
Effects of Vine Water Status on Malate Metabolism and γ-Aminobutyric Acid (GABA) Pathway-Related Amino Acids in Marselan (Vitis vinifera L.) Grape Berries

Abstract
:1. Introduction
2. Materials and Methods
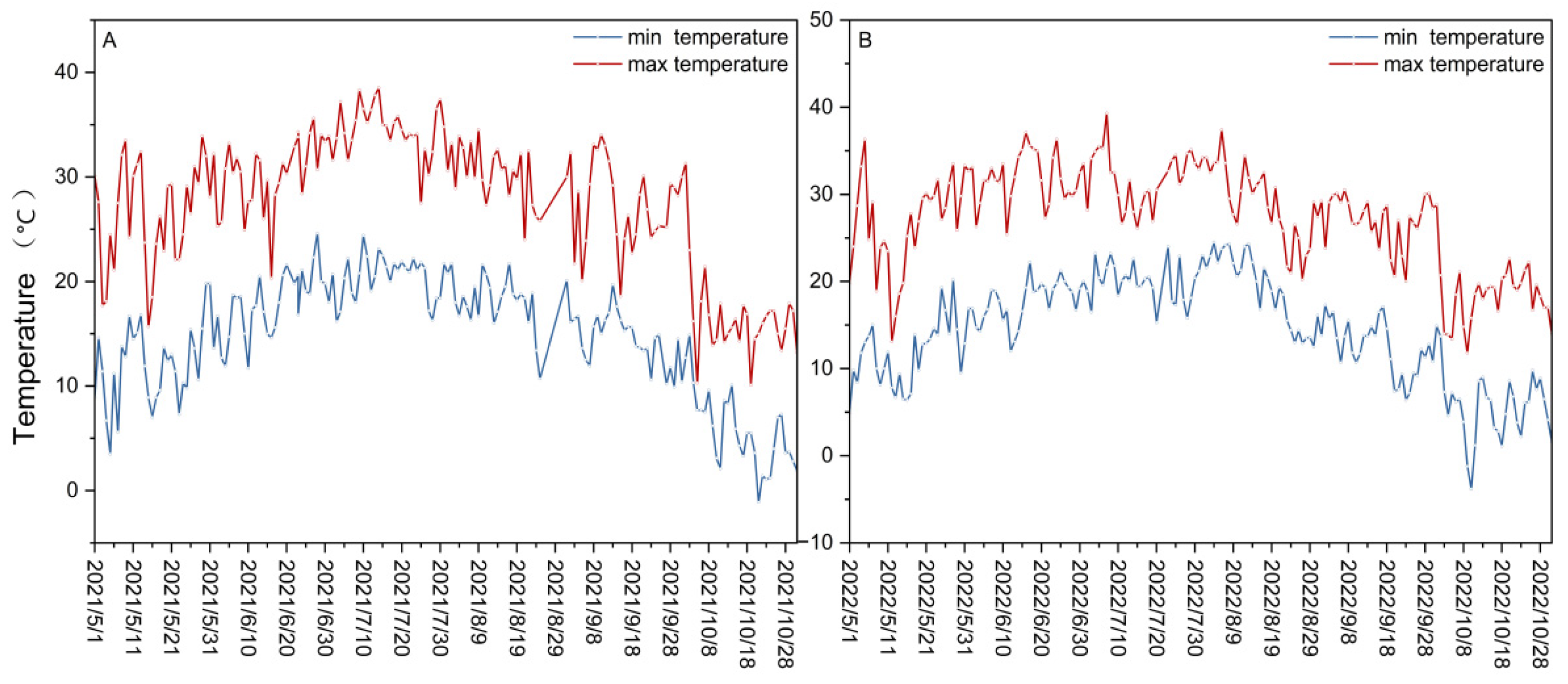
2.1. Climatic Conditions and Experimental Design
2.2. Water Status and Phytochemicals of the Grape Berries
2.3. Organic Acids, Enzyme Extraction, and Activity Assay
2.4. Analysis of Soluble Sugars and Amino Acids Using HPLC
2.5. qRT-PCR Analysis
2.6. Statistical Analysis
3. Results
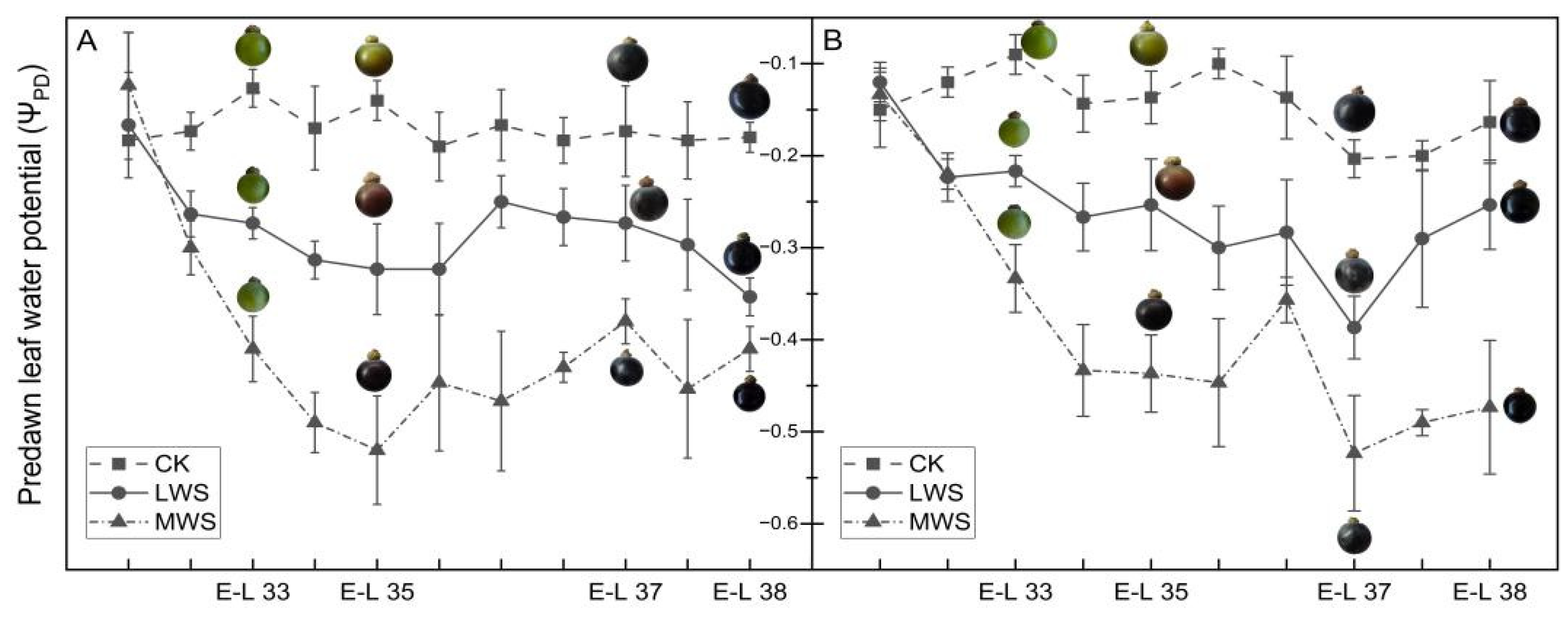
3.1. The ΨPD of ‘Marselan’ under Water Stress
3.2. Phenotype and Phytochemicals of the Grape Berries under Water Stress
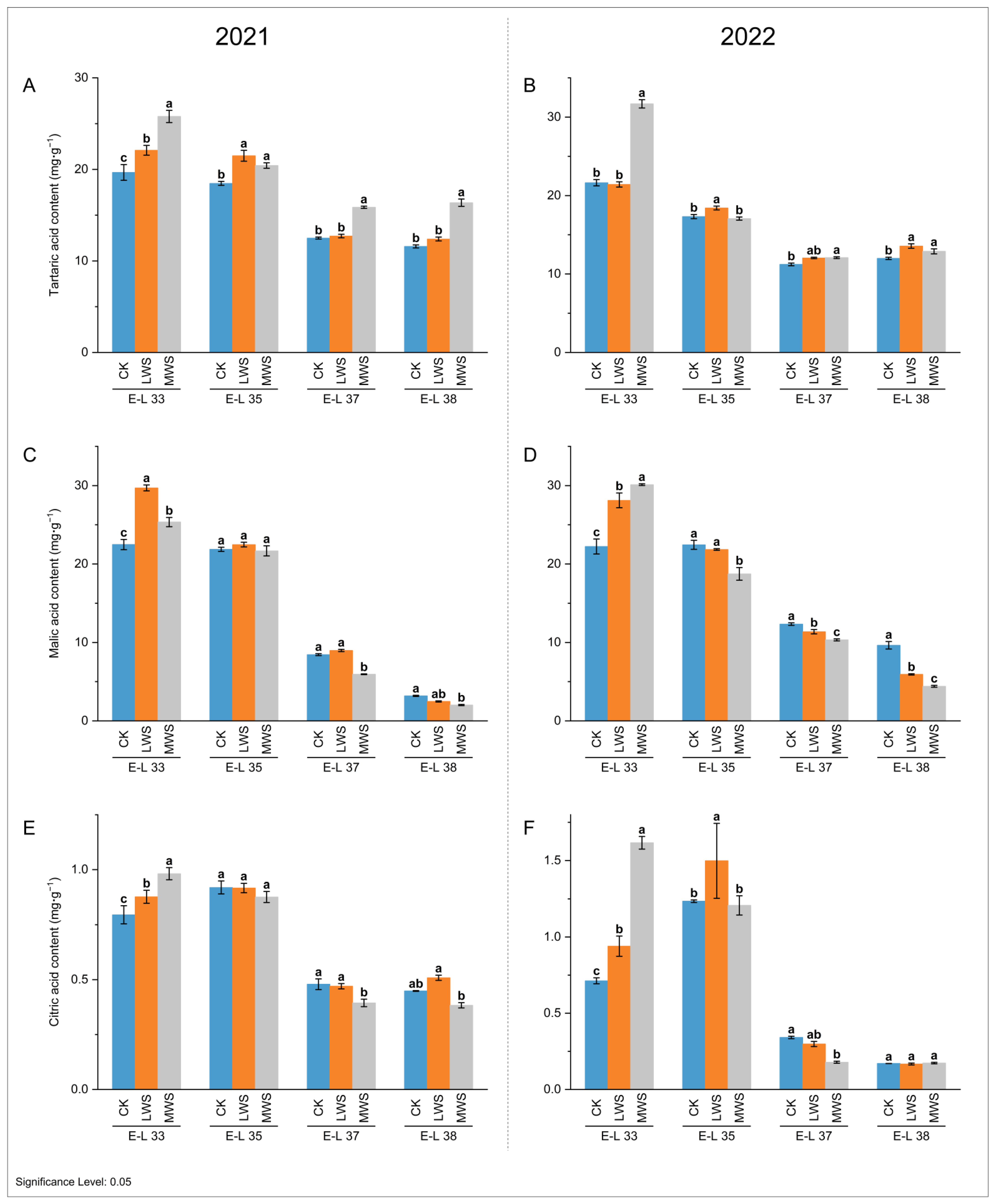
3.3. Analysis of the Main Organic Acids in Grape Berries under Water Stress
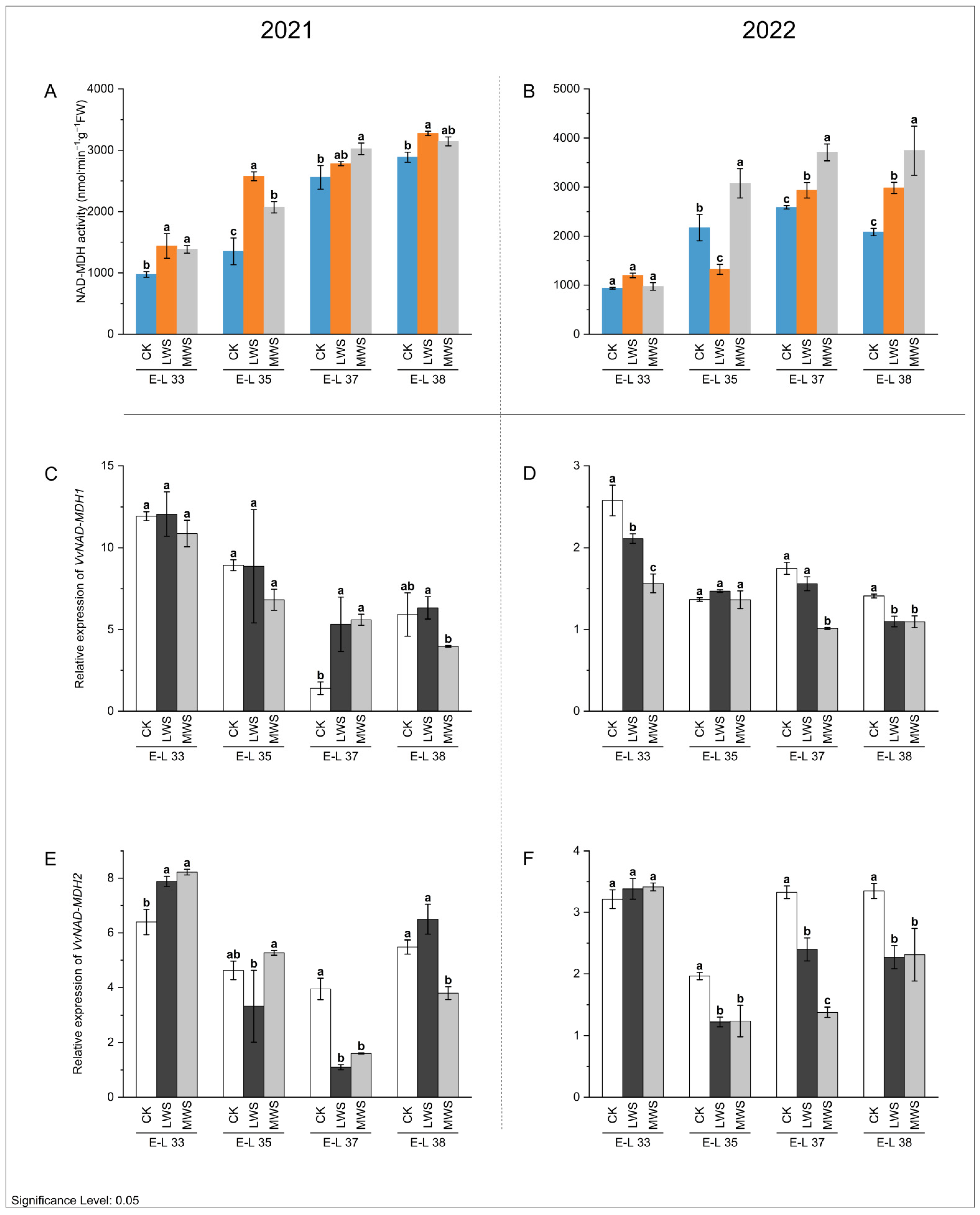
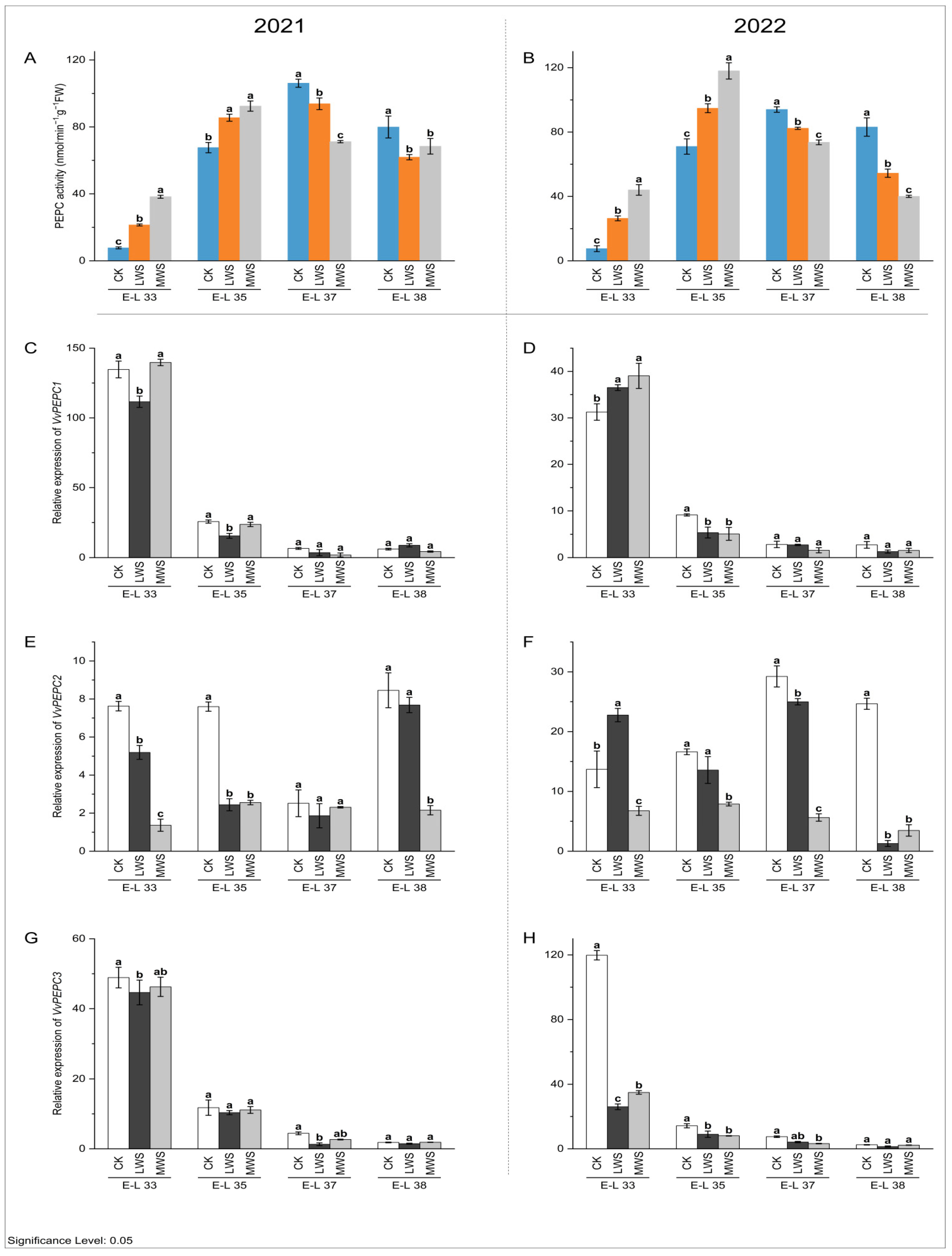
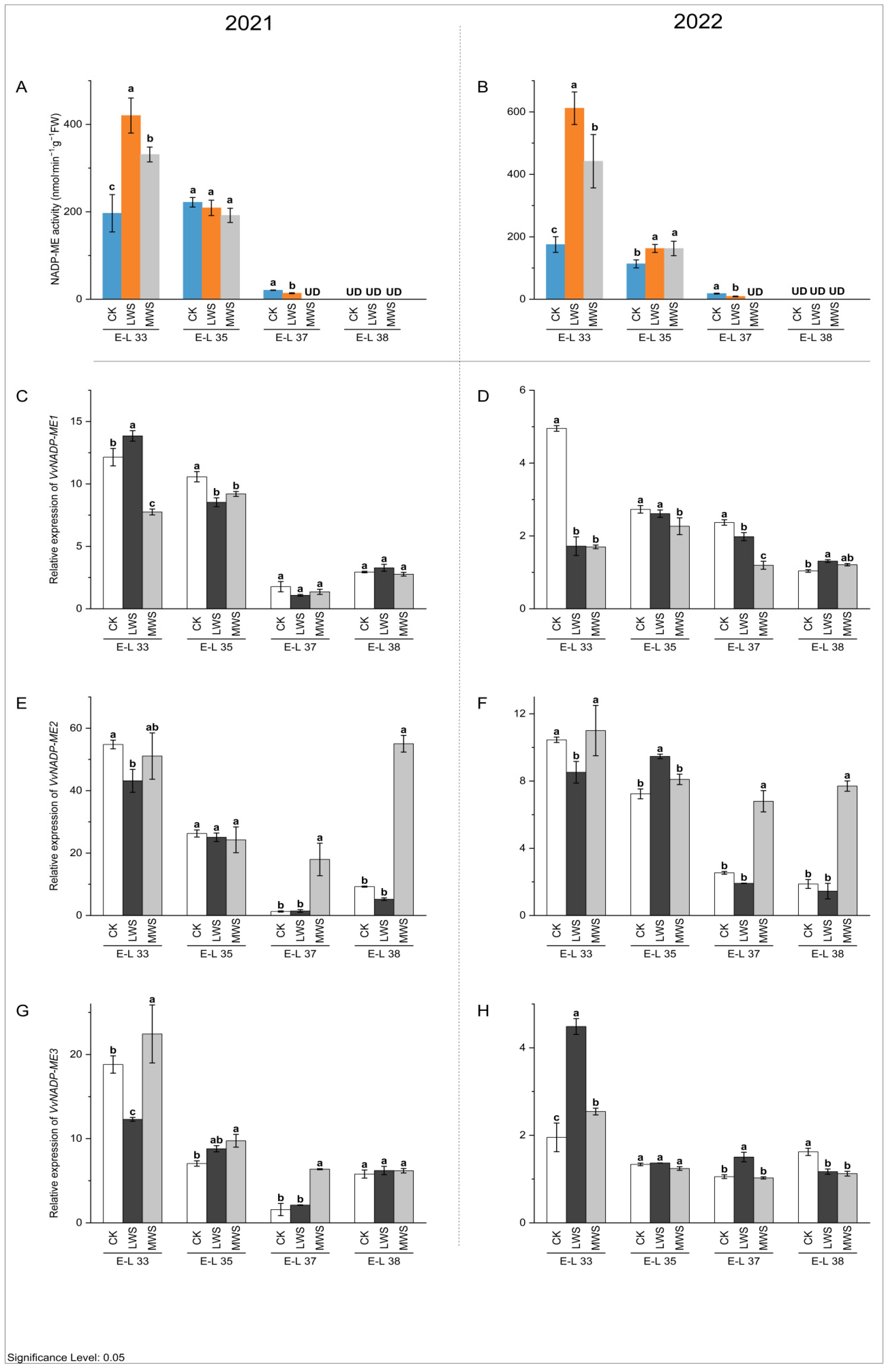
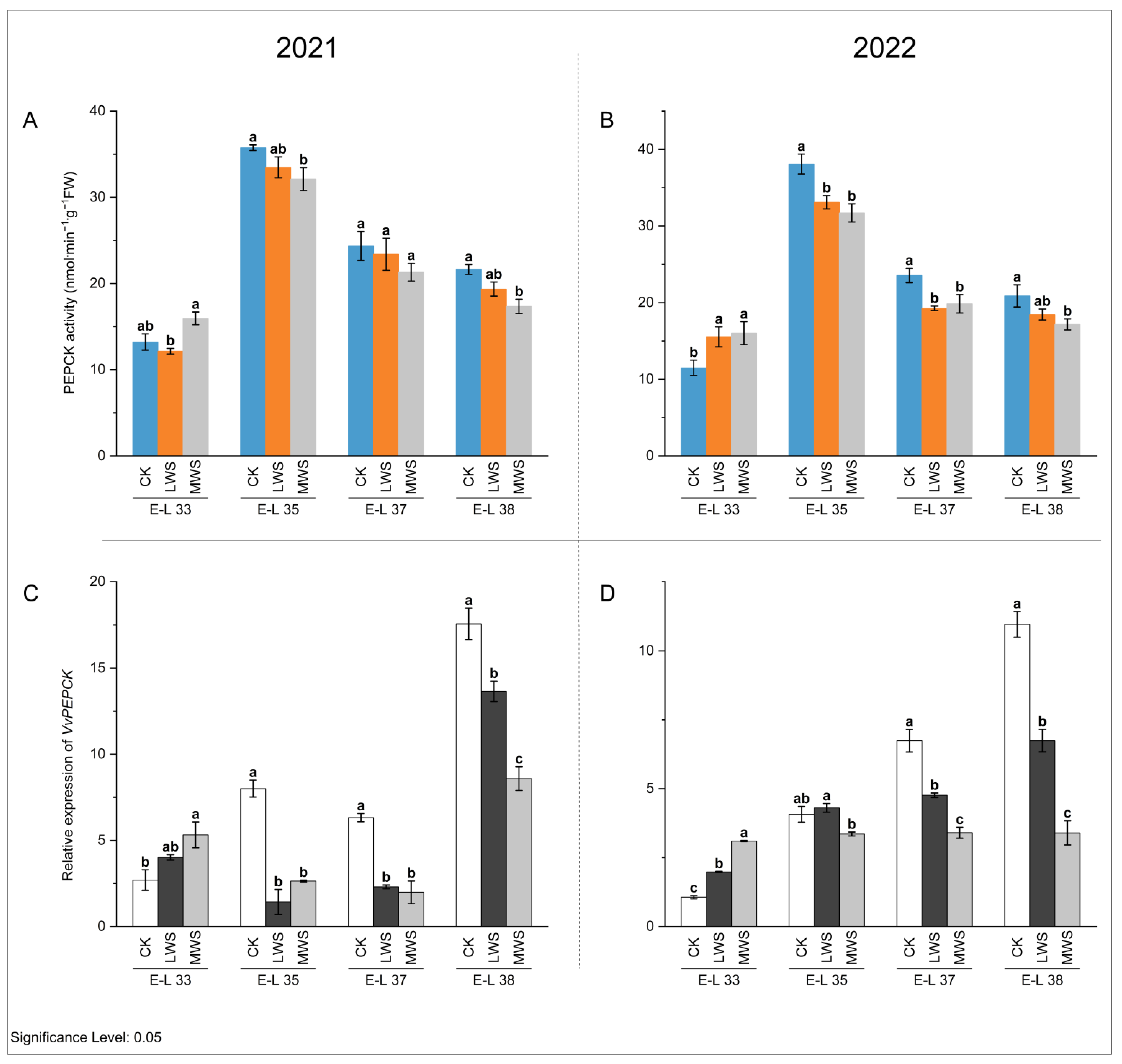
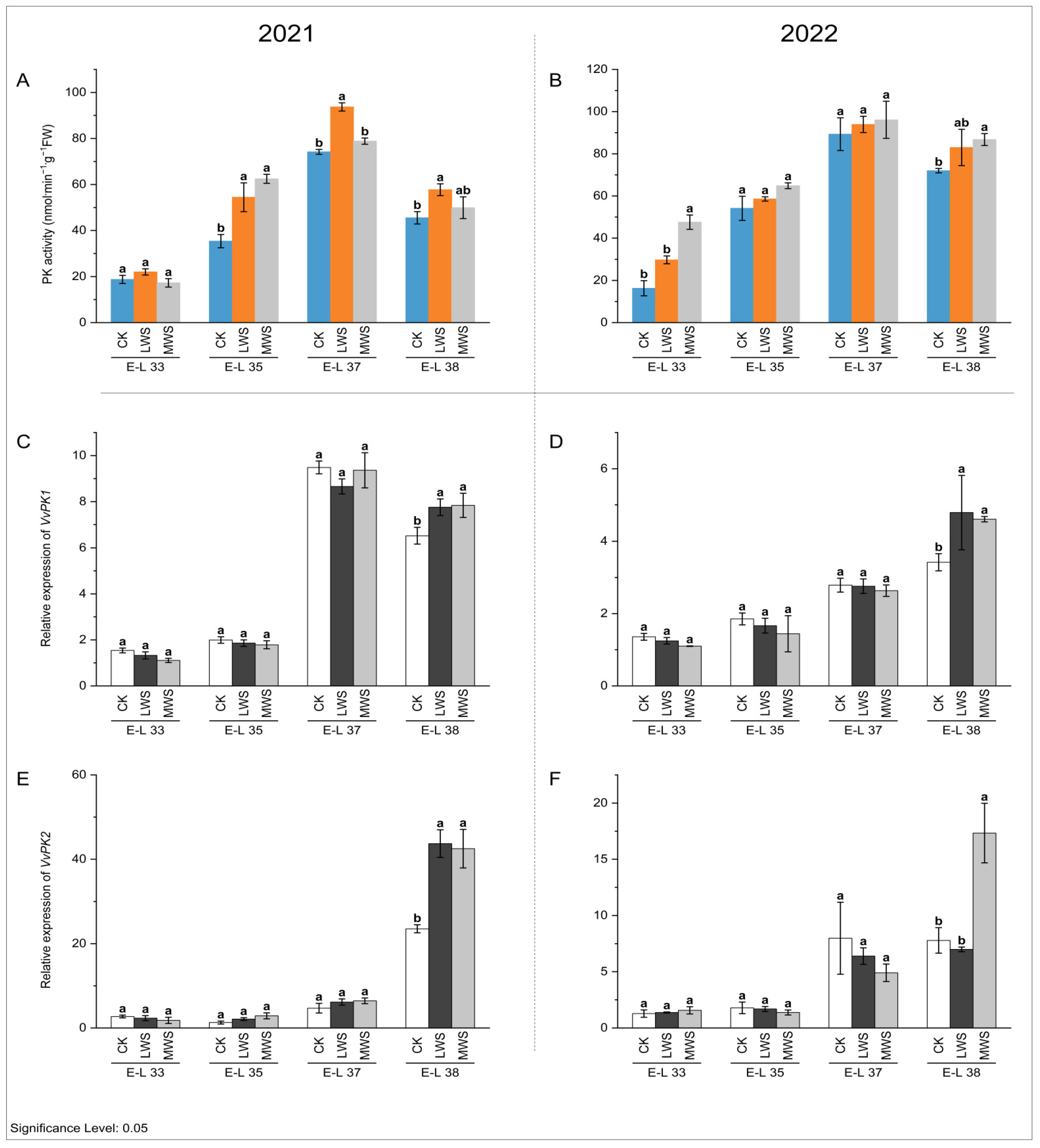
3.4. Malic Acid Metabolic Enzyme Activity and Gene Expression under Water Stress
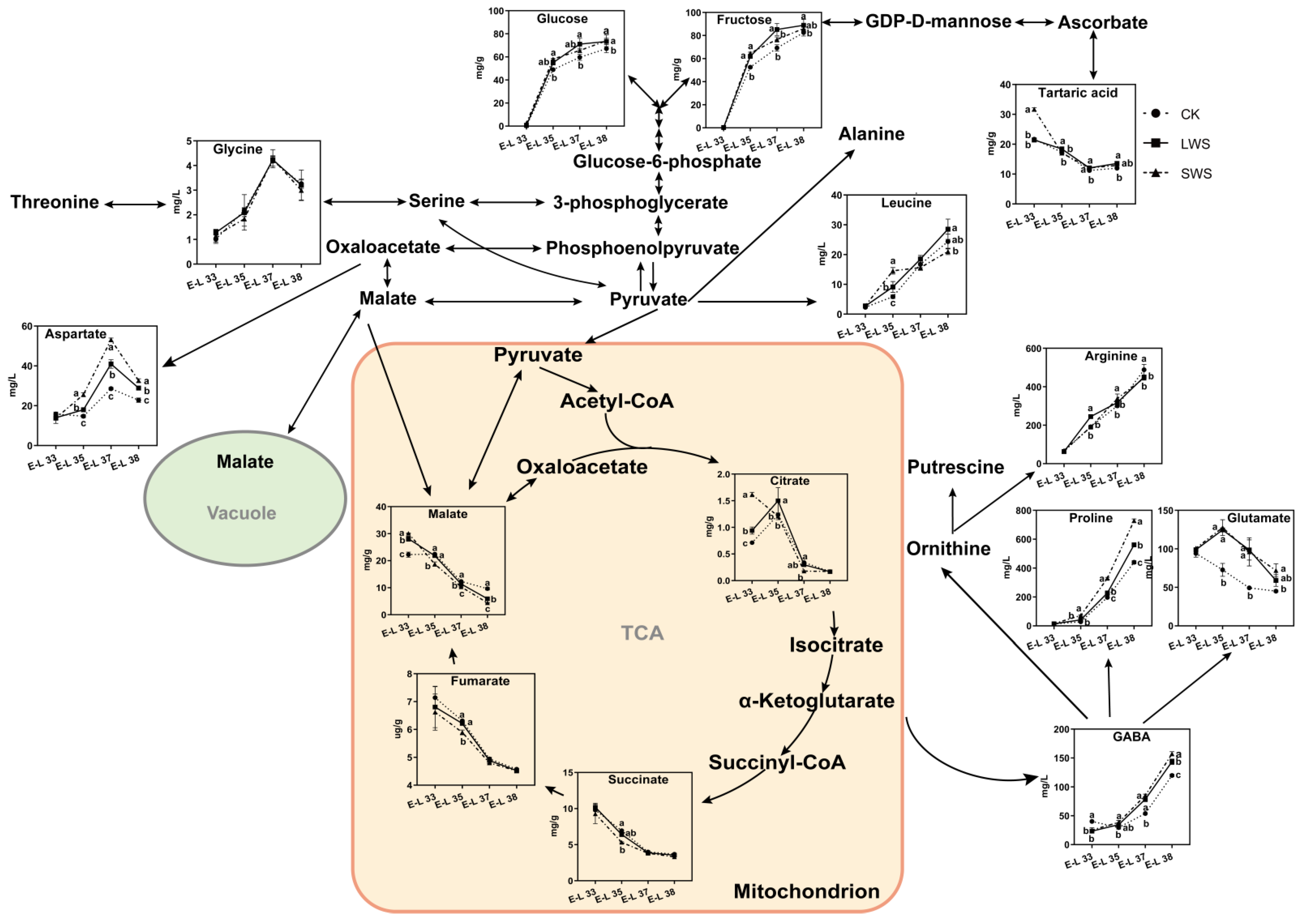
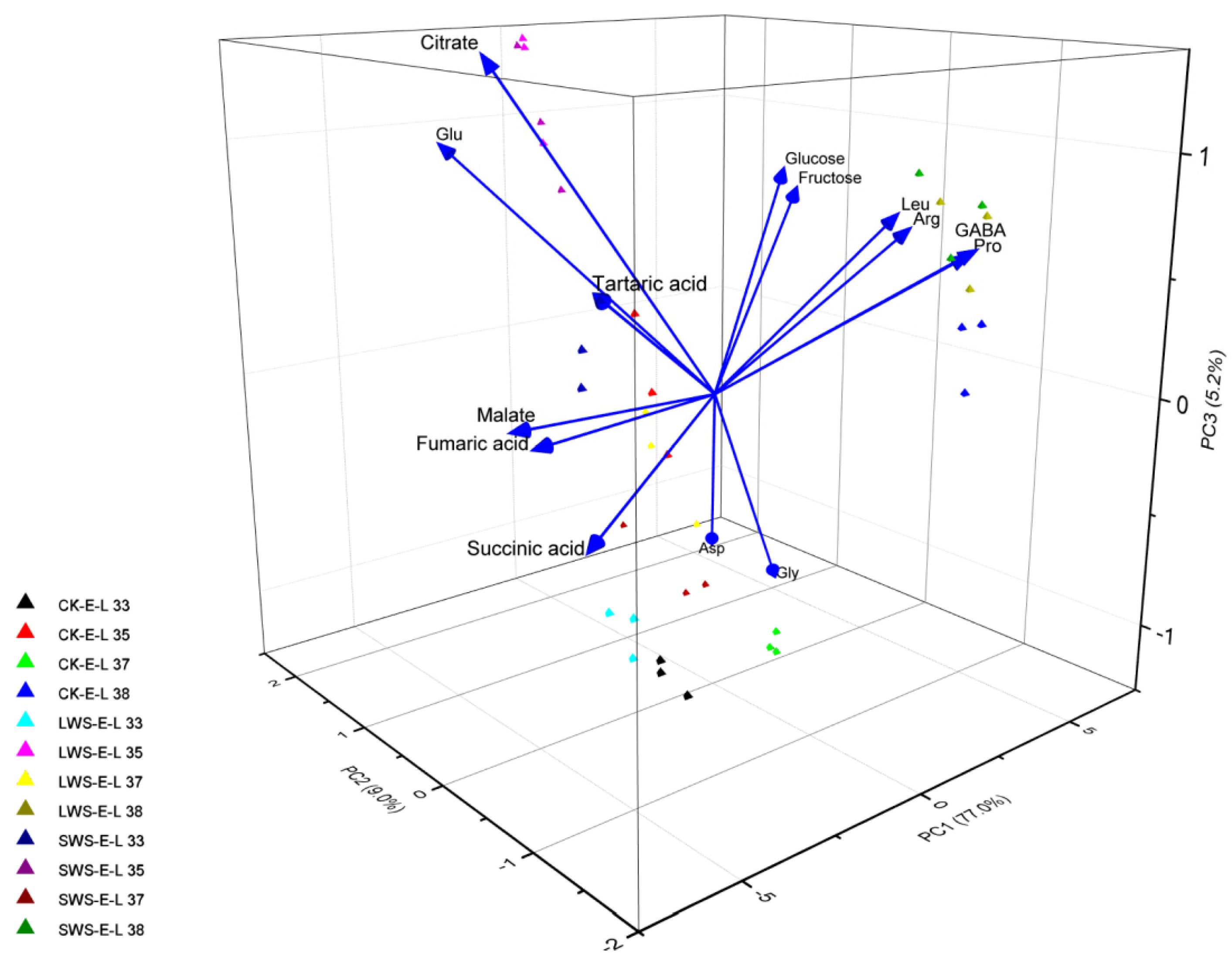
3.5. Differential Metabolites Affected by Water Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Umer, M.J.; Bin Safdar, L.; Gebremeskel, H.; Zhao, S.; Yuan, P.; Zhu, H.; Kaseb, M.O.; Anees, M.; Lu, X.; He, N.; et al. Identification of key gene networks controlling organic acid and sugar metabolism during watermelon fruit development by integrating metabolic phenotypes and gene expression profiles. Hortic. Res. 2020, 7, 193. [Google Scholar] [CrossRef] [PubMed]
- Sweetman, C.; Deluc, L.G.; Cramer, G.R.; Ford, C.M.; Soole, K.L. Regulation of malate metabolism in grape berry and other developing fruits. Phytochemistry 2009, 70, 1329–1344. [Google Scholar] [CrossRef]
- Zhang, X.; Kontoudakis, N.; Šuklje, K.; Antalick, G.; Blackman, J.W.; Rutledge, D.N.; Schmidtke, L.M.; Clark, A.C. Changes in Red Wine Composition during Bottle Aging: Impacts of Grape Variety, Vineyard Location, Maturity, and Oxygen Availability during Aging. J. Agric. Food Chem. 2020, 68, 13331–13343. [Google Scholar] [CrossRef] [PubMed]
- Levin, A.D.; Deloire, A.; Gambetta, G.A. Does water deficit negatively impact wine grape yield over the long-term? IVES Tech. Rev. 2020, 4029. [Google Scholar] [CrossRef]
