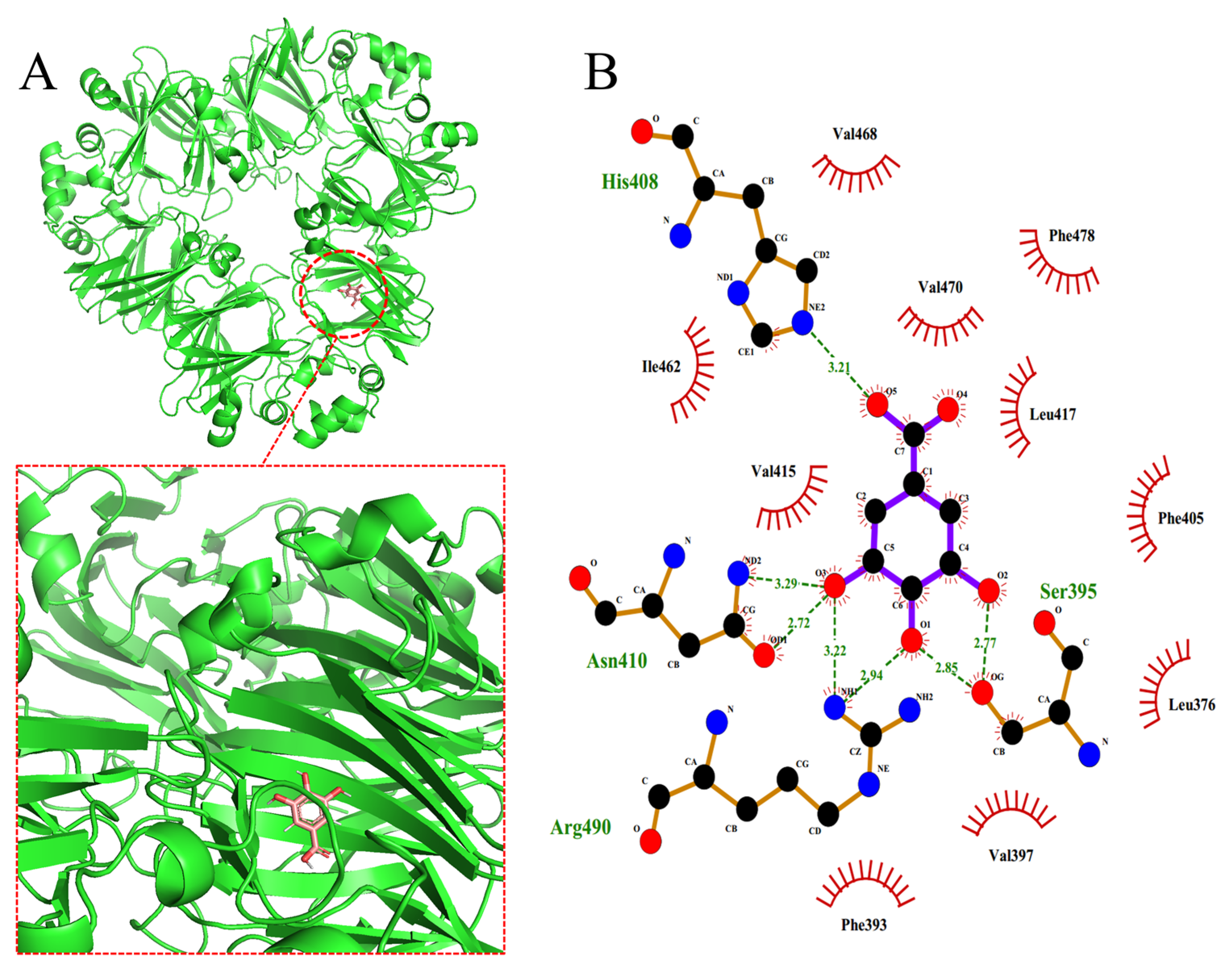
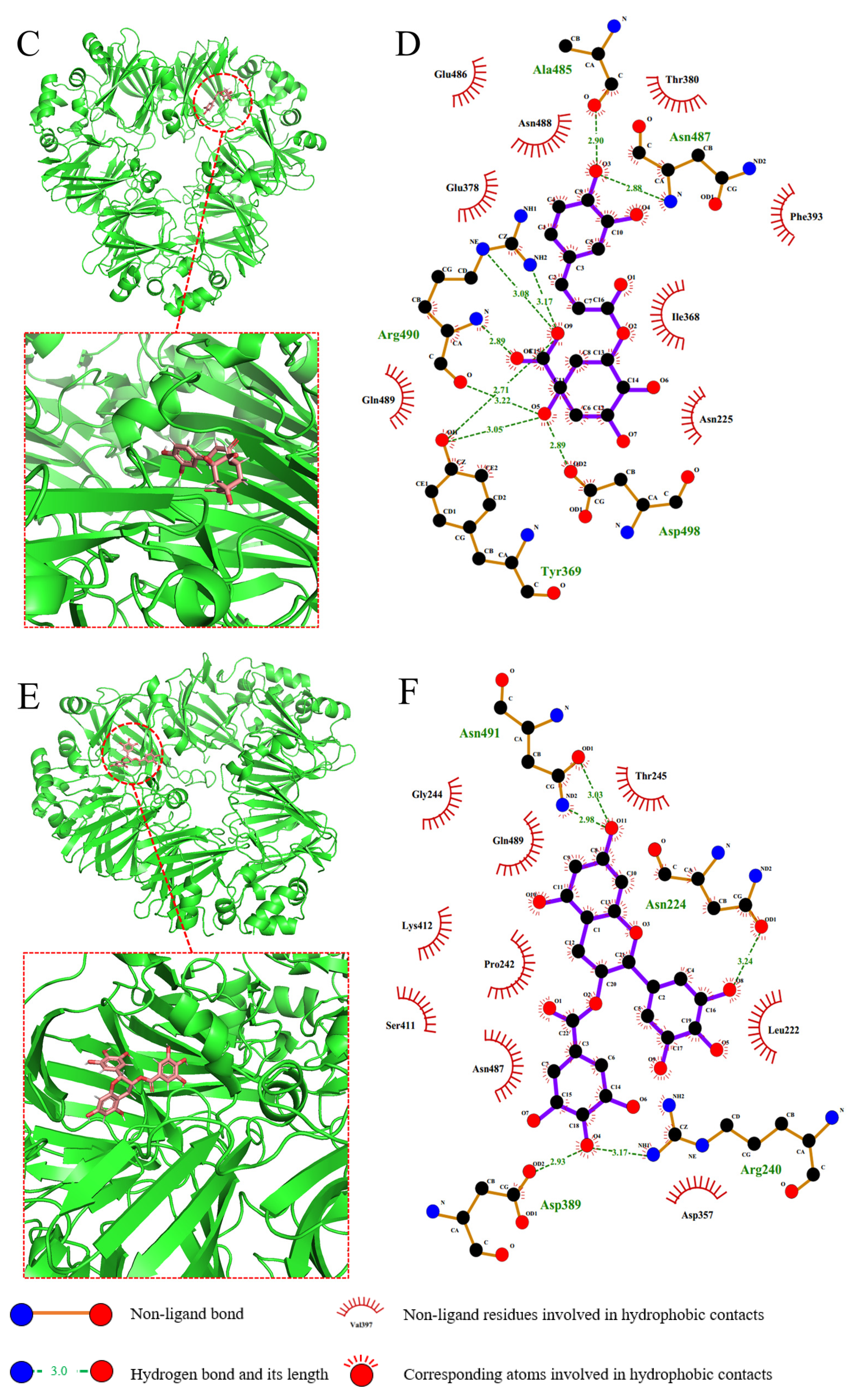
2.7. Molecular Docking
2.8. Statistical Analysis
3. Results and Discussion
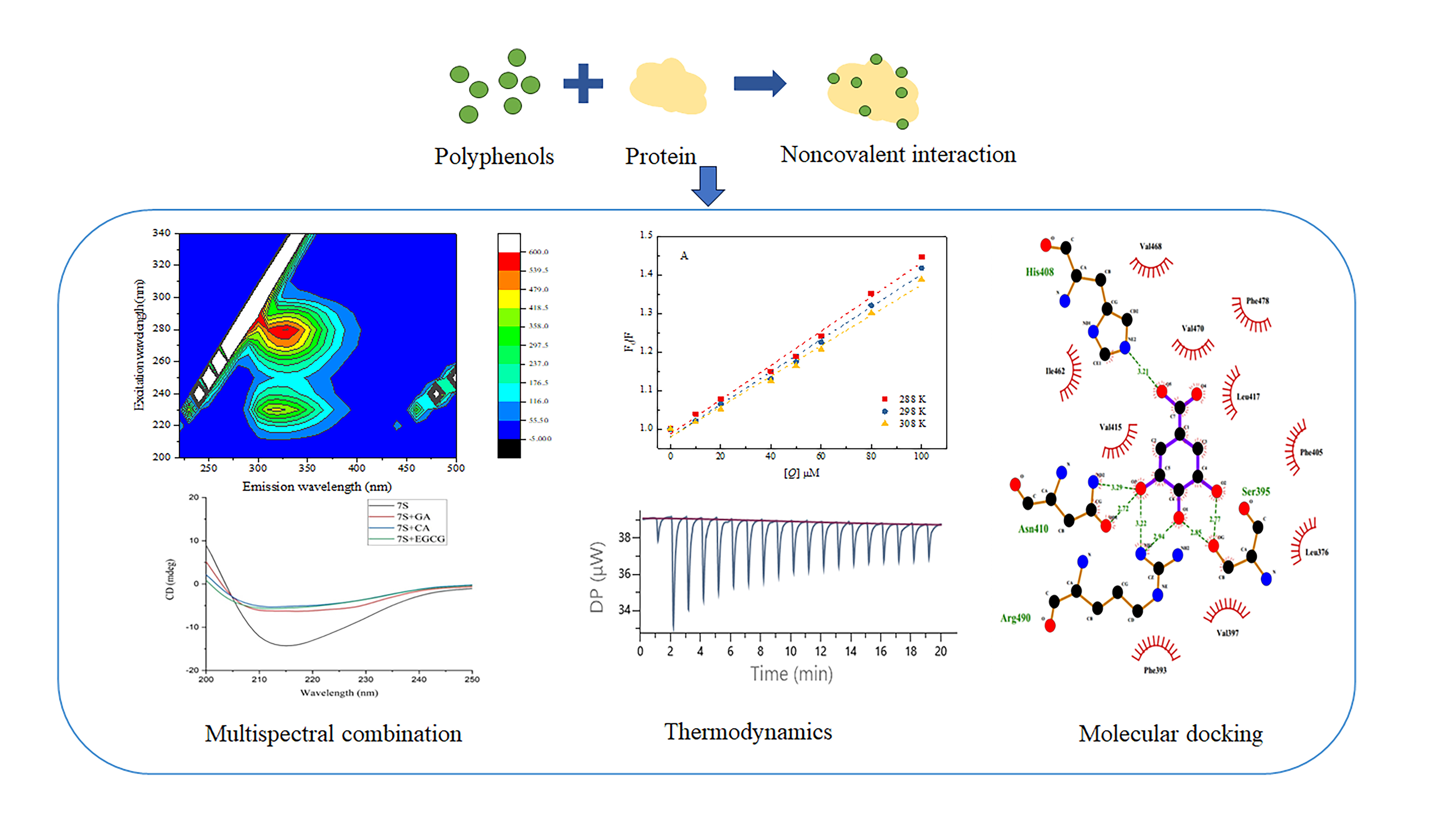
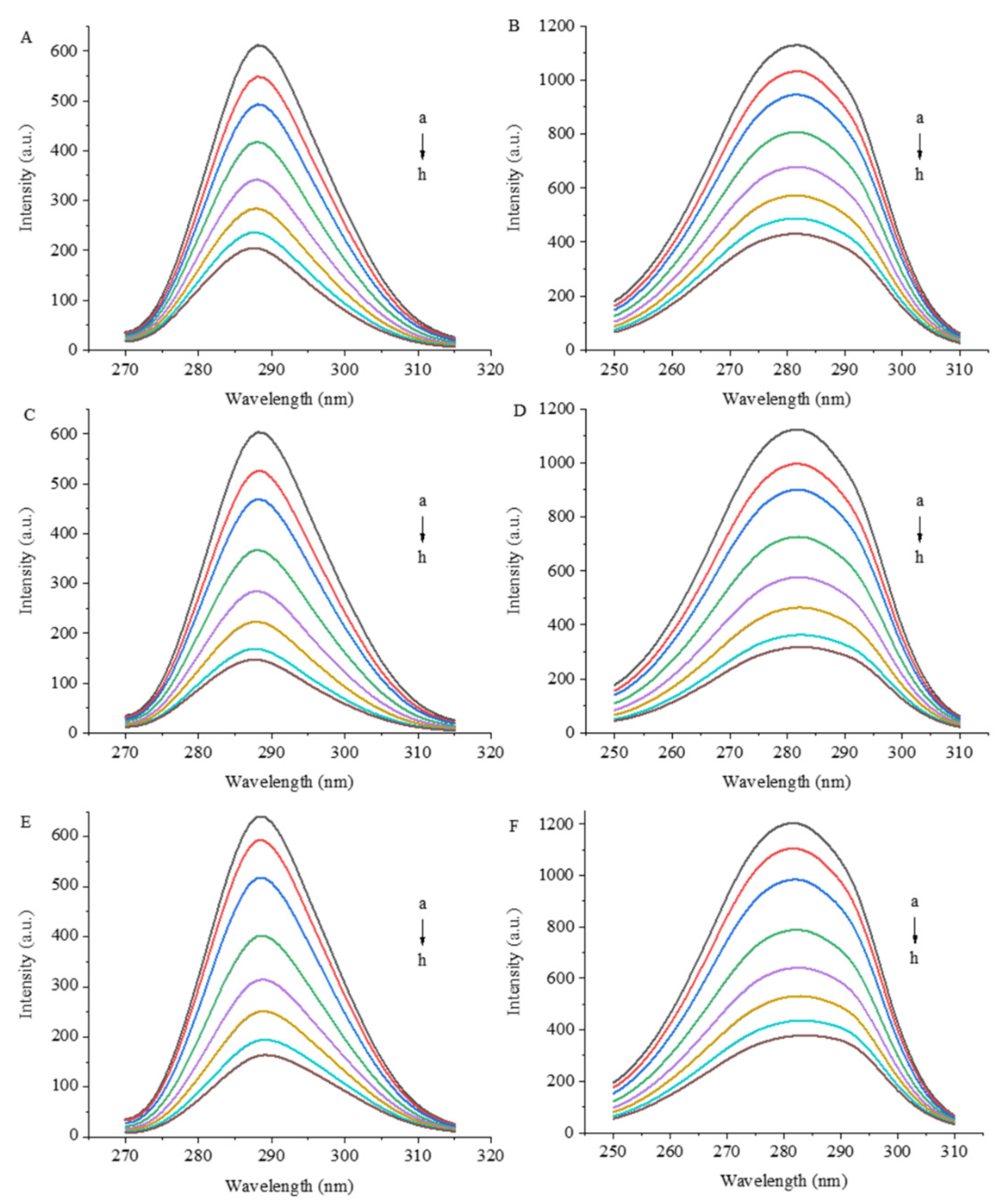
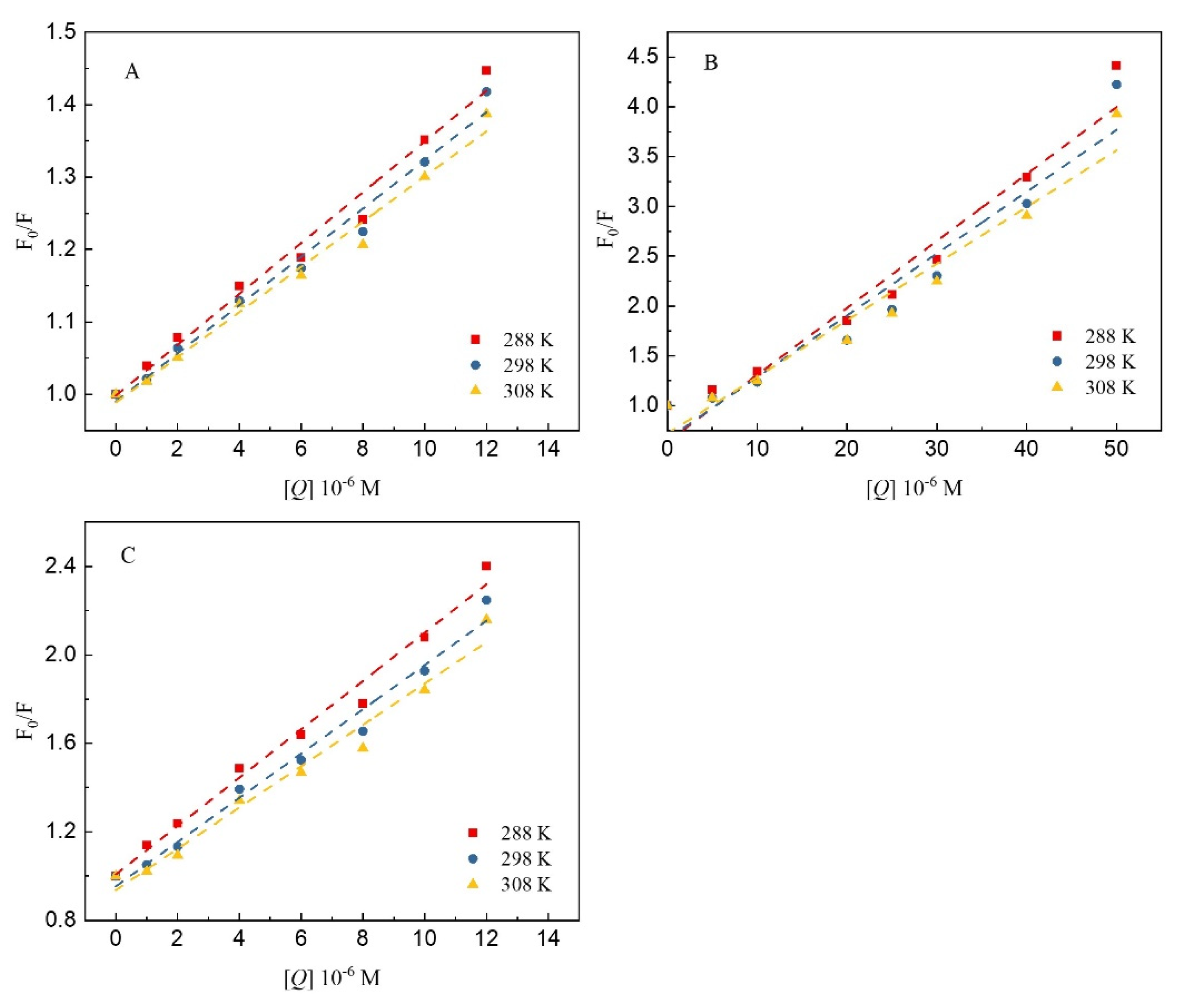
3.1. Fluorescence Quenching Assays
3.2. Synchronous Fluorescence Spectrometry
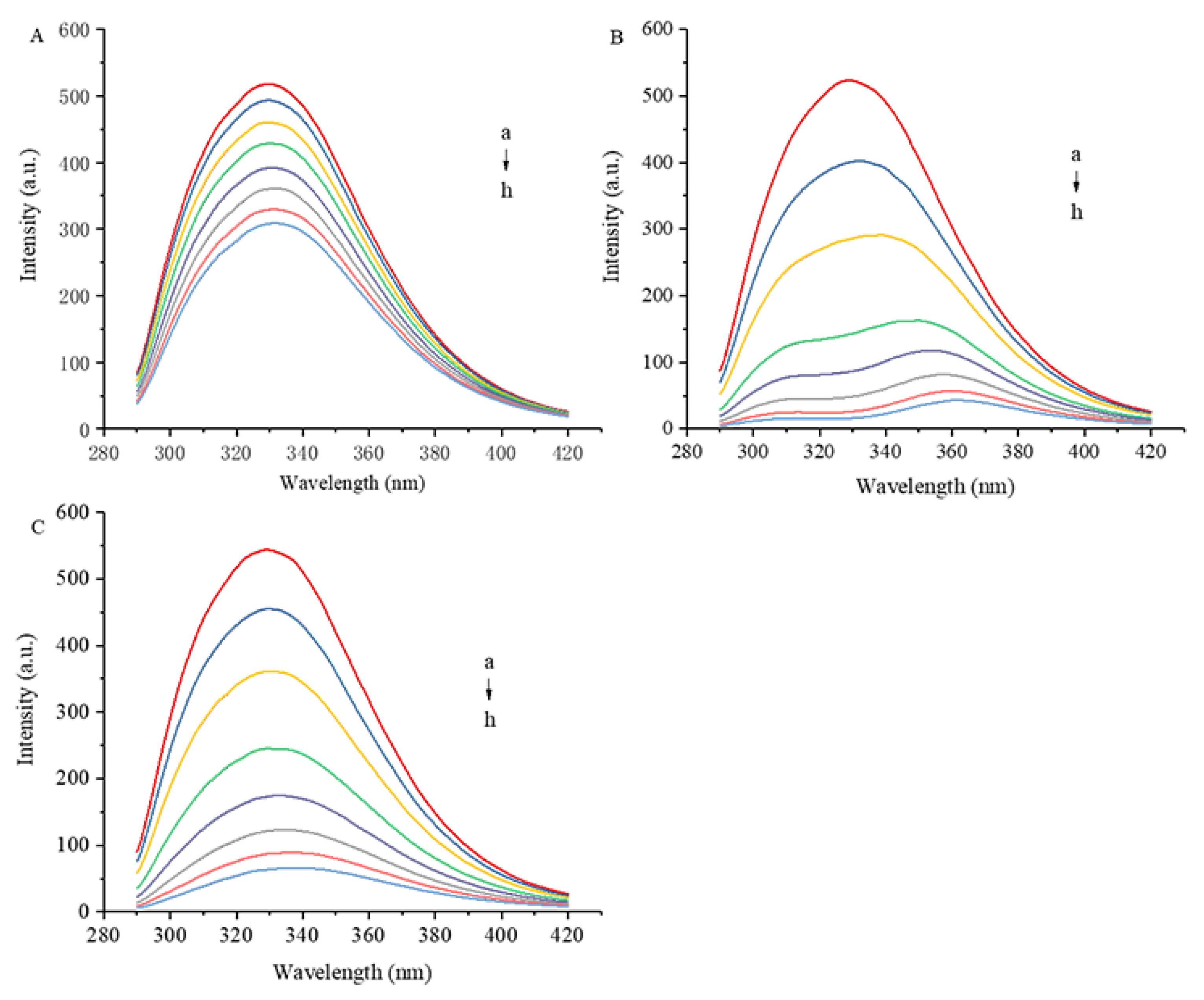
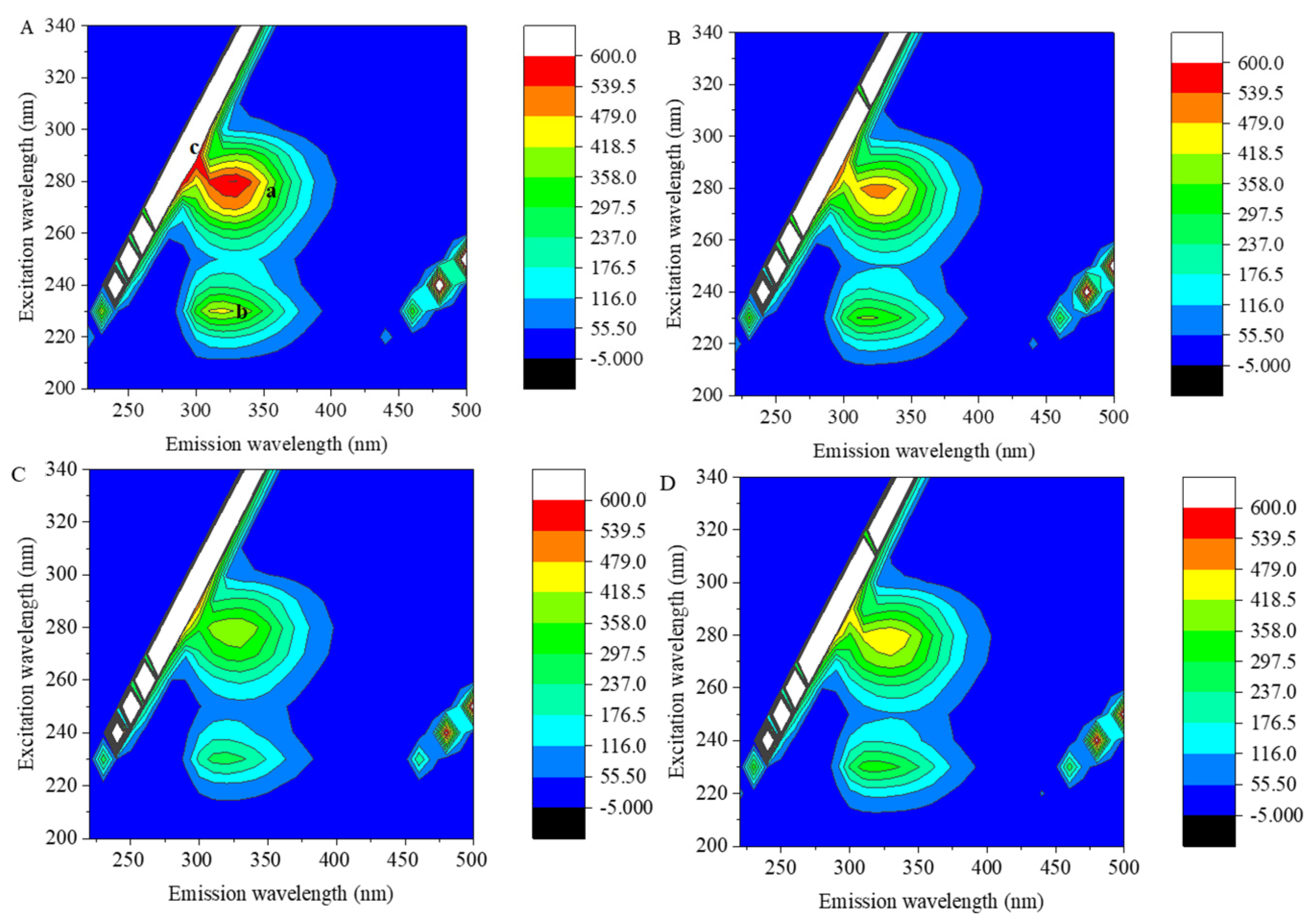
3.3. 3D Fluorescence Spectra
3.4. Mechanism of Fluorescence Quenching
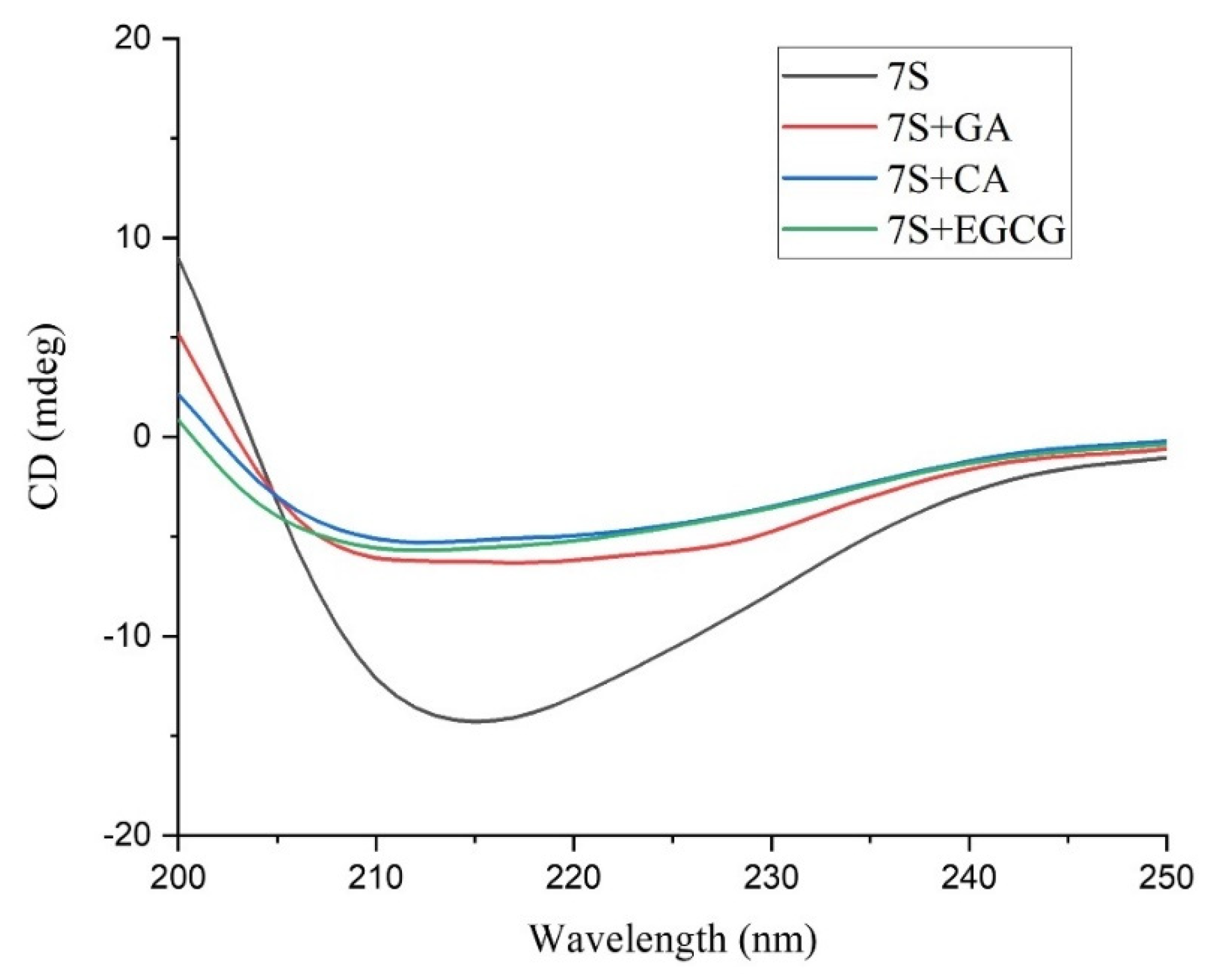
3.5. Changes in the Secondary and Tertiary Structure of 7S
3.6. Surface Hydrophobicity
3.7. ITC
3.8. Molecular Docking
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jia, Y.; Yan, X.; Huang, Y.; Zhu, H.; Qi, B.; Li, Y. Different interactions driving the binding of soy proteins (7S/11S) and flavonoids (quercetin/rutin): Alterations in the conformational and functional properties of soy proteins. Food Chem. 2022, 396, 133685. [Google Scholar] [PubMed]
- Liu, Y.; Yang, J.; Lei, L.; Wang, L.; Wang, X.; Ying Ma, K.; Yang, X.; Chen, Z.-Y. 7S protein is more effective than total soybean protein isolate in reducing plasma cholesterol. J. Funct. Foods 2017, 36, 18–26. [Google Scholar] [CrossRef]
- Han, K.; Feng, G.; Li, T.; Wan, Z.; Zhao, W.; Yang, X. Extension Region Domain of Soybean 7S Globulin Contributes to Serum Triglyceride-Lowering Effect via Modulation of Bile Acids Homeostasis. Mol. Nutr. Food Res. 2023, 67, 2200883. [Google Scholar]
- Tachibana, N.; Yamashita, Y.; Nagata, M.; Wanezaki, S.; Ashida, H.; Horio, F.; Kohno, M. Soy β-conglycinin improves glucose uptake in skeletal muscle and ameliorates hepatic insulin resistance in Goto-Kakizaki rats. Nutr. Res. 2014, 34, 160–167. [Google Scholar] [PubMed]
