
Valorisation of Three Underutilised Native Australian Plants: Phenolic and Organic Acid Profiles and In Vitro Antimicrobial Activity

 , ,
, , 
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material and Extraction
2.3. Sugar Analysis
2.4. Vitamin C Analysis
2.5. Organic Acid Analysis
2.6. Total Phenolic Content (TPC)
2.7. DPPH Radical Scavenging Capacity
2.8. HRAM Analysis and Tentative Identification of Non-Anthocyanin Phenolic Compounds
2.9. Antimicrobial Activity
2.9.1. Design of Experiments
2.9.2. Agar Well Diffusion Assay
2.10. Statistical Analysis
3. Results and Discussion
3.1. Chemical Composition
3.2. Identification of Non-Anthocyanin Polyphenols
3.2.1. Phenolic Acids
3.2.2. Flavonoids
3.2.3. Flavonoid Glycosides
3.2.4. Polyflavonoids
3.2.5. Tannins
3.2.6. Other Polyphenols
3.3. Antimicrobial Activity of Extract Blends
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alderees, F.; Akter, S.; Mereddy, R.; Sultanbawa, Y. Antimicrobial activity of nanoencapsulated essential oils of Tasmannia lanceolata, Backhousia citriodora and Syzygium anisatum against weak-acid resistant Zygosaccharomyces bailii in clear apple juice. Beverages 2021, 7, 67. [Google Scholar] [CrossRef]
- Bobasa, E.M.; Phan, A.D.T.; Netzel, M.E.; Cozzolino, D.; Sultanbawa, Y. Hydrolysable tannins in Terminalia ferdinandiana Exell fruit powder and comparison of their functional properties from different solvent extracts. Food Chem. 2021, 358, 129833. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.K. Syzygium aqueum. In Edible Medicinal and Non Medicinal Plants: Fruits; Springer: Dordrecht, The Netherlands, 2012; Volume 3, pp. 738–742. [Google Scholar]
- Kupsch, K. Diploglottis. Australian Bushfoods Magazine. Available online: https://www.ausbushfoods.com/bushfoodsonline/backissues/Issue15/Issue_15_2.htm (accessed on 17 November 2021).
- Winnett, V.; Boyer, H.; Sirdaarta, J.; Cock, I. The potential of tasmannia lanceolata as a natural preservative and medicinal agent: Antimicrobial activity and toxicity. Pharmacogn. Commun. 2014, 4, 42–52. [Google Scholar] [CrossRef]
- Cock, I. Medicinal and aromatic plants—Australia. In Ethnopharmacology, Encyclopedia of Life Support Systems (EOLSS); UNESCO: Oxford, UK, 2011. [Google Scholar]
- Bouarab Chibane, L.; Degraeve, P.; Ferhout, H.; Bouajila, J.; Oulahal, N. Plant antimicrobial polyphenols as potential natural food preservatives. J. Sci. Food Agric. 2019, 99, 1457–1474. [Google Scholar] [CrossRef]
- Park, C.H.; Yeo, H.J.; Baskar, T.B.; Park, Y.E.; Park, J.S.; Lee, S.Y.; Park, S.U. In Vitro Antioxidant and Antimicrobial Properties of Flower, Leaf, and Stem Extracts of Korean Mint. Antioxidants 2019, 8, 75. [Google Scholar] [CrossRef]
- Bobinaitė, R.; Viškelis, P.; Sarkinas, A.; Venskutonis, P. Phytochemical composition, antioxidant and antimicrobial properties of raspberry fruit, pulp, and marc extracts. CyTA—J. Food 2013, 11, 334–342. [Google Scholar] [CrossRef]
- Boudet, A.-M. Evolution and current status of research in phenolic compounds. Phytochemistry 2007, 68, 2722–2735. [Google Scholar] [CrossRef]
- Deng, Y.; Zhao, Y.; Padilla-Zakour, O.; Yang, G. Polyphenols, antioxidant and antimicrobial activties of leaf and bark extracts of Solidago canadensis L. Ind. Crops Prod. 2015, 74, 803–809. [Google Scholar] [CrossRef]
- Yang, D.; Chen, Y.; Guo, F.; Huang, B.; Okyere, S.A.; Wang, H. Comparative analysis of chemical composition, antioxidant and antimicrobial activties of leaves, leaf tea and root from Codonopsis pilosula. Ind. Crops Prod. 2019, 142, 111844. [Google Scholar] [CrossRef]
- Hong, H.T.; Phan, A.D.T.; O’Hare, T.J. Temperature and Maturity Stages Affect Anthocyanin Development and Phenolic and Sugar Content of Purple-Pericarp Supersweet Sweetcorn during Storage. J. Agric. Food Chem. 2021, 69, 922–931. [Google Scholar] [CrossRef]
- Phan, A.D.T.; Chaliha, M.; Sultanbawa, Y.; Netzel, M.E. Nutritional Characteristics and Antimicrobial Activity of Australian Grown Feijoa (Acca sellowiana). Foods 2019, 8, 376. [Google Scholar] [CrossRef]
- Moldoveanu, S.C.; Poole, T.; Scott, W.A. An LC-MS Method for the Analysis of Some Organic Acids in Tobacco Leaf, Snus, and Wet Snuff. Contrib. Tob. Res. 2018, 28, 30–41. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Method Enzymol. 1999, 299, 152–178. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Hmdb-Database. The Human Metabolome Database. Available online: https://hmdb.ca/ (accessed on 21 November 2021).
- Djenane, D.; Yangüela, J.; Montañés, L.; Djerbal, M.; Roncalés, P. Antimicrobial activity of Pistacia lentiscus and Satureja montana essential oils against Listeria monocytogenes CECT 935 using laboratory media: Efficacy and synergistic potential in minced beef. Food Control 2011, 22, 1046–1053. [Google Scholar] [CrossRef]
- Shezi, S.; Magwaza, L.S.; Tesfay, S.Z.; Mditshwa, A. Biochemical changes in response to canopy position of avocado fruit (cv. ‘Carmen’ and ‘Hass’) during growth and development and relationship with maturity. Sci. Hortic. 2020, 265, 109227. [Google Scholar] [CrossRef]