- Wei, Z.; Luo, J.; Huang, Y.; Guo, W.; Zhang, Y.; Guan, H.; Xu, C.; Lu, J. Profile of Polyphenol Compounds of Five Muscadine Grapes Cultivated in the United States and in Newly Adapted Locations in China. Int. J. Mol. Sci. 2017, 18, 631. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Yao, H.; Zhang, J.X.; Li, Y.; Fang, Y. Effect of regulated deficit irrigation on the content of soluble sugars, organic acids and endogenous hormones in cabernet sauvignon in the ningxia region of china. Food Chem. 2020, 312, 126020. [Google Scholar] [CrossRef] [PubMed]
- Leng, F.; Zhou, J.; Wang, C.; Sun, L.; Zhang, Y.; Li, Y.; Wang, L.; Wang, S.; Zhang, X.; Xie, Z. Post-veraison different frequencies of water deficit strategies enhance Reliance grapes quality under root restriction. Food Chem. 2022, 390, 133181. [Google Scholar] [CrossRef]
- Ma, W.; Lu, S.; Li, W.; Nai, G.; Ma, Z.; Li, Y.; Chen, B.; Mao, J. Transcriptome and metabolites analysis of water-stressed grape berries at different growth stages. Physiol. Plant. 2023, 175, e13910. [Google Scholar] [CrossRef]
- Jan, N.; Anjum, S.; Wani, S.M.; Mir, S.A.; Malik, A.R.; Wani, S.A.; Hussein, D.S.; Rasheed, R.A.; Gatasheh, M.K. Influence of Canning and Storage on Physicochemical Properties, Antioxidant Properties, and Bioactive Compounds of Apricot (Prunus armeniaca L.) Wholes, Halves, and Pulp. Front. Nutr. 2022, 9, 850730. [Google Scholar] [CrossRef]
- Bigard, A.; Romieu, C.; Sire, Y.; Veyret, M.; Ojeda, H.; Torregrosa, L. The kinetics of grape ripening revisited through berry density sorting. Oeno One 2019, 53, 709–724. [Google Scholar] [CrossRef]
- Wang, H.; Miao, Y.; Xu, X.; Ye, P.; Wu, H.; Wang, B.; Shi, X. Effects of Blending on Phenolic, Colour, Antioxidant and Aroma Components of Cabernet Sauvignon Wine from Xinjiang (China). Foods 2022, 11, 3332. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhong, H.; Zhou, X.; Pan, M.; Xu, J.; Liu, M.; Wang, M.; Liu, G.; Xu, T.; Wang, Y.; et al. Grafting with rootstocks promotes phenolic compound accumulation in grape berry skin during development based on integrative multi-omics analysis. Hortic. Res. 2022, 9, uhac055. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Yuan, Y.; Gao, M.; Li, C.; Ogutu, C.; Li, M.; Ma, F. Determination of Predominant Organic Acid Components in Malus Species: Correlation with Apple Domestication. Metabolites 2018, 8, 74. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Wang, P.; Zhang, J.; Guo, X.; Mu, X.; Du, J. Organic acid metabolism in Chinese dwarf cherry [Cerasus humilis (Bge.) Sok.] is controlled by a complex gene regulatory network. Front. Plant Sci. 2022, 13, 982112. [Google Scholar] [CrossRef]