- Carbonaro, M.; Nucara, A. Legume Proteins and Peptides as Compounds in Nutraceuticals: A Structural Basis for Dietary Health Effects. Nutrients 2022, 14, 1188. [Google Scholar]
- Yan, S.; Xie, F.; Zhang, S.; Jiang, L.; Qi, B.; Li, Y. Effects of soybean protein isolate-polyphenol conjugate formation on the protein structure and emulsifying properties: Protein-polyphenol emulsification performance in the presence of chitosan. Colloids Surf. A Physicochem. Eng. Asp. 2021, 609, 125641. [Google Scholar]
- Wang, T.; Wang, N.; Yu, Y.; Yu, D.; Xu, S.; Wang, L. Study of soybean protein isolate-tannic acid non-covalent complexes by multi-spectroscopic analysis, molecular docking, and interfacial adsorption kinetics. Food Hydrocoll. 2023, 137, 108330. [Google Scholar]
- Dai, S.; Lian, Z.; Qi, W.; Chen, Y.; Tong, X.; Tian, T.; Lyu, B.; Wang, M.; Wang, H.; Jiang, L. Non-covalent interaction of soy protein isolate and catechin: Mechanism and effects on protein conformation. Food Chem. 2022, 384, 132507. [Google Scholar] [CrossRef]
- Ren, C.; Xiong, W.; Li, B. Binding interaction between β-conglycinin/glycinin and cyanidin-3-O-glucoside in acidic media assessed by multi-spectroscopic and thermodynamic techniques. Int. J. Biol. Macromol. 2019, 137, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, Y.; Shibata, M.; Yamada, S.; Nambu, Y.; Hirotsuka, M.; Matsumura, Y. Effects of heat treatment under low moisture conditions on the protein and oil in soybean seeds. Food Chem. 2019, 275, 577–584. [Google Scholar]
- Jia, Y.; Fu, Y.; Man, H.; Yan, X.; Huang, Y.; Sun, S.; Qi, B.; Li, Y. Comparative study of binding interactions between different dietary flavonoids and soybean β-conglycinin and glycinin: Impact on structure and function of the proteins. Food Res. Int. 2022, 161, 111784. [Google Scholar] [PubMed]