- Blakey, R.J.; Tesfay, S.Z.; Bower, J.P. Changes in sugars, total protein, and oil in ‘Hass’ avocado (Parsea americana Mill.) fruit during ripening. J. Hortic. Sci. Biotechnol. 2012, 87, 381–387. [Google Scholar] [CrossRef]
- Sheng, X.; Jung, T.; Wesson, J.A.; Ward, M.D. Adhesion at calcium oxalate crystal surfaces and the effect of urinary constituents. Proc. Natl. Acad. Sci. USA 2004, 102, 267–272. [Google Scholar] [CrossRef]
- Ramos-Aguilar, A.L.; Ornelas-Paz, J.; Tapia-Vargas, L.M.; Ruiz-Cruz, S.; Gardea-Béjar, A.A.; Yahia, E.M.; Ornelas-Paz, J.D.J.; Pérez-Martínez, J.D.; Rios-Velasco, C.; Ibarra-Junquera, V. The importance of the bioactive compounds of avocado fruit (Persea americana Mill.) on human health. Biotecnia 2019, 21, 154–162. [Google Scholar] [CrossRef]
- Jang, S.-A.; Park, D.W.; Kwon, J.E.; Song, H.S.; Park, B.; Jeon, H.; Sohn, E.-H.; Koo, H.J.; Kang, S.C. Quinic acid inhibits vascular inflammation in TNF-α-stimulated vascular smooth muscle cells. Biomed. Pharmacother. 2017, 96, 563–571. [Google Scholar] [CrossRef]
- Yazdi, S.E.; Prinsloo, G.; Heyman, H.M.; Oosthuizen, C.B.; Klimkait, T.; Meyer, J.J.M. Anti-HIV-1 activity of quinic acid isolated from Helichrysum mimetes using NMR-based metabolomics and comutational analysis. S. Afr. J. Bot. 2019, 126, 328–339. [Google Scholar] [CrossRef]
- Alderees, F.; Mereddy, R.; Webber, D.; Nirmal, N.; Sultanbawa, Y. Mechanism of action against food spoilage yeasts and bioactivity of Tasmannia lanceloata, Backhousia citriodora and Syzygium anisatum plant solvent extracts. Foods 2018, 7, 179. [Google Scholar] [CrossRef] [PubMed]
- Netzel, M.; Netzel, G.; Tian, Q.; Schwartz, S.; Konczak, I. Native Australian fruits—A novel source of antioxidants for food. Innov. Food Sci. Emerg. Technol. 2007, 8, 339–346. [Google Scholar] [CrossRef]
- Konczak, I.; Zabaras, D.; Dunstan, M.; Aguas, P. Antioxidant capacity and hydrophilic phytochemicals in commercially grown native Australian fruits. Food Chem. 2010, 123, 1048–1054. [Google Scholar] [CrossRef]
- Jaakola, L.; Määttä-Riihinen, K.; Kärenlampi, S.; Hohtola, A. Activation of flavonoid biosynthesis by solar radiation in bilberry (Vaccinium myrtillus L.) leaves. Planta. 2004, 218, 721–728. [Google Scholar] [PubMed]
- Cheynier, V.; Comte, G.; Davies, K.M.; Lattanzio, V.; Martens, S. Plant phenolics: Recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [Google Scholar] [CrossRef]
- Jaiswal, R.; Müller, H.; Müller, A.; Karar, M.G.E.; Kuhnert, N. Identification and characterization of chlorogenic acids, chlorogenic acid glycosides and flavonoids from Lonicera henryi L. (Caprifoliaceae) leaves by LC–MS. Phytochemistry 2014, 108, 252–263. [Google Scholar] [CrossRef]
- Trujillo-Mayol, I.; Casas-Forero, N.; Pastene-Navarrete, E.; Silva, F.L.; Alarcón-Enos, J. Fractionation and Hydrolyzation of Avocado Peel Extract: Improvement of Antibacterial Activity. Antibiotics 2020, 10, 23. [Google Scholar] [CrossRef]
- Liu, S.; Marsol-Vall, A.; Laaksonen, O.; Kortesniemi, M.; Yang, B. Characterization and Quantification of Nonanthocyanin Phenolic Compounds in White and Blue Bilberry (Vaccinium myrtillus) Juices and Wines Using UHPLC-DAD−ESI-QTOF-MS and UHPLC-DAD. J. Agric. Food Chem. 2020, 68, 7734–7744. [Google Scholar] [CrossRef]
- Qian, W.; Wu, W.; Kang, Y.; Wang, Y.; Yang, P.; Deng, Y.; Ni, C.; Huang, J. Comprehensive identification of minor components and bioassay-guided isolation of an unusual antioxidant from Azolla imbricata using ultra-high performance liquid chromatography—Quadrupole time-of-flight mass spectrometry combined with multicomponent knockout and bioactivity evaluation. J. Chromatogr. A 2019, 1609, 460435. [Google Scholar] [CrossRef]
- Hong, Y.; Wang, Z.; Barrow, C.; Dunshea, F.; Suleria, H. High-Throughput Screening and Characterization of Phenolic Compounds in Stone Fruits Waste by LC-ESI-QTOF-MS/MS and Their Potential Antioxidant Activities. Antioxidants 2021, 10, 234. [Google Scholar] [CrossRef]
- Wang, H.; Ye, Y.h.; Wang, H.h.; Liu, J.; Liu, Y.j.; Jiang, B.W. HPLC-QTOF-MS/MS profiling, antioxidant, and α-glucosidase inhibitory activities of Pyracantha fortuneana fruit extracts. J. Food Biochem. 2019, 43, e12821. [Google Scholar] [CrossRef] [PubMed]
- Mallmann, L.P.; Tischer, B.; Vizzotto, M.; Rodrigues, E.; Manfroi, V. Comprehensive identification and quantification of unexploited phenolic compounds from red and yellow araçá (Psidium cattleianum Sabine) by LC-DAD-ESI-MS/MS. Food Res. Int. 2020, 131, 108978. [Google Scholar] [CrossRef]
- Li, H.; Zhang, D.; Tan, L.-H.; Yu, B.; Zhao, S.-P.; Cao, W.-G. Comparison of the antioxidant properties of various solvent extracts from Dipsacus asperoides and identification of phenolic compounds by LC-ESI-QTOF-MS–MS. South Afr. J. Bot. 2017, 109, 1–8. [Google Scholar] [CrossRef]
- Singh, D.; Siew, Y.-Y.; Chong, T.-I.; Yew, H.-C.; Ho, S.S.-W.; Lim, C.S.E.-S.; Tan, W.-X.; Neo, S.-Y.; Koh, H.-L. Identification of Phytoconstituents in Leea indica (Burm. F.) Merr. Leaves by High Performance Liquid Chromatography Micro Time-of-Flight Mass Spectrometry. Molecules 2019, 24, 714. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Yang, Q.-Q.; Chen, Z.; Zhang, J.-R.; Chen, Q.; Wang, Y.; Wei, X. Distinct Changes of Metabolic Profile and Sensory Quality during Qingzhuan Tea Processing Revealed by LC-MS-Based Metabolomics. J. Agric. Food Chem. 2020, 68, 4955–4965. [Google Scholar] [CrossRef] [PubMed]