- Uriarte, D.; Intrigliolo, D.S.; Mancha, L.A.; Valdés, E.; Gamero, E.; Prieto, M.H. Combined effects of irrigation regimes and crop load on ‘Tempranillo’ grape composition. Agric. Water Manag. 2016, 165, 97–107. [Google Scholar] [CrossRef]
- Gambetta, G.A.; Herrera, J.C.; Dayer, S.F.; Feng, Q.; Hochbergm, U.; Castellarin, S.D. The physiology of drought stress in grapevine: Towards an integrative definition of drought tolerance. J. Exp. Bot. 2020, 71, 4658–4676. [Google Scholar] [CrossRef]
- Chen, Y.; Zeng, L.; Liao, Y.; Li, J.; Zhou, B.; Yang, Z.; Tang, J. Enzymatic Reaction-Related Protein Degradation and Proteinaceous Amino Acid Metabolism during the Black Tea (Camellia sinensis) Manufacturing Process. Foods 2020, 9, 66. [Google Scholar] [CrossRef]
- Araujo, W.L.; Nunes-Nesi, A.; Sweetlove, L.J.; Fernie, A.R. Metabolic control and regulation of the tricarboxylic acid cycle in photosynthetic and heterotrophic plant tissues. Plant Cell Environ. 2012, 35, 1–21. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; Shao, H.; Qi, W.; Hamoud, Y.A.; Shaghaleh, H.; Khan, N.U.; Yang, R.; Tang, B. GABA-Alleviated Oxidative Injury Induced by Salinity, Osmotic Stress and their Combination by Regulating Cellular and Molecular Signals in Rice. Int. J. Mol. Sci. 2019, 20, 5709. [Google Scholar] [CrossRef]
- Wang, Z.P.; Deloire, A.; Carbonneau, A.; Federspiel, B.; Lopez, F. An in vivo experimental system to study sugar phloem unloading in ripening grape berries during water deficiency stress. Ann. Bot. 2003, 92, 523–528. [Google Scholar] [CrossRef]
- Coombe, B.G. Adoption of a system for identifying grapevine growth stages. Aust. J. Grape Wine Res. 1995, 1, 104–110. [Google Scholar] [CrossRef]
- Deloire, A.; Pellegrino, A.; Rogiers, S. A few words on grapevine leaf water potential. IVES Tech. Rev. Vine Wine 2020. [Google Scholar] [CrossRef]
- Lian, T.T.; Moe, M.M.; Kim, Y.J.; Bang, K.S. Effects of Different Colored LEDs on the Enhancement of Biologically Active Ingredients in Callus Cultures of Gynura procumbens (Lour.) Merr. Molecules 2019, 24, 4336. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Yue, Q.; Bian, F.; Sun, H.; Zhai, H.; Yao, Y. Melatonin enhances phenolics accumulation partially via ethylene signaling and resulted in high antioxidant capacity in grape berries. Front. Plant Sci. 2017, 8, 1426. [Google Scholar] [CrossRef]