- Zhang, J.; Tian, Z.; Liang, L.; Subirade, M.; Chen, L. Binding interactions of β-conglycinin and glycinin with vitamin B12. J. Phys. Chem. B 2013, 117, 14018–14028. [Google Scholar] [CrossRef]
- Jin, B.; Zhou, X.; Liu, Y.; Li, X.; Mai, Y.; Liao, Y.; Liao, J. Physicochemical stability and antioxidant activity of soy protein/pectin/tea polyphenol ternary nanoparticles obtained by photocatalysis. Int. J. Biol. Macromol. 2018, 116, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Budryn, G.; Pałecz, B.; Rachwał-Rosiak, D.; Oracz, J.; Zaczyńska, D.; Belica, S.; Navarro-González, I.; Meseguer, J.M.V.; Pérez-Sánchez, H. Effect of inclusion of hydroxycinnamic and chlorogenic acids from green coffee bean in β-cyclodextrin on their interactions with whey, egg white and soy protein isolates. Food Chem. 2015, 168, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Mao, F.; Yang, F.; Zhao, Y.; Zhang, C.; Yamamoto, K. Interaction of dietary polyphenols with bovine milk proteins: Molecular structure-affinity relationship and influencing bioactivity aspects. Mol. Nutr. Food Res. 2011, 55, 1637–1645. [Google Scholar] [CrossRef] [PubMed]
- Abdelhameed, A.S.; Alam, P.; Khan, R.H. Binding of Janus kinase inhibitor tofacitinib with human serum albumin: Multi-technique approach. J. Biomol. Struct. Dyn. 2016, 34, 2037–2044. [Google Scholar] [CrossRef] [PubMed]
- Patra, D.; Mishra, A.K. Recent developments in multi-component synchronous fluorescence scan analysis. TrAC Trends Anal. Chem. 2002, 21, 787–798. [Google Scholar] [CrossRef]