- Olennikov, D.N.; Chirikova, N.K.; Kashchenko, N.I.; Nikolaev, V.M.; Kim, S.-W.; Vennos, C. Bioactive Phenolics of the Genus Artemisia (Asteraceae): HPLC-DAD-ESI-TQ-MS/MS Profile of the Siberian Species and Their Inhibitory Potential Against α-Amylase and α-Glucosidase. Front. Pharmacol. 2018, 9, 756. [Google Scholar] [CrossRef]
- Yapıcı, İ.; Altay, A.; Öztürk Sarıkaya, B.; Korkmaz, M.; Atila, A.; Gülçin, İ.; Köksal, E. In vitro antioxidant and cytotoxic activities of extracts of endemic Tanacetum erzincanense together with phenolic content by LC-ESI-QTOF-MS. Chem. Biodivers. 2021, 18, e2000812. [Google Scholar] [CrossRef]
- Li, J.; Wang, Y.-H.; Smillie, T.J.; Khan, I.A. Identification of phenolic compounds from Scutellaria lateriflora by liquid chromatography with ultraviolet photodiode array and electrospray ionization tandem mass spectrometry. J. Pharm. Biomed. Anal. 2012, 63, 120–127. [Google Scholar] [CrossRef]
- Koprivica, M.R.; Trifković, J.; Dramićanin, A.M.; Gašić, U.M.; Akšić, M.M.F.; Milojković-Opsenica, D.M. Determination of the phenolic profile of peach (Prunus persica L.) kernels using UHPLC–LTQ OrbiTrap MS/MS technique. Eur. Food Res. Technol. 2018, 244, 2051–2064. [Google Scholar] [CrossRef]
- Kečkeš, S.; Gašić, U.; Veličković, T.Ć.; Milojković-Opsenica, D.; Natić, M.; Tešić, Ž. The determination of phenolic profiles of Serbian unifloral honeys using ultra-high-performance liquid chromatography/high resolution accurate mass spectrometry. Food Chem. 2013, 138, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, R.; Karar, M.G.E.; Gadir, H.A.; Kuhnert, N. Identification and characterisation of phenolics from Ixora coccinea L. (Rubiaceae) by liquid chromatography multi-stage mass spectrometry. Phytochem. Anal. 2014, 25, 567–576. [Google Scholar] [CrossRef]
- Karar, M.G.E.; Kuhnert, N. UPLC-ESI-Q-TOF-MS/MS Characterization of Phenolics from Crataegus monogyna and Crataegus laevigata (Hawthorn) Leaves, Fruits and their Herbal Derived Drops (Crataegutt Tropfen). J. Chem. Biol. Ther. 2016, 1, 102. [Google Scholar] [CrossRef]
- Silva, N.A.d.; Rodrigues, E.; Mercadante, A.Z.; de Rosso, V.V. Phenolic compounds and carotenoids from four fruits native from the Brazilian Atlantic forest. J. Agric. Food Chem. 2014, 62, 5072–5084. [Google Scholar] [CrossRef]
- Olate-Gallegos, C.; Barriga, A.; Vergara, C.; Fredes, C.; García, P.; Giménez, B.; Robert, P. Identification of Polyphenols from Chilean Brown Seaweeds Extracts by LC-DAD-ESI-MS/MS. J. Aquat. Food Prod. Technol. 2019, 28, 375–391. [Google Scholar] [CrossRef]
- Goufo, P.; Singh, R.K.; Cortez, I. A Reference List of Phenolic Compounds (Including Stilbenes) in Grapevine (Vitis vinifera L.) Roots, Woods, Canes, Stems, and Leaves. Antioxidants 2020, 9, 398. [Google Scholar] [CrossRef]
- Ablajan, K.; Abliz, Z.; Shang, X.Y.; He, J.M.; Zhang, R.P.; Shi, J.G. Structural characterization of flavonol 3,7-di-O-glycosides and determination of the glycosylation position by using negative ion electrospray ionization tandem mass spectrometry. J. Mass Spectrom. 2006, 41, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Price, W.E.; Ashton, J.; Tapsell, L.C.; Johnson, S. Identification and characterization of phenolic compounds in hydromethanolic extracts of sorghum wholegrains by LC-ESI-MSn. Food Chem. 2016, 211, 215–226. [Google Scholar] [CrossRef]
- Yasir, M.; Sultana, B.; Nigam, P.S.; Owusu-Apenten, R. Antioxidant and genoprotective activity of selected cucurbitaceae seed extracts and LC–ESIMS/MS identification of phenolic components. Food Chem. 2016, 199, 307–313. [Google Scholar] [CrossRef]
- De Beer, D.; Schulze, A.E.; Joubert, E.; De Villiers, A.; Malherbe, C.J.; Stander, M.A. Food Ingredient Extracts of Cyclopia subternata (Honeybush): Variation in Phenolic Composition and Antioxidant Capacity. Molecules 2012, 17, 14602–14624. [Google Scholar] [CrossRef]
- Lhuillier, A.; Fabre, N.; Moyano, F.; Martins, N.; Claparols, C.; Fourasté, I.; Moulis, C. Comparison of flavonoid profiles of Agauria salicifolia (Ericaceae) by liquid chromatography-UV diode array detection–electrospray ionisation mass spectrometry. J. Chromatogr. A 2007, 1160, 13–20. [Google Scholar] [CrossRef]
- Ojwang, L.O.; Yang, L.; Dykes, L.; Awika, J. Proanthocyanidin profile of cowpea (Vigna unguiculata) reveals catechin-O-glucoside as the dominant compound. Food Chem. 2013, 139, 35–43. [Google Scholar] [CrossRef]
- Maier, V.; Metzler, D.M.; Huber, A. 3-0-Caffeoylshikimic acid (dactylifric acid) and its isomers, a new class of enzymic browning substrates. Biochem. Biophys. Res. Commun. 1964, 14, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Zerbib, M.; Mazauric, J.-P.; Meudec, E.; Le Guernevé, C.; Lepak, A.; Nidetzky, B.; Cheynier, V.; Terrier, N.; Saucier, C. New flavanol O-glycosides in grape and wine. Food Chem. 2018, 266, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Avello, D.; Olmo-Cunillera, A.; Lozano-Castellón, J.; Marhuenda-Muñoz, M.; Vallverdú-Queralt, A. A Targeted Approach by High Resolution Mass Spectrometry to Reveal New Compounds in Raisins. Molecules 2020, 25, 1281. [Google Scholar] [CrossRef] [PubMed]