- Wu, Z.F.; Tu, M.M.; Yang, X.P.; Xu, J.H.; Yu, Z.F. Effect of cutting and storage temperature on sucrose and organic acids metabolism in postharvest melon fruit. Postharvest Biol. Technol. 2020, 161, 111081. [Google Scholar] [CrossRef]
- Sweetman, C.; Sadras, V.O.; Hancock, R.D.; Soole, K.L.; Ford, C.M. Metabolic effects of elevated temperature on organic acid degradation in ripening Vitis vinifera fruit. J. Exp. Bot. 2014, 65, 5975–5988. [Google Scholar] [CrossRef]
- Hirai, M.; Ueno, I. Development of citrus fruits: Fruit development and enzymatic changes in juice vesicle tissue. Plant Cell Physiol. 1977, 18, 791–799. [Google Scholar] [CrossRef]
- Han, S.; Nan, Y.; Qu, W.; He, Y.; Ban, Q.; Lv, Y.; Rao, J. Exogenous γ-Aminobutyric Acid Treatment That Contributes to Regulation of Malate Metabolism and Ethylene Synthesis in Apple Fruit during Storage. J. Agric. Food Chem. 2018, 66, 13473–13482. [Google Scholar] [CrossRef]
- Zhan, Z.N.; Wang, N.; Chen, Z.M.; Zhang, Y.X.; Geng, K.Q.; Li, D.M.; Wang, Z.P. Effects of water stress on endogenous hormones and free polyamines in different tissues of grapevines (Vitis vinifera L. cv. ‘Merlot’). Funct. Plant Biol. 2023. online ahead of print. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Ye, D.Q.; Zhu, B.Q.; Wu, G.F.; Duan, C.Q. Rapid HPLC analysis of amino acids and biogenic amines in wines during fermentation and evaluation of matrix effect. Food Chem. 2014, 163, 6–15. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Etienne, A.; Génard, M.; Lobit, P.; Mbeguié-A-Mbéguié, D.; Bugaud, C. What controls fleshy fruit acidity? A review of malate and citrate accumulation in fruit cells. J. Exp. Bot. 2013, 64, 1451–1469. [Google Scholar] [CrossRef] [PubMed]
- Garrido, I.; Uriarte, D.; Hernández, M.; Llerena, J.L.; Valdés, M.E.; Espinosa, F. The Evolution of Total Phenolic Compounds and Antioxidant Activities during Ripening of Grapes (Vitis vinifera L., cv. Tempranillo) Grown in Semiarid Region: Effects of Cluster Thinning and Water Deficit. Int. J. Mol. Sci. 2016, 17, 1923. [Google Scholar] [CrossRef] [PubMed]
- Picard, M.; van Leeuwen, C.; Guyon, F.; Gaillard, L.; de Revel, G.; Marchand, S. Vine Water Deficit Impacts Aging Bouquet in Fine Red Bordeaux Wine. Front. Chem. 2017, 5, 56. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Kurtural, S.K. Proximal Sensing of Soil Electrical Conductivity Provides a Link to Soil-Plant Water Relationships and Supports the Identification of Plant Water Status Zones in Vineyards. Front. Plant Sci. 2020, 11, 244. [Google Scholar] [CrossRef]