- Kaur, J.; Katopo, L.; Hung, A.; Ashton, J.; Kasapis, S. Combined spectroscopic, molecular docking and quantum mechanics study of β-casein and p-coumaric acid interactions following thermal treatment. Food Chem. 2018, 252, 163–170. [Google Scholar] [CrossRef]
- Chen, N.; Gao, H.-X.; He, Q.; Yu, Z.-L.; Zeng, W.-C. Influence of structure complexity of phenolic compounds on their binding with maize starch. Food Struct. 2022, 33, 100286. [Google Scholar]
- Koshiyama, I. Comparison of acid-induced conformation changes between 7S and 11S globulin in soybean seeds. J. Sci. Food Agric. 1972, 23, 853–859. [Google Scholar] [CrossRef]
- Ghisaidoobe, A.B.; Chung, S.J. Intrinsic tryptophan fluorescence in the detection and analysis of proteins: A focus on Förster resonance energy transfer techniques. Int. J. Mol. Sci. 2014, 15, 22518–22538. [Google Scholar] [CrossRef]
- Miriani, M.; Keerati-u-rai, M.; Corredig, M.; Iametti, S.; Bonomi, F. Denaturation of soy proteins in solution and at the oil–water interface: A fluorescence study. Food Hydrocoll. 2011, 25, 620–626. [Google Scholar] [CrossRef]
- Sui, X.; Sun, H.; Qi, B.; Zhang, M.; Li, Y.; Jiang, L. Functional and conformational changes to soy proteins accompanying anthocyanins: Focus on covalent and non-covalent interactions. Food Chem. 2018, 245, 871–878. [Google Scholar] [CrossRef]
- Jia, Z.; Zheng, M.; Tao, F.; Chen, W.; Huang, G.; Jiang, J. Effect of covalent modification by (-)-epigallocatechin-3-gallate on physicochemical and functional properties of whey protein isolate. LWT Food Sci. Technol. 2016, 66, 305–310. [Google Scholar] [CrossRef]
- Dou, Z.; Chen, C.; Fu, X. The effect of ultrasound irradiation on the physicochemical properties and α-glucosidase inhibitory effect of blackberry fruit polysaccharide. Food Hydrocoll. 2019, 96, 568–576. [Google Scholar] [CrossRef]