- García-Villalba, R.; Espín, J.C.; Tomás-Barberán, F.A.; Rocha-Guzmán, N.E. Comprehensive characterization by LC-DAD-MS/MS of the phenolic composition of seven Quercus leaf teas. J. Food Compos. Anal. 2017, 63, 38–46. [Google Scholar] [CrossRef]
- Li, D.; Zhao, J.; Xie, J.; Li, S. A novel sample preparation and on-line HPLC–DAD–MS/MS–BCD analysis for rapid screening and characterization of specific enzyme inhibitors in herbal extracts: Case study of α-glucosidase. J. Pharm. Biomed. Anal. 2014, 88, 130–135. [Google Scholar] [CrossRef]
- Bijttebier, S.; Van der Auwera, A.; Voorspoels, S.; Noten, B.; Hermans, N.; Pieters, L.; Apers, S. A First Step in the Quest for the Active Constituents in Filipendula ulmaria (Meadowsweet): Comprehensive Phytochemical Identification by Liquid Chromatography Coupled to Quadrupole-Orbitrap Mass Spectrometry. Planta Medica 2016, 82, 559–572. [Google Scholar] [CrossRef]
- Arapitsas, P.; Menichetti, S.; Vincieri, F.F.; Romani, A. Hydrolyzable Tannins with the Hexahydroxydiphenoyl Unit and the m-Depsidic Link: HPLC-DAD-MS Identification and Model Synthesis. J. Agric. Food Chem. 2007, 55, 48–55. [Google Scholar] [CrossRef]
- Arapitsas, P. Hydrolyzable tannin analysis in food. Food Chem. 2012, 135, 1708–1717. [Google Scholar] [CrossRef]
- Escobar-Avello, D.; Lozano-Castellón, J.; Mardones, C.; Pérez, A.J.; Saéz, V.; Riquelme, S.; von Baer, D.; Vallverdú-Queralt, A. Phenolic profile of grape canes: Novel compounds identified by LC-ESI-LTQ-Orbitrap-MS. Molecules 2019, 24, 3763. [Google Scholar]
- Abid, M.; Yaich, H.; Cheikhrouhou, S.; Khemakhem, I.; Bouaziz, M.; Attia, H.; Ayadi, M.A. Antioxidant properties and phenolic profile characterization by LC–MS/MS of selected Tunisian pomegranate peels. J. Food Sci. Technol. 2017, 54, 2890–2901. [Google Scholar] [CrossRef]
- Mata, A.; Ferreira, J.; Semedo, C.; Serra, T.; Duarte, C.; Bronze, M. Contribution to the characterization of Opuntia spp. juices by LC–DAD–ESI-MS/MS. Food Chem. 2016, 210, 558–565. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, C.E.V.; Stamford, T.L.M.; Neto, N.J.G.; de Souza, E.L. Inhibition of Staphylococcus aureus in broth and meat broth using synergies of phenolics and organic acids. Int. J. Food Microbiol. 2010, 137, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.-I.; Apostolidis, E.; Labbe, R.; Shetty, K. Inhibition of Staphylococcus aureus by phenolic phytochemicals of selected clonal herbs species of Lamiaceae family and likely mode of action through proline oxidation. Food Biotechnol. 2007, 21, 71–89. [Google Scholar] [CrossRef]
- Ma, Y.; Ding, S.; Fei, Y.; Liu, G.; Jang, H.; Fang, J. Antimicrobial activity of anthocyanins and catechins agianst foodborne pathogens Escherichia coli and Salmonella. Food Control 2019, 106, 106712. [Google Scholar] [CrossRef]
- Sorrentino, E.; Succi, M.; Tipaldi, L.; Pannella, G.; Maiuro, L.; Sturchio, M.; Coppola, R.; Tremonte, P. Antimicrobial activity of gallic acid against food-related Pseudomonas strains and its use as biocontrol tool to improve the shelf life of fresh black truffles. Int. J. Food Microbiol. 2018, 266, 183–189. [Google Scholar] [CrossRef]
- Lou, Z.; Wang, H.; Zhu, S.; Ma, C.; Wang, Z. Antibacterial Activity and Mechanism of Action of Chlorogenic Acid. J. Food Sci. 2011, 76, M398–M403. [Google Scholar] [CrossRef]
- Nile, S.H.; Park, S.W. Edible berries: Bioactive components and their effect on human health. Nutrition 2014, 30, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Lacombe, A.; Wu, V.C.; Tyler, S.; Edwards, K. Antimicrobial action of the American cranberry constituents; phenolics, anthocyanins, and organic acids, against Escherichia coli O157:H7. Int. J. Food Microbiol. 2010, 139, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Perez, C.; Wei, Y.; Rapoza, E.; Su, G.; Bou-Abdallah, F.; Chasteen, N. Iron-binding properties of plant phenolics and cranberry’s bio-effects. Dalton Trans. 2007, 4951–4961. [Google Scholar] [CrossRef] [PubMed]
- Stratford, M.; Nebe-Von-Caron, G.; Steels, H.; Novodvorska, M.; Ueckert, J.; Archer, D.B. Weak-acid preservatives: pH and proton movements in the yeast Saccharomyces cerevisiae. Int. J. Food Microbiol. 2013, 161, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Martínez, G.; Regente, M.; Jacobi, S.; Del Rio, M.; Pinedo, M.; de la Canal, L. Chlorogenic acid is a fungicide active against phytopathogenic fungi. Pestic. Biochem. Physiol. 2017, 140, 30–35. [Google Scholar] [CrossRef] [PubMed]


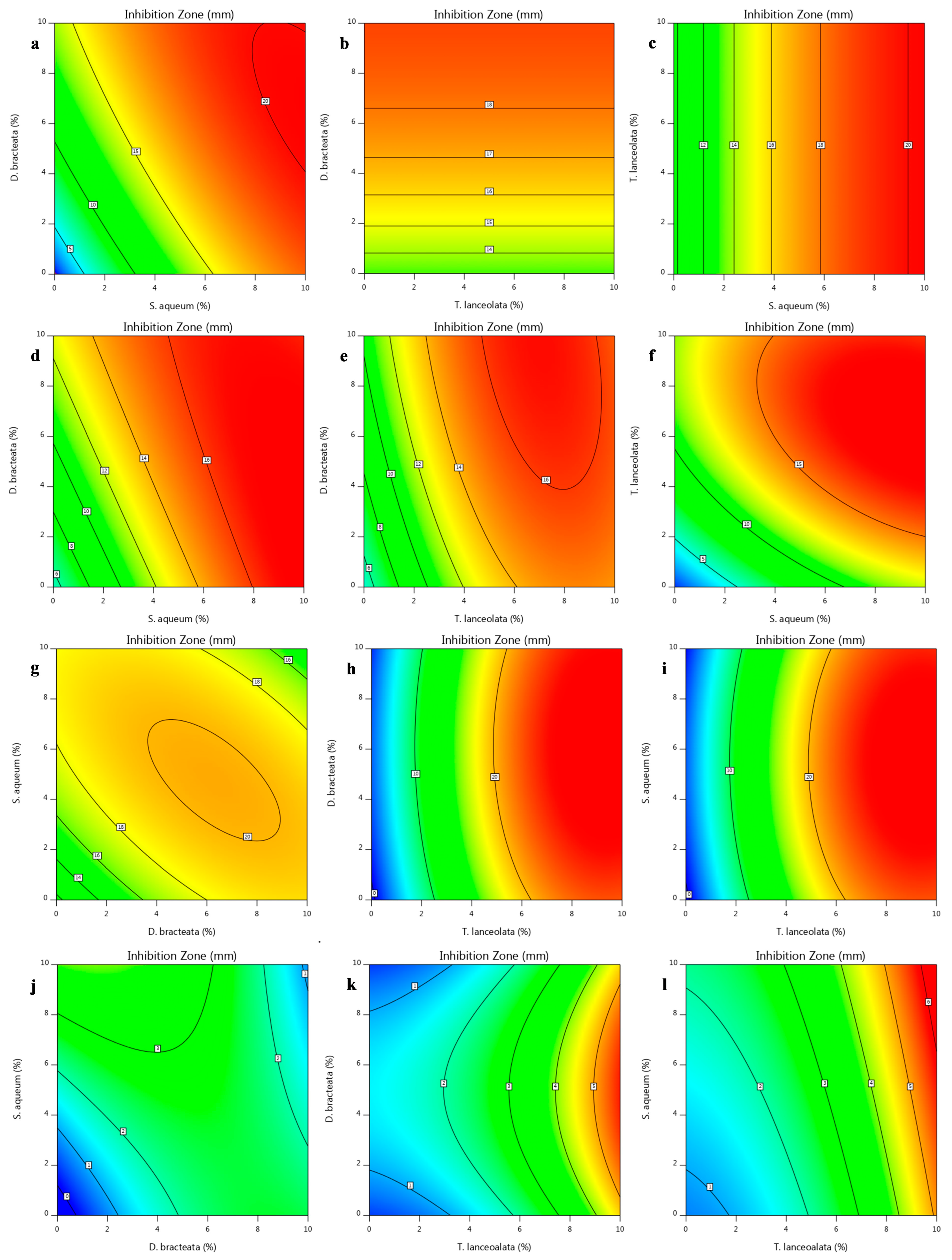
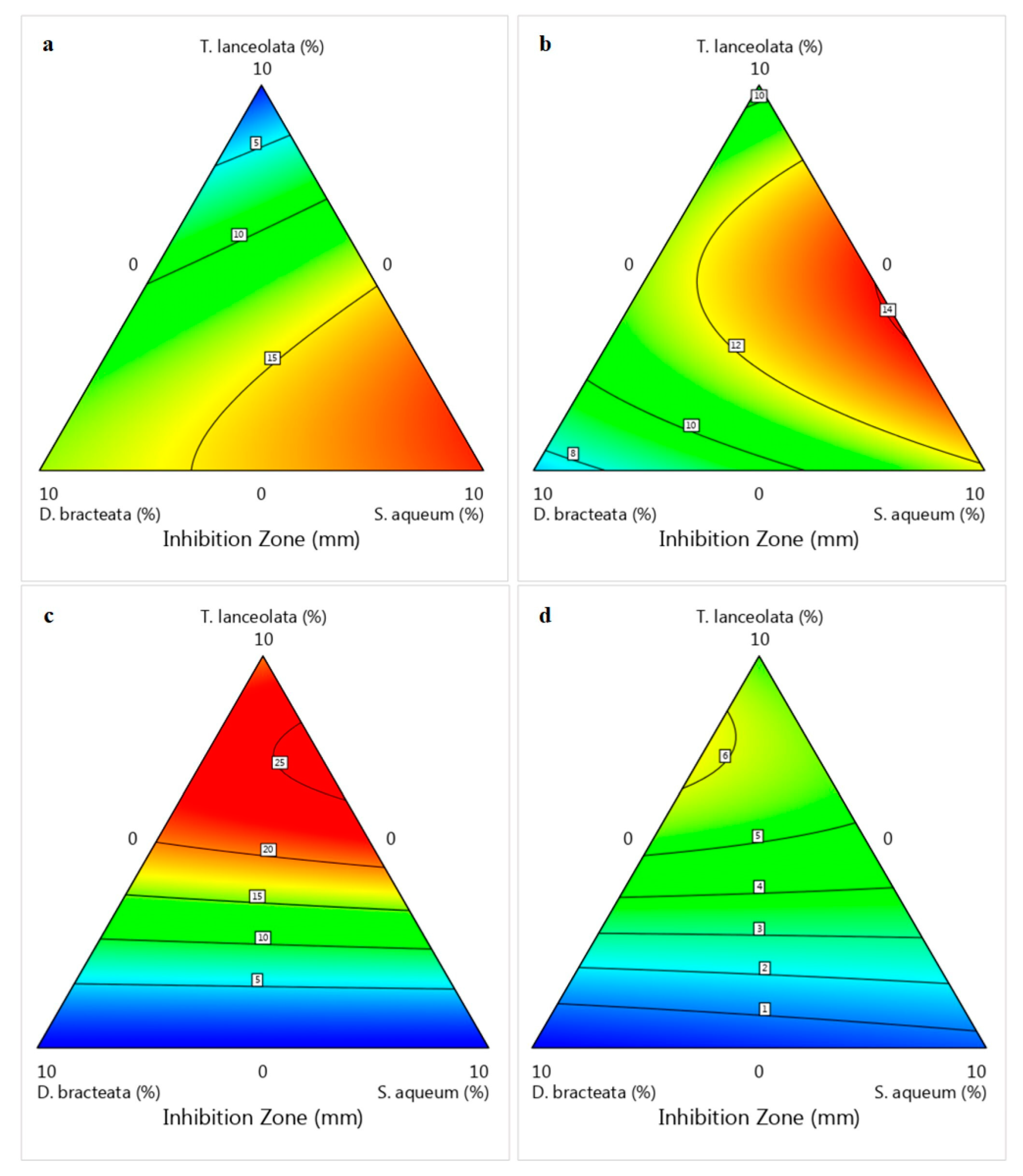
 ), Bacillus subtilis ((b);
), Bacillus subtilis ((b);  ), Rhodotorula diobovata ((c);
), Rhodotorula diobovata ((c);  ) and Alternaria alternata ((d);
) and Alternaria alternata ((d);  ) as a function of independent factors (Simplex–Lattice design).
), Bacillus subtilis ((b); ), Rhodotorula diobovata ((c); ) and Alternaria alternata ((d); ) as a function of independent factors (Simplex–Lattice design).
) as a function of independent factors (Simplex–Lattice design).