- Pellegrino, A.; Clingeleffer, P.; Cooley, N.; Walker, R. Management practices impact vine carbohydrate status to a greater extent than vine productivity. Front. Plant Sci. 2014, 5, 283. [Google Scholar] [CrossRef]
- Blancquaert, E.H.; Oberholster, A.; Ricardo-Da-Silva, J.M.; Deloire, A.J. Effects of Abiotic Factors on Phenolic Compounds in the Grape Berry—A Review. S. Afr. J. Enol. Vitic. 2018, 40, 3060. [Google Scholar] [CrossRef]
- Deloire, A.; Pellegrino, A. Review of vine water deficit. What levers for the vineyard in the short and medium term? IVES Tech. Rev. Vine Wine 2021, 9, 4842. [Google Scholar] [CrossRef]
- Wang, X.; Tu, M.; Wang, D.; Liu, J.; Li, Y.; Li, Z.; Wang, Y.; Wang, X. CRISPR/Cas9-mediated efficient targeted mutagenesis in grape in the first generation. Plant Biotechnol. J. 2018, 16, 844–855. [Google Scholar] [CrossRef]
- Shahood, R.; Torregrosa, L.; Savoi, S.; Romieu, C. First quantitative assessment of growth, sugar accumulation and malate breakdown in a single ripening berry. Oeno One 2020, 4, 1077–1092. [Google Scholar] [CrossRef]
- Ruffner, H.P.; Hawker, J.S.; Hale, C.R. Temperature and enzymic control of malate metabolism in berries of vitis vinifera. Phytochemistry 1976, 15, 1877–1880. [Google Scholar] [CrossRef]
- Ford, C.M. The Biochemistry of organic acids in the Grape. In The Biochemistry of the Grape Berry; Geròs, H., Chaves, M.M., Delrot, S., Eds.; Bentham Books: Dubai, United Arab Emirates, 2012; pp. 67–88. [Google Scholar] [CrossRef]
- Rienth, M.; Torregrosa, L.; Sarah, G.; Ardisson, M.; Brillouet, J.M.; Romieu, C. Temperature desynchronizes sugar and organic acid metabolism in ripening grapevine fruits and remodels their transcriptome. BMC Plant Biol. 2016, 16, 164. [Google Scholar] [CrossRef] [PubMed]
- Lecourieux, F.; Kappel, C.; Pieri, P.; Charon, J.; Pillet, J.; Hilbert, G.; Renaud, C.; Gomès, E.; Delrot, S.; Lecourieux, D. Dissecting the Biochemical and Transcriptomic Effects of a Locally Applied Heat Treatment on Developing Cabernet Sauvignon Grape Berries. Front. Plant Sci. 2017, 8, 53. [Google Scholar] [CrossRef] [PubMed]
- Cooley, N.M.; Clingeleffer, P.R.; Walker, R.R. Effect of water deficits and season on berry development and composition of Cabernet Sauvignon (Vitis vinifera L.) grown in a hotclimate. Aust. J. Grape Wine Res. 2017, 23, 260–272. [Google Scholar] [CrossRef]