- Jia, J.; Gao, X.; Hao, M.; Tang, L. Comparison of binding interaction between β-lactoglobulin and three common polyphenols using multi-spectroscopy and modeling methods. Food Chem. 2017, 228, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Al-Hanish, A.; Stanic-Vucinic, D.; Mihailovic, J.; Prodic, I.; Minic, S.; Stojadinovic, M.; Radibratovic, M.; Milcic, M.; Cirkovic Velickovic, T. Noncovalent interactions of bovine α-lactalbumin with green tea polyphenol, epigalocatechin-3-gallate. Food Hydrocoll. 2016, 61, 241–250. [Google Scholar] [CrossRef]
- Hasni, I.; Bourassa, P.; Hamdani, S.; Samson, G.; Carpentier, R.; Tajmir-Riahi, H.-A. Interaction of milk α- and β-caseins with tea polyphenols. Food Chem. 2011, 126, 630–639. [Google Scholar] [CrossRef]
- Liu, F.; Ma, C.; McClements, D.J.; Gao, Y. A comparative study of covalent and non-covalent interactions between zein and polyphenols in ethanol-water solution. Food Hydrocoll. 2017, 63, 625–634. [Google Scholar] [CrossRef]
- Li, Y.; Lim, L.T.; Kakuda, Y. Electrospun zein fibers as carriers to stabilize (−)-epigallocatechin gallate. J. Food Sci. 2009, 74, C233–C240. [Google Scholar] [CrossRef] [PubMed]
- Kaspchak, E.; Misugi Kayukawa, C.T.; Meira Silveira, J.L.; Igarashi-Mafra, L.; Mafra, M.R. Interaction of Quillaja bark saponin and bovine serum albumin: Effect on secondary and tertiary structure, gelation and in vitro digestibility of the protein. LWT 2020, 121, 108970. [Google Scholar] [CrossRef]