), Bacillus subtilis ((b); ), Rhodotorula diobovata ((c); ) and Alternaria alternata ((d); ) as a function of independent factors (Simplex–Lattice design).

{kind=link}
{kind=link}
| Analysis | Analytical Instrument | Electrospray Ionization | Multiple Reaction Monitoring (MRM) Transition * | Stationary Phase | Mobile Phase | Gradient Program | Reference |
|---|---|---|---|---|---|---|---|
| Sugars | Shimadzu Nexara X2 UHPLC, (Kyoto, Japan) equipped with a triple quadrupole mass spectrometer (MS-8045, Shimadzu). Data collection and processing were performed by Lab Solutions Insight LC-MC software (v.3.2, Shimadzu). | Negative mode. Nebulizer gas flow 3 L.min−1, drying gas flow of 10 L.min−1, desolvation line temperature of 250 °C, heat block temperature of 400 °C. | Fructose (179.2 → 113.1/89.0), Glucose (179.2 → 113.1/89.0), Sucrose (341.2 → 179.2/161.2/119.1) | Acquity UPLC BEH Amide column (100 × 2.1 mm i.d.; 1.7 µm particle size; Waters, Dublin, Ireland) at 40 °C. | Mobile phase A: 80% aqueous acetonitrile containing 0.1% NH4OH. Mobile phase B: 0.1% aqueous NH4OH. | 0% B, 1 min; linear increase to 40% B, 7 min; conditioning, 1 min; re-equilibration, 3 min. Flow rate of 0.2 mL.min−1. | [13] |
| Vitamin C | Waters UPLC-PDA system. Detection was carried out at 245 nm. | - | - | Waters HSS-T3 column (100 × 2.1 mm i.d.; 1.8 µm particle size) at 25 °C. | Mobile phase: 0.1% aqueous formic acid. | Flow rate of 0.3 mL.min−1. | [14] |
| Organic Acids | Shimadzu Nexera X2 UHPLC system, equipped with a MS-8045-TQ-mass spectrometer (0.2 µL injection). Data collection and processing were performed using Lab Solutions Insight LC-MS software (v.3.2, Shimadzu). | Negative mode. Nebulizer gas flow 3 L.min−1, drying gas flow of 10 L.min−1, desolvation line temperature of 250 °C, heat block temperature of 400 °C. | Succinic acid (117.00 → 73.00/99.10), Malic acid (133.00 → 114.95/70.95), Tartaric acid (149.00 → 87.00/72.85), Fumaric acid (115.00 → 71.05/26.95), Maleic acid (115.00 → 70.95/27.00), Isocitric acid (191.00 → 110.95/173.00), Citric acid (191.00 → 110.95/86.90), Shikimic acid (173.15 → 92.90/111.00), Quinic acid (191.00 → 84.95/92.90). | Waters HSS-T3 column (150 × 2.1 mm i.d.; 1.8 µm particle size) maintained at 40 °C. | Mobile phase A: 0.1% aqueous formic acid. Mobile phase B: 0.1% methanolic formic acid. | 0% B, 1 min; 5% B, 7 min; 50% B, 8 min; 50% B, 9 min; 0% B, 9.10 min; 0% B, 12 min. Flow rate of 0.2 mL.min−1. | [15] |
| Non-anthocyanin Phenolic Compounds | HRAM Thermo Q Exactive Orbitrap spectrometer (Thermo Fisher Scientific, Melbourne, VIC, Australia) equipped with an Ultimate Dionex 3000 RS UHPLC-UV/Vis detector (Thermo Fisher Scientific). UHPLC-UV chromatograms were recorded at 280 and 360 nm. Data processing was performed with Xcalibur software (v.4.1, Thermo). | Negative mode. Collision energy of 25 V, resolving power of 70,000 FWHM, 3 × 106 AGC (automatic gain control) target, injection time of 200 ms (for peak identification). | - | Waters HSS-T3 column (150 × 2.1 mm i.d.; 1.8 µm) at 40 °C. | Mobile phase A: 0.1% aqueous formic acid, Mobile phase B: 0.1% formic acid in acetonitrile. | 5% B, 1 min; 20% B, 5 min; 20% B, 7 min; 45% B, 12 min; 100% B, 19 min; 100% B, 22 min; 5% B, 22.1 min; 5% B, 26 min. Flow rate of 0.3 mL.min−1. | - |
| Run | A/Component1 (TL, %) | B/Component2 (DB, %) | C/Component3 (SA, %) | Pseudomonas viridiflava | Bacillus subtilis | Rhodotorula diobovata | Alternaria alternata |
|---|---|---|---|---|---|---|---|
| Box–Behnken | |||||||
| 1 | 0 | 10 | 5 | 19.06 | 10.51 | 0.00 | 0.00 |
| 2 | 0 | 0 | 5 | 13.20 | 5.63 | 0.00 | 0.00 |
| 3 | 5 | 10 | 0 | 14.37 | 13.05 | 19.29 | 2.34 |
| 4 | 10 | 10 | 5 | 18.40 | 14.96 | 22.48 | 5.96 |
| 5 | 0 | 5 | 10 | 20.33 | 12.14 | 0.00 | 3.57 |
| 6 | 10 | 5 | 10 | 20.13 | 16.98 | 21.39 | 5.46 |
| 7 | 5 | 5 | 5 | 17.75 | 15.25 | 19.56 | 3.52 |
| 8 | 5 | 0 | 0 | 0.00 | 5.76 | 9.17 | 0.00 |
| 9 | 5 | 5 | 5 | 17.25 | 15.69 | 19.68 | 2.78 |
| 10 | 5 | 0 | 10 | 18.19 | 16.66 | 18.02 | 3.65 |
| 11 | 10 | 0 | 5 | 13.29 | 14.24 | 24.33 | 4.73 |
| 12 | 0 | 5 | 0 | 9.02 | 0.00 | 0.00 | 0.00 |
| 13 | 10 | 5 | 0 | 9.84 | 11.42 | 23.86 | 4.82 |
| 14 | 5 | 5 | 5 | 17.16 | 15.89 | 20.78 | 2.47 |
| 15 | 5 | 5 | 5 | 17.08 | 15.07 | 20.82 | 3.04 |
| 16 | 5 | 5 | 5 | 17.08 | 14.20 | 20.29 | 2.51 |
| 17 | 5 | 10 | 10 | 20.00 | 15.99 | 16.88 | 0.00 |
| Simplex–Lattice Mixture | |||||||
| 1 | 5.00 | 5.00 | 0.00 | 10.05 | 10.04 | 19.05 | 3.71 |
| 2 | 0.00 | 0.00 | 10.00 | 19.52 | 10.87 | 0.00 | 0.00 |
| 3 | 6.67 | 1.67 | 1.67 | 11.14 | 12.37 | 22.74 | 8.29 |
| 4 | 5.00 | 5.00 | 0.00 | 9.15 | 12.29 | 18.42 | 5.24 |
| 5 | 1.67 | 6.67 | 1.67 | 13.58 | 12.55 | 5.18 | 1.76 |
| 6 | 1.67 | 1.67 | 6.67 | 16.48 | 12.86 | 5.05 | 1.69 |
| 7 | 0.00 | 10.00 | 0.00 | 14.17 | 7.58 | 0.00 | 0.00 |
| 8 | 0.00 | 5.00 | 5.00 | 16.36 | 10.42 | 0.00 | 0.00 |
| 9 | 3.33 | 3.33 | 3.33 | 13.60 | 10.99 | 16.29 | 3.28 |
| 10 | 10.00 | 0.00 | 0.00 | 0.00 | 9.26 | 19.50 | 4.47 |
| 11 | 10.00 | 0.00 | 0.00 | 0.00 | 9.80 | 21.26 | 5.34 |
| 12 | 0.00 | 0.00 | 10.00 | 18.15 | 12.00 | 0.00 | 1.76 |
| 13 | 5.00 | 0.00 | 5.00 | 14.39 | 14.13 | 20.93 | 3.67 |
| 14 | 0.00 | 10.00 | 0.00 | 12.75 | 5.58 | 0.00 | 0.00 |
| Positive/Negative Controls | |||||||
| Streptomycin (20 µg.mL−1) | 17.16 ± 0.05 | 20.22 ± 0.05 | - | - | |||
| Voriconazole (200 µg.mL−1) | - | - | 27.82 ± 0.30 | 41.14 ± 0.64 | |||
| Sterile water | 0.00 | 0.00 | 0.00 | 0.00 | |||