- Savoi, S.; Wong, D.C.J.; Degu, A.; Herrera, J.C.; Bucchetti, B.; Peterlunger, E.; Fait, A.; Mattivi, F.; Castellarin, S.D. Multi-Omics and Integrated Network Analyses Reveal New Insights into the Systems Relationships between Metabolites, Structural Genes, and Transcriptional Regulators in Developing Grape Berries (Vitis vinifera L.) Exposed to Water Deficit. Front. Plant Sci. 2017, 8, 1124. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.M.; Anwar, R.; Rehman, R.N.U.; Ejaz, S.; Ali, S.; Yousef, A.F.; Ercisli, S.; Hu, X.; Hou, Y.; Chen, F. Sugar and acid profile of loquat (Eriobotrya japonica Lindl.), enzymes assay and expression profiling of their metabolism-related genes as influenced by exogenously applied boron. Front. Plant Sci. 2022, 13, 1039360. [Google Scholar] [CrossRef]
- Chen, Y.; Fu, Z.; Zhang, H.; Tian, R.; Yang, H.; Sun, C.; Wang, L.; Zhang, W.; Guo, Z.; Zhang, X.; et al. Cytosolic malate dehydrogenase 4 modulates cellular energetics and storage reserve accumulation in maize endosperm. Plant Biotechnol. J. 2020, 18, 2420–2435. [Google Scholar] [CrossRef]
- Yao, Y.X.; Li, M.; Zhai, H.; You, C.X.; Hao, Y.J. Isolation and characterization of an apple cytosolic malate dehydrogenase gene reveal its function in malate synthesis. J. Plant Physiol. 2011, 168, 474–480. [Google Scholar] [CrossRef]
- Iannetta, P.P.; Escobar, N.M.; Ross, H.A.; Souleyre, E.J.; Hancock, R.D.; Witte, C.P.; Davies, H.V. Identification, cloning and expression analysis of strawberry (Fragaria x ananassa) mitochondrial citrate synthase and mitochondrial malate dehydrogenase. Physiol. Plant. 2010, 121, 15–26. [Google Scholar] [CrossRef]
- Wang, Q.J.; Sun, H.; Dong, Q.L.; Sun, T.Y.; Jin, Z.X.; Hao, Y.J.; Yao, Y.X. The enhancement of tolerance to salt and cold stresses by modifying the redox state and salicylic acid content via the cytosolic malate dehydrogenase gene in transgenic apple plants. Plant Biotechnol. J. 2016, 14, 1986–1997. [Google Scholar] [CrossRef]
- Huang, J.; Niazi, A.K.; Young, D.; Rosado, L.A.; Vertommen, D.; Bodra, N.; Abdelgawwad, M.R.; Vignols, F.; Wei, B.; Wahni, K.; et al. Self-protection of cytosolic malate dehydrogenase against oxidative stress in Arabidopsis. J. Exp. Bot. 2018, 69, 3491–3505. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.C.; Fang, Z.Z.; Zhou, D.R.; Pan, S.L.; Ye, X.F. Changes in secondary metabolites, organic acids and soluble sugars during the development of plum fruit cv. ‘Furongli’ (Prunus salicina Lindl). J. Sci. Food Agric. 2019, 99, 1010–1019. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.X.; Liu, X.H.; Chen, L.S. Developmental changes in pulp organic acid concentration and activities of acid-metabolising enzymes during the fruit development of two loquat (Eriobotrya japonica Lindl.) cultivars differing in fruit acidity. Food Chem. 2009, 114, 657–664. [Google Scholar] [CrossRef]
