
New Insights into the Inhibition of Hesperetin on Polyphenol Oxidase: Inhibitory Kinetics, Binding Characteristics, Conformational Change and Computational Simulation


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. PPO Activity Assay
2.2.2. Kinetic Analysis of Inhibitory Type
2.2.3. Multivariate Curve Resolution–Alternate Least Squares (MCR–