| T. lanceolata | D. bracteata | S. aqueum | ||
|---|---|---|---|---|
| Sugars (g. 100 g−1 dw) | Fructose | 2.08 ± 0.07 c | 17.15 ± 0.51 a | 10.52 ± 0.44 b |
| Glucose | 1.79 ± 0.06 c | 15.12 ± 0.64 a | 6.33 ± 0.25 b | |
| Sucrose | 5.69 ± 0.08 a | 1.41 ± 0.07 b | 0.02 ± 0.00 c | |
| Vitamin C (mg. 100 g−1 dw) | L-AA | 0.62 ± 0.01 b | 1.75 ± 0.02 a | 0.59 ± 0.01 b |
| Total AA | 0.89 ± 0.02 c | 2.43 ± 0.03 a | 1.20 ± 0.10 b | |
| Organic acids (g. 100 g−1 dw) | Citric acid | 1.43 ± 0.11 b | 1.75 ± 0.07 b | 28.81 ± 0.34 a |
| Fumaric acid | 0.00 ± 0.00 b | 0.01 ± 0.00 a | 0.01 ± 0.00 a | |
| Isocitric acid | 0.36 ± 0.03 b | 0.37 ± 0.02 b | 0.49 ± 0.02 a | |
| Malic acid | 2.44 ± 0.21 c | 22.19 ± 0.11 a | 15.95 ± 0.43 b | |
| Quinic acid | 1.50 ± 0.11 b | 1.92 ± 0.08 b | 29.39 ± 0.28 a | |
| Shikimic acid | 5.30 ± 0.30 a | 0.01 ± 0.00 b | 0.09 ± 0.00 b | |
| Succinic acid | 0.02 ± 0.00 c | 0.10 ± 0.00 a | 0.06 ± 0.00 b | |
| TPC (mg GAE.g−1 dw) | 123.47 ± 1.29 a | 6.07 ± 0.77 b | 6.45 ± 0.19 b | |
| DPPH IC50 (μg.mL−1) | 36.59 ± 0.41 c | 353.60 ± 9.23 a | 299.89 ± 3.11 b |
| Compound No. | RT (min) | [M-H]− (m/z) | Molecular Formula | ΔM (ppm) | MS2 Fragmentation (m/z) | Tentative Identification |
|---|---|---|---|---|---|---|
| Phenolic Acids | ||||||
| 1 | 2.75 | 371.0984 | C16H20O10 | 0.0808 | 371.0959; 191.0557; 135.0444; 85.0284 | Hydroxydihydrocaffeoylquinic acid |
| 3 | 3.87 | 315.0714 | C13H16O9 | −2.3994 | 315.0708; 108.0209; 152.0109; 207.9458 | Protocatechuic acid O-hexoside |
| 4 | 5.19 | 153.0189 | C7H6O4 | −2.8231 | 153.0189; 109.0286; 123.0445; 91.0173 | Protocatechuic acid |
| 6 | 6.40 | 371.0979 | - | - | 119.0496; 163.0394; 359.1305 | p-coumaric acid derivative |
| 8 | 6.64 | 707.1789 | - | - | 707.1731; 191.0559; 243.0657; 173.0452; 323.0540; 463.1036; 515.1109 | Unknown, perhaps caffeoylquinic acid glucoside derivative |
| 10 | 6.87 | 447.1867 | - | - | 153.0916; 137.0238; 271.0969; 359.0724 | Unknown, perhaps hydroxybenzoic acid derivative |
| 12 | 7.30 | 353.0864 | C16H18O9 | −3.9820 | 353.0857; 191.0556; 85.0284; 127.0392 | Chlorogenic acid (syn: 5-caffeoylquinic acid) ** |
| 13 | 7.36 | 707.1799 | C32H36O18 | −4.2252 | 191.0556; 85.0284; 353.0851 | Chlorogenic acid dimer |
| 14 | 8.13 | 707.1787 | C32H36O18 | −5.9220 | 191.0556; 85.0284; 353.0832; 593.1383 | Chlorogenic acid dimer isomer |
| 16 | 8.40 | 337.0920 | C16H18O8 | −2.6431 | 337.0886; 93.0336; 119.0495; 173.0450; 163.0393; 87.0077; 255.1010 | 4-O-p-coumaroylquinic acid |
| Flavonoids and derivatives | ||||||
| 15 | 8.27 | 461.1658 | - | - | 101.0235; 113.0236; 289.0708; 153.0913; 161.0447; 329.1339 | Unknown, perhaps tricin derivative |
| 26 | 11.47 | 417.0824 | - | - | 417.0818; 284.0324; 315.0487; 133.0288 | Unknown, luteolin derivative |
| 27 | 11.47 | 547.1651 | - | - | 285.0395; 284.0325; 192.0422; 89.0233; 493.1665 | Unknown, luteolin derivative |
| 32 | 12.97 | 331.1208 | - | - | 331.1208; 96.9592; 219.1385; 263.1286; 269.0450 | Unknown, perhaps apigenin derivative |
| 33 | 13.07 | 505.2056 | - | - | 343.1535; 328.1302; 251.1653; 427.1900 | Unknown, perhaps luteolin-trimethyl ester-O-hexoside |
| 34 | 13.88 | 301.0349 | C15H10O7 | −1.5812 | 301.0337; 133.0290; 151.0033; 121.0290; 83.0128 | Quercetin ** |
| 35 | 14.11 | 483.2434 | - | - | 299.0552; 284.0319; 209.0805; 165.0910 | Unknown, perhaps diosmetin derivative |
| 36 | 14.23 | 459.2218 | - | - | 96.9592; 331.1207; 299.0551; 284.0317 | Unknown, perhaps diosmetin derivative |
| 37 | 14.94 | 269.0449 | C15H10O5 | −2.4047 | 269.0453; 117.0339; 83.0128; 151.0030 | Apigenin |
| 38 | 16.27 | 299.0550 | C16H12O6 | −3.7183 | 299.0552; 284.0321; 133.0289; 203.1437; 107.0131; 168.9887; 256.0364; 265.1436; 83.0128 | Diosmetin |
| 39 | 17.29 | 283.0603 | C16H12O5 | −3.1689 | 283.0602; 117.0337; 268.0371; 237.1491; 211.0395; 107.0130; 151.0030; 239.0344; 191.1434; 83.0127 | Apigenin 7,4′-dimethyl ether (syn: Genkwanin, Acacetin) |
| Flavonoid glycosides | ||||||
| 17 | 8.57 | 435.2226 | - | - | 289.0709; 177.0189; 339.0463; 245.0815 | Unknown, perhaps catechin rhamnoside |
| 21 | 9.94 | 609.1442 | C27H30O16 | −3.1322 | 609.1413; 300.0275; 447.0931; 151.0030 | Rutin ** |
| 22 | 10.57 | 463.0879 | C21H20O12 | −0.6478 | 463.0865; 271.0243; 300.0266; 255.0293; 151.0029; 243.0294; 178.9978 | Quercetin-3-O-glucoside ** |
| 23 | 11.05 | 863.1996 | - | - | 431.0974; 283.0605; 311.0549; 96.9591; 151.0030; 345.0993; 131.0495 | Unknown, perhaps vitexin/isovitexin dimer |
| 24 | 11.05 | 431.0981 | C21H20O10 | −0.6263 | 431.0975; 283.0606; 311.0549; 96.9591; 151.0030; 345.0996 | Vitexin/isovitexin |
| 25 | 11.25 | 593.1490 | C27H30O15 | −3.6988 | 593.1443; 285.0392; 255.0295; 361.1616; 165.0547; 523.2097 | Kaempferol glycoside (perhaps Kaempferol O-hexosyl-deoxyhexose) |
| 28 | 11.54 | 563.1383 | C26H28O14 | −4.1357 | 563.1362; 285.0397; 192.0423; 89.0232 | Kaempferol glycoside (perhaps Kaempferol 3-O-rhamnoside-7-O-xyloside) |
| 30 | 12.33 | 447.0929 | C21H20O11 | −0.8611 | 447.0861; 285.0402 | Luteolin glycoside (perhaps Luteolin 8-C-glucoside) |
| 31 | 12.43 | 593.1477 | C27H30O15 | −5.8905 | 269.0451; 547.1425 | Apigenin dihexoside |
| Biflavonoids & polyflavonoids | ||||||
| 2 | 3.37 | 865.1781 | - | - | 140.0110; 287.0547; 407.0700; 543.0834; 451.0955 | Perhaps procyanidin trimer (B-type) |
| 9 | 6.77 | 577.1331 | C30H26O12 | −3.5520 | 577.1282; 125.0237; 289.0710; 161.0239; 245.0814; 407.0778; 205.0498 | (epi)catechin-(epi)catechin OR procyanidin dimer (B type) |