- Jiang, J.; Zhang, Z.; Zhao, J.; Liu, Y. The effect of non-covalent interaction of chlorogenic acid with whey protein and casein on physicochemical and radical-scavenging activity of in vitro protein digests. Food Chem. 2018, 268, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Xiong, W.; Li, J.; Li, B. Comparison of binding interactions of cyanidin-3-O-glucoside to β-conglycinin and glycinin using multi-spectroscopic and thermodynamic methods. Food Hydrocoll. 2019, 92, 155–162. [Google Scholar] [CrossRef]
- Barratt, E.; Bingham, R.J.; Warner, D.J.; Laughton, C.A.; Phillips, S.E.; Homans, S.W. Van der Waals interactions dominate ligand-protein association in a protein binding site occluded from solvent water. J. Am. Chem. Soc. 2005, 127, 11827–11834. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhong, Q. Binding between bixin and whey protein at pH 7.4 studied by spectroscopy and isothermal titration calorimetry. J. Agric. Food Chem. 2012, 60, 1880–1886. [Google Scholar] [CrossRef]
- Frazier, R.A.; Deaville, E.R.; Green, R.J.; Stringano, E.; Willoughby, I.; Plant, J.; Mueller-Harvey, I. Interactions of tea tannins and condensed tannins with proteins. J. Pharm. Biomed. Anal. 2010, 51, 490–495. [Google Scholar] [CrossRef]
- Nagy, K.; Courtet-Compondu, M.C.; Williamson, G.; Rezzi, S.; Kussmann, M.; Rytz, A. Non-covalent binding of proteins to polyphenols correlates with their amino acid sequence. Food Chem. 2012, 132, 1333–1339. [Google Scholar] [CrossRef]
- Lelis, C.A.; Hudson, E.A.; Ferreira, G.M.D.; Ferreira, G.M.D.; da Silva, L.H.M.; da Silva, M.D.C.H.; Pinto, M.S.; Dos Santos Pires, A.C. Binding thermodynamics of synthetic dye Allura Red with bovine serum albumin. Food Chem. 2017, 217, 52–58. [Google Scholar] [CrossRef]
- Singh, A.; Meena, M.; Kumar, D.; Dubey, A.K.; Hassan, M.I. Structural and functional analysis of various globulin proteins from soy seed. Crit. Rev. Food Sci. Nutr. 2015, 55, 1491–1502. [Google Scholar] [CrossRef]