- Zhang, X.M.; Du, L.Q.; Sun, G.M.; Gong, D.Q.; Chen, J.Y.; Li, W.C.; Xie, J.H. Changes in organic acid concentrations and the relative enzyme activities during the development of Cayenne pineapple fruit. J. Fruit Sci. 2007, 24, 381–384. [Google Scholar]
- Diakou, P.; Svanella, L.; Raymond, P.; Gaudillère, J.P.; Moing, A. Phosphoenolpyruvate carboxylase during grape berry development: Protein level, enzyme activity and regulation. Aust. J. Plant Physiol. 2000, 27, 221–229. [Google Scholar] [CrossRef]
- Liu, L.Y. Study on the Accumulation Rule and Metabolic Regulation Mechanism of Glucuronic Acid. Ph.D. Thesis, The Northwest A & F University, Shanxi, China, 2016; p. 55. [Google Scholar]
- Ruffner, H.P. Metabolism of tartaric and malic acids in Vitis: A review—Part B. Vitis 1982, 21, 346–358. [Google Scholar] [CrossRef]
- Or, E.; Baybik, J.; Sadka, A.; Saks, Y. Isolation of mitochondrial malate dehydrogenase and phosphoenolpyruvate carboxylase cDNA clones from grape berries and analysis of their expression pattern throughout berry development. J. Plant Physiol. 2000, 157, 527–534. [Google Scholar] [CrossRef]
- Yao, Y.X.; Li, M.; Liu, Z.; You, C.X.; Wang, D.M.; Zhai, H.; Hao, Y.J. Molecular cloning of three malic acid related genes MdPEPC, MdVHA-A, MdcyME and their expression analysis in apple fruits. Sci. Hortic. 2009, 122, 404–408. [Google Scholar] [CrossRef]
- Hang, J.; Chen, Y.; Liu, L.; Chen, L.; Fang, J.; Wang, F.; Wang, M. Antitumor effect and metabonomics of niclosamide micelles. J. Cell. Mol. Med. 2022, 26, 4814–4824. [Google Scholar] [CrossRef]
- Teixeira, A.; Martins, V.; Noronha, H.; Eiras-Dias, J.; Gerós, H. The first insight into the metabolite profiling of grapes from three Vitis vinifera L. cultivars of two controlled appellation (DOC) regions. Int. J. Mol. Sci. 2014, 15, 4237–4254. [Google Scholar] [CrossRef]