| 18 | 8.66 | 739.1623 | C39H32O15 | −6.1474 | 739.1487; 289.0714; 177.0191; 339.0488; 245.0814; 459.0654; 587.1086; 117.0551 | Procyanidin dimer monoglycoside |
| 19 | 8.75 | 577.1325 | C30H26O12 | −4.5916 | 125.0239; 289.0712; 245.0814; 491.01842; 203.0705 | Procyanidin dimer (B type) |
| 20 | 8.99 | 739.1622 | C39H32O15 | −6.2827 | 739.1502; 289.0712; 177.0190; 339.0489; 245.0813; 459.0662; 137.0239; 569.0995 | Procyanidin dimer monoglycoside |
| Other polyphenols | ||||||
| 5 | 6.27 | 356.0976 | - | - | 121.0289; 237.403; 149.0603; 219.0293; 293.0631 | Unknown, perhaps hydroxybenzaldehyde derivative |
| 7 | 6.56 | 371.1336 | - | - | 243.0657; 323.0540; 289.0556; 173.0454 | Unknown, perhaps piceatannol derivative |
| 11 | 7.05 | 401.1434 | - | - | 401.1385; 96.9592; 361.0948; 134.0367; 239.0919; 271.0964 | Unknown, perhaps pelargonidin-3-pentoside derivative |
| 29 | 11.67 | 451.1031 | C24H20O9 | −0.7891 | 451.1018; 189.0189; 217.0136; 341.0650; 177.0188; 109.0287; 123.0444; 151.0395; 255.0294; 402.1244; 447.0855 | Cinochonain l |
| Compound No. | RT (min) | [M-H]− (m/z) | Molecular Formula | ΔM (ppm) | MS2 Fragmentation (m/z) | Tentative Identification |
|---|---|---|---|---|---|---|
| Organic acids | ||||||
| 1 | 1.39 | 133.0136 | C4H6O5 | −4.8641 | 133.0136; 115.0029; 89.0230; 111.0195; 124.0143 | Malic acid |
| 2 | 1.96 | 117.0185 | - | - | 117.0185; 100.0394 | Unknown, perhaps succinic acid |
| Phenolic acids | ||||||
| 14 | 9.96 | 319.0790 | - | - | 119.0496 | Unknown, perhaps coumaric acid derivative |
| 16 | 11.14 | 355.1027 | - | - | 147.0445; 168.9885; 216.9797; 273.9771; 114.9481 | Unknown, perhaps cinnamic acid derivative |
| 17 | 11.80 | 415.1964 | - | - | 341.0607; 161.0609 | Unknown, perhaps caffeoyl glucose derivative |
| 20 | 12.51 | 631.2519 | - | - | 245.1545; 201.1647; 523.1197 | Unknown, perhaps heptyl cinnamate derivative |
| 22 | 14.62 | 509.2578 | - | - | 101.0235; 85.0284; 113.0239; 231.0989 | Unknown, perhaps tetrahydrofurfuryl cinnamate derivative |
| Flavonoids and derivatives | ||||||
| 8 | 7.38 | 289.0713 | C15H14O6 | −1.5982 | 289.0710; 109.0287; 123.0444; 191.0556; 97.0286 | Catechin ** |
| 11 | 8.15 | 417.1319 | - | - | 125.0239; 177.0190; 151.0396; 287.0552; 243.0294 | Unknown, perhaps eriodictyol derivative |
| 12 | 8.27 | 289.0707 | C15H14O6 | −3.6738 | 289.0710; 109.0286; 123.0444; 245.0814; 83.0127 | Epicatechin ** |
| 15 | 10.91 | 495.1125 | - | - | 151.0034; 285.0397; 125.0236; 107.0130; 178.9980; 83.0128; 340.9699; 303.0478; 449.1015 | Unknown, perhaps luteolin derivative |
| 21 | 12.67 | 445.2071 | - | - | 445.1998; 165.1280; 209.1180; 283.1545; 337.0700; 87.0441 | Unknown, perhaps wogonin derivative |
| Flavonoid glycosides | ||||||
| 9 | 7.82 | 447.1497 | - | - | 125.0236; 289.0712; 161.0238; 407.0796 | Unknown, perhaps catechin glycoside |
| 18 | 11.90 | 477.0980 | - | - | 477.0980; 299.0194; 119.0496; 314.0470 | Perhaps isorhamnetin-3-O-hexoside |
| Biflavonoids and polyflavonoids | ||||||
| 5 | 6.30 | 593.1262 | C30H26O13 | −6.5145 | 177.0191; 339.0832; 273.0395 | Prodelphinidin A-type |
| 6 | 6.77 | 577.1323 | C30H26O12 | −4.9381 | 577.1257; 125.0238; 289.0714; 161.0240; 245.0814; 407.0787 | Procyanidin dimer B-type |
| 7 | 7.15 | 577.1317 | C30H26O12 | −5.9778 | 161.0241; 289.0709; 407.0772; 339.0813 | Procyanidin dimer B-type |
| 10 | 7.82 | 577.1317 | C30H26O12 | −5.9778 | 577.1245; 125.0236; 289.0712; 407.0786; 161.0238; 245.0813; 137.0237; 339.0815; 425.0938 | Procyanidin dimer B-type |
| 13 | 8.43 | 865.1772 | - | - | 165.0916; 287.0553; 543.0829; 577.1254 | Perhaps procyanidin trimer/epicatechin-epicatechin-epicatechin |
| Other polyphenols | ||||||
| 3 | 3.16 | 344.1282 | - | - | 147.0441; 164.0703 | Unknown, perhaps coumarin derivative |
| 4 | 3.89 | 154.0505 | - | - | 82.0288; 108.0213 | Unknown, perhaps hypogallic acid |
| 19 | 12.43 | 489.2277 | - | - | 269.0455; 167.0343; 331.1887 | Unknown, perhaps carnosic acid derivative |
| Compound No. | RT (min) | [M-H]− (m/z) | Molecular Formula | ΔM (ppm) | MS2 Fragmentation (m/z) | Tentative Identification |
|---|---|---|---|---|---|---|
| Organic acid | ||||||
| 1 | 1.46 | 133.0135 | C4H6O5 | −5.6159 | 133.0130; 111.0078; 115.0028; 96.9590; 107.2179 | Malic acid |
| 2 | 1.61 | 191.0189 | C6H8O7 | −4.3241 | 111.0078; 87.0076; 155.9504; 170.0026 | Citric acid |
| Phenolic acids | ||||||
| 3 | 2.52 | 169.0132 | C7H6O5 | −6.1947 | 125.0236 | Gallic acid ** |
| 5 | 6.87 | 327.0709 | C14H16O9 | −3.8401 | 312.045; 206.0210; 207.0290; 193.0130; 205.0137; 234.0162; 327.0663; 192.0060; 164.0109; 136.0159 | Bergenin |
| 8 | 7.53 | 759.1153 | C37H28O18 | −6.5707 | 759.1153; 175.0032; 289.0333; 301.0338; 423.0757; 345.0189; 481.0681; 468.0614 | Theacitrin A |
| Flavonoids and derivatives | ||||||
| 7 | 7.30 | 511.1069 | - | - | 447.0876; 284.0318; 166.0265; 109.0288; 329.0816 | Unknown, perhaps kaempferol derivative |
| 11 | 8.12 | 305.0691 | - | - | 96.9592; 125.0238; 169.0137; 177.0188; 243.0294; 305.0660 | Unknown, perhaps gallocatechin gallate |
| 26 | 12.39 | 317.0290 | C15H10O8 | −4.0721 | 301.0347; 151.0031; 109.0287; 137.0238; 178.9981; 227.343 | Myricetin ** |
| 27 | 13.88 | 301.0342 | C15H10O7 | −3.9065 | 151.0031; 107.0130; 121.0288; 93.0036; 83.0128 | Quercetin ** |
| Flavonoid glycosides | ||||||
| 6 | 7.30 | 447.0927 | C21H20O11 | −1.3084 | 447.0082; 285.0385; 241.0499; 147.0081; 199.0394; 329.0816 | Luteolin-3-glucoside |