- Chanphai, P.; Tajmir-Riahi, H.A. Tea polyphenols bind serum albumins: A potential application for polyphenol delivery. Food Hydrocoll. 2019, 89, 461–467. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | T (K) | KSV (104 L/mol) | n | Ka (104 L/mol) |
|---|---|---|---|---|
| 7S−GA | 308 | 0.40 ± 0.01 a | 0.94 ± 0.04 a | 4.21 ± 0.22 a |
| 298 | 0.42 ± 0.02 ab | 1.12 ± 0.06 b | 5.10 ± 0.30 b | |
| 288 | 0.44 ± 0.02 b | 1.18 ± 0.06 b | 5.37 ± 0.30 b | |
| 7S−CA | 308 | 5.69 ± 0.03 a | 1.17 ± 0.06 a | 6.23 ± 0.32 a |
| 298 | 6.21 ± 0.17 b | 1.42 ± 0.05 b | 7.44 ± 0.32 b | |
| 288 | 6.72 ± 0.13 c | 1.36 ± 0.06 b | 7.09 ± 0.31 b | |
| 7S−EGCG | 308 | 2.35 ± 0.03 a | 0.90 ± 0.04 a | 4.54 ± 0.20 a |
| 298 | 2.53 ± 0.01 b | 1.25 ± 0.06 b | 6.23 ± 0.30 b | |
| 288 | 2.77 ± 0.01 c | 1.52 ± 0.11 c | 7.57 ± 0.56 c |
| Sample | Secondary Structure Content (%) | Surface Hydrophobicity | |||
|---|---|---|---|---|---|
| α-Helixes | β-Sheets | β-Turns | Random Coils | S0 | |
| 7S | 22.6 ± 0.2 c | 36.2 ± 0.6 b | 16.3 ± 0.5 c | 24.9 ± 0.2 a | 56.80 ± 1.23 d |
| 7S−GA | 20.7 ± 0.1 b | 37.2 ± 0.4 b | 12.7 ± 0.0 a | 29.4 ± 0.6 b | 45.57 ± 0.65 c |
| 7S−CA | 17.9 ± 0.3 a | 33.3 ± 0.2 a | 18.3 ± 0.4 d | 30.5 ± 0.1 b | 41.59 ± 0.81 b |
| 7S−EGCG | 18.5 ± 0.2 ab | 34.7 ± 0.1 a | 14.0 ± 0.2 b | 32.8 ± 0.1 c | 25.97 ± 0.53 a |
| Compounds | N (Sites) | KD (M) | ΔH (kJ/mol) | ΔG (kJ/mol) | TΔS (kJ/mol) |
|---|---|---|---|---|---|
| 7S−GA | 10.0 ± 0.40 c | 7.20 × 10−4 ± 2.91 × 10−5 b | −3.66 ± 0.22 c | −18.0 | −14.3 |
| 7S−CA | 7.23 ± 0.75 b | 4.44 × 10−4 ± 1.75 × 10−5 a | −162 ± 4.62 b | −13.4 | 148 |
| 7S−EGCG | 1.68 ± 0.38 a | 4.85 × 10−4 ± 1.35 × 10−5 a | −335 ± 17.67 a | −18.9 | 316 |
| Polyphenol | Donor Atom | Acceptor Atom | Hydrogen Bond Length (Å) |