- Clément, G.; Moison, M.; Soulay, F.; Reisdorf-Cren, M.; Masclaux-Daubresse, C. Metabolomics of laminae and midvein during leaf senescence and source-sink metabolite management in Brassica napus L. leaves. J. Exp. Bot. 2018, 69, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Yong, B.; Xie, H.; Li, Z.; Li, Y.P.; Zhang, Y.; Nie, G.; Zhang, X.Q.; Ma, X.; Huang, L.K.; Yan, Y.H.; et al. Exogenous Application of GABA Improves PEG-Induced Drought Tolerance Positively Associated with GABA-Shunt, Polyamines, and Proline Metabolism in White Clover. Front. Physiol. 2017, 8, 1107. [Google Scholar] [CrossRef] [PubMed]
- Ngaffo Mekontso, F.; Duan, W.; Cisse, E.; Chen, T.; Xu, X. Alleviation of Postharvest Chilling Injury of Carambola Fruit by γ-aminobutyric Acid: Physiological, Biochemical, and Structural Characterization. Front. Nutr. 2021, 8, 752583. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Berry Fresh Mass (g) of 50 Berries | TSS (°Brix) | Titratable Acidity (g/L) | Total Anthocyanins Content (mg/g) | |
|---|---|---|---|---|---|
| 2021 | CK | 50.79 ± 1.39 a | 22.77 ± 0.12 b | 9.41 ± 0.37 a | 3.49 ± 0.24 b |
| LWS | 36.11 ± 0.86 b | 23.27 ± 0.09 a | 7.41 ± 0.24 b | 4.29 ± 0.28 a | |
| MWS | 30.39 ± 0.49 c | 23.81 ± 0.93 a | 7.22 ± 0.22 b | 4.44 ± 0.31 a | |
| 2022 | CK | 60.20 ± 0.39 a | 24.77 ± 0.45 c | 8.40 ± 0.21 a | 4.22 ± 0.30 c |
| LWS | 50.79 ± 1.18 b | 26.30 ± 0.33 b | 6.39 ± 0.17 b | 5.23 ± 0.07 b | |
| MWS | 46.53 ± 0.80 c | 27.23 ± 0.25 a | 5.48 ± 0.32 c | 6.12 ± 0.33 a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhan, Z.; Zhang, Y.; Geng, K.; Xue, X.; Deloire, A.; Li, D.; Wang, Z. Effects of Vine Water Status on Malate Metabolism and γ-Aminobutyric Acid (GABA) Pathway-Related Amino Acids in Marselan (Vitis vinifera L.) Grape Berries. Foods 2023, 12, 4191. https://doi.org/10.3390/foods12234191
Zhan Z, Zhang Y, Geng K, Xue X, Deloire A, Li D, Wang Z. Effects of Vine Water Status on Malate Metabolism and γ-Aminobutyric Acid (GABA) Pathway-Related Amino Acids in Marselan (Vitis vinifera L.) Grape Berries. Foods. 2023; 12(23):4191. https://doi.org/10.3390/foods12234191
Chicago/Turabian StyleZhan, Zhennan, Yanxia Zhang, Kangqi Geng, Xiaobin Xue, Alain Deloire, Dongmei Li, and Zhenping Wang. 2023. "Effects of Vine Water Status on Malate Metabolism and γ-Aminobutyric Acid (GABA) Pathway-Related Amino Acids in Marselan (Vitis vinifera L.) Grape Berries" Foods 12, no. 23: 4191. https://doi.org/10.3390/foods12234191
APA StyleZhan, Z., Zhang, Y., Geng, K., Xue, X., Deloire, A., Li, D., & Wang, Z. (2023). Effects of Vine Water Status on Malate Metabolism and γ-Aminobutyric Acid (GABA) Pathway-Related Amino Acids in Marselan (Vitis vinifera L.) Grape Berries. Foods, 12(23), 4191. https://doi.org/10.3390/foods12234191