| 14 | 9.00 | 479.0811 | C21H20O13 | −4.2038 | 479.0776; 316.0215; 271.0242; 287.0191; 372.9598; 214.0260 | Myricetin-3-O-β-D-galactopyranoside isomer |
| 15 | 9.11 | 479.0807 | C21H20O13 | −5.0387 | 479.0750; 316.0220; 271.0250; 287.0190; 109.0290 | Myricetin-3-O-β-D-galactopyranoside |
| 17 | 10.03 | 449.0719 | - | - | 449.0633; 316.0214; 271.0246; 287.0185; 283.0604; 242.0224; 405.9114; 214.0263 | Kamepferol derivative; perhaps dihydrokaempferol-hexoside |
| 18 | 10.20 | 431.0970 | C21H20O10 | −3.1779 | 283.0605; 311.0553; 341.0697; 323.0517; 239.0716; 211.0756 | Vitexin or isovitexin |
| 20 | 10.41 | 597.1743 | - | - | 597.1736; 357.0947; 387.1089; 209.0447; 239.0576; 417.1104 | Perhaps phloretin-di-glucoside |
| 21 | 10.55 | 463.0862 | C21H20O12 | −4.3188 | 463.0862; 300.0268; 271.0245; 255.0295; 151.0031; 356.9632; 390.9250 | Quercetin-3-O-glucoside ** |
| 22 | 10.83 | 479.0798 | C21H20O13 | −6.9173 | 479.0728; 178.9982; 317.0288; 406.9325 | Myricetin-glycoside |
| 23 | 11.10 | 433.0806 | C20H18O11 | 6.8463 | 433.0674; 300.0269; 301.0339; 271.0248; 255.0287; 243.0299; 390.9264; 356.9643 | Quercetin-glycoside |
| 24 | 11.33 | 435.1340 | C21H24O10 | 9.9487 | 125.0240; 167.0340; 273.0750; 315.0840; 369.0010 | Phloridzin |
| 25 | 11.53 | 433.0766 | C20H18O11 | −2.3898 | 433.0691; 271.0246; 300.0246; 315.0123; 163.0029; 299.9917 | Quercetin-glycoside |
| Tannins | ||||||
| 4 | 3.67 | 933.0374 | - | - | 933.0371; 300.9981; 275.0193; 125.0238; 229.0140; 314.0032; 421.0121; 467.0170; 492.9950; 569.0468; 613.0347; 871.0412 | Perhaps castalagin |
| 9 | 7.61 | 1139.3467 | - | - | 177.0189; 125.0237; 169.0139; 243.0297; 759.1021; 633.0806 | Unknown, perhaps galloylated tannin |
| 10 | 7.68 | 953.0593 | - | - | 953.0590; 125.0240; 177.0190; 169.0140; 165.0190; 137.0240; 151.0400; 243.0300; 275.0180; 301.0320; 299.0160; 423.0650; 759.1010; 935.0510; 633.0800 | Perhaps chebulagic acid |
| 12 | 8.52 | 911.1115 | - | - | 911.1061; 125.0239; 169.0137; 96.9592; 177.0188; 137.0238; 285.0399; 571.0787; 741.0919; 423.0636; 615.0666; 305.0635 | Perhaps theasinesin A |
| 13 | 8.66 | 935.0502 | - | - | 935.0495; 300.9984; 125.0238; 169.0138; 275.0186; 633.0611 | Perhaps casuarinin |
| 16 | 9.43 | 895.1121 | - | - | 895.1100; 299.9907; 447.0488; 361.1577; 300.9964; 555.0822 | Unknown, ellagic acid derivative |
| 19 | 10.33 | 300.9978 | C14H6O8 | −3.9568 | 300.9978; 145.0287; 169.0133; 117.0338; 245.0082; 283.9944; 228.0052; 200.0106 | Ellagic acid ** |
| Parameters | Box–Behnken Design | Simplex–Lattice Design | |||||||
|---|---|---|---|---|---|---|---|---|---|
| PV | BS | RD | AA | PV | BS | RD | AA | ||
| Model | df | 6 | 9 | 7 | 6 | 5 | 4 | 4 | 4 |
| F-value | 428.33 | 53.74 | 59.88 | 13.06 | 85.69 | 7.43 | 431.83 | 19.38 | |
| p-value | <0.0001 | <0.0001 | <0.0001 | <0.0003 | <0.0001 | <0.0063 | <0.0001 | <0.0002 | |
| Residual | df | 10 | 7 | 9 | 10 | 8 | 9 | 9 | 9 |
| msq | 1.55 | 0.7169 | 3.19 | 0.7455 | 1.03 | 1.71 | 0.0246 | 0.0836 | |
| ssq | 15.54 | 5.02 | 28.70 | 7.45 | 8.25 | 15.39 | 0.2212 | 0.7527 | |
| Lack of fit | df | 6 | 3 | 5 | 6 | 4 | 5 | 5 | 5 |
| F-value | 1.81 | 2.51 | 15.49 | 5.95 | 2.51 | 1.51 | 7.63 | 0.7315 | |
| p-value | 0.2937 | 0.1980 | 0.0100 | 0.0531 | 0.1971 | 0.3558 | 0.0357 | 0.6365 | |
| Pure error | df | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 |
| msq | 1.04 | 0.4357 | 0.3524 | 0.1879 | 0.5872 | 1.33 | 0.0052 | 0.0983 | |
| ssq | 4.18 | 1.74 | 1.41 | 0.7516 | 2.35 | 5.34 | 0.0210 | 0.3932 | |
| R2 | 0.9961 | 0.9857 | 0.9790 | 0.8869 | 0.9817 | 0.7675 | 0.9948 | 0.8960 | |
| Ra2 | 0.9938 | 0.9674 | 0.9626 | 0.8190 | 0.9702 | 0.6641 | 0.9925 | 0.8497 | |
| Response | Predicted Mean Value | Experimental Value | −95% Prediction | +95% Prediction |
|---|---|---|---|---|
| Box–Behnken | ||||
| Pseudomonas viridiflava | 17.21 | 16.78 | 16.78 | 17.64 |
| Bacillus subtilis | 15.90 | 14.54 | 14.52 | 17.29 |
| Rhodotorula diobovata | 25.29 | 26.34 | 22.49 | 28.09 |
| Alternaria alternata | 5.29 | 6.31 | 4.03 | 6.55 |
| Simplex–Lattice | ||||
| Pseudomonas viridiflava | 15.12 | 14.53 | 12.93 | 17.32 |
| Bacillus subtilis | 14.03 | 12.95 | 11.33 | 16.72 |
| Rhodotorula diobovata | 20.41 | 22.84 | 17.53 | 23.44 |
| Alternaria alternata | 4.42 | 6.96 | 2.07 | 7.31 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seididamyeh, M.; Phan, A.D.T.; Sivakumar, D.; Netzel, M.E.; Mereddy, R.; Sultanbawa, Y. Valorisation of Three Underutilised Native Australian Plants: Phenolic and Organic Acid Profiles and In Vitro Antimicrobial Activity. Foods 2023, 12, 623. https://doi.org/10.3390/foods12030623
Seididamyeh M, Phan ADT, Sivakumar D, Netzel ME, Mereddy R, Sultanbawa Y. Valorisation of Three Underutilised Native Australian Plants: Phenolic and Organic Acid Profiles and In Vitro Antimicrobial Activity. Foods. 2023; 12(3):623. https://doi.org/10.3390/foods12030623
Chicago/Turabian StyleSeididamyeh, Maral, Anh Dao Thi Phan, Dharini Sivakumar, Michael E. Netzel, Ram Mereddy, and Yasmina Sultanbawa. 2023. "Valorisation of Three Underutilised Native Australian Plants: Phenolic and Organic Acid Profiles and In Vitro Antimicrobial Activity" Foods 12, no. 3: 623. https://doi.org/10.3390/foods12030623
APA StyleSeididamyeh, M., Phan, A. D. T., Sivakumar, D., Netzel, M. E., Mereddy, R., & Sultanbawa, Y. (2023). Valorisation of Three Underutilised Native Australian Plants: Phenolic and Organic Acid Profiles and In Vitro Antimicrobial Activity. Foods, 12(3), 623. https://doi.org/10.3390/foods12030623