|---|---|---|---|
| GA | NE2 (His-408) | O5 (GA) | 3.21 |
| O3 (GA) | OD1 (Asn-410) | 2.72 | |
| ND2 (Asn-410) | O3 (GA) | 3.29 | |
| NH1 (Arg-490) | O3 (GA) | 2.94 | |
| NH1 (Arg-490) | O1 (GA) | 2.94 | |
| OG (Ser-395) | O1 (GA) | 2.85 | |
| O2 (GA) | OG (Ser-395) | 2.77 | |
| CA | O3 (CA) | O (Ala-485) | 2.90 |
| N (Asn-487) | O3 (CA) | 2.88 | |
| NH2 (Arg-490) | O9 (CA) | 3.17 | |
| OH (Tyr-369) | O9 (CA) | 2.71 | |
| N (Arg-490) | O8 (CA) | 2.89 | |
| O5 (CA) | O (Arg-490) | 3.22 | |
| O5 (CA) | OD2 (Asp-498) | 2.89 | |
| EGCG | O11 (EGCG) | OD1 (Asn-491) | 3.03 |
| ND2 (Asn-491) | O11 (EGCG) | 2.98 | |
| O8 (EGCG) | OD1 (Asn-224) | 3.24 | |
| O4 (EGCG) | OD2 (Asp-389) | 2.93 | |
| NH1 (Arg-240) | O4 (EGCG) | 3.17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, S.; Meng, L.; Lin, Y.; Dong, X.; Dong, M. Exploring the Interactions of Soybean 7S Globulin with Gallic Acid, Chlorogenic Acid and (−)-Epigallocatechin Gallate. Foods 2023, 12, 4013. https://doi.org/10.3390/foods12214013
Zhou S, Meng L, Lin Y, Dong X, Dong M. Exploring the Interactions of Soybean 7S Globulin with Gallic Acid, Chlorogenic Acid and (−)-Epigallocatechin Gallate. Foods. 2023; 12(21):4013. https://doi.org/10.3390/foods12214013
Chicago/Turabian StyleZhou, Siduo, Ling Meng, Yanfei Lin, Xueqian Dong, and Mingsheng Dong. 2023. "Exploring the Interactions of Soybean 7S Globulin with Gallic Acid, Chlorogenic Acid and (−)-Epigallocatechin Gallate" Foods 12, no. 21: 4013. https://doi.org/10.3390/foods12214013
APA StyleZhou, S., Meng, L., Lin, Y., Dong, X., & Dong, M. (2023). Exploring the Interactions of Soybean 7S Globulin with Gallic Acid, Chlorogenic Acid and (−)-Epigallocatechin Gallate. Foods, 12(21), 4013. https://doi.org/10.3390/foods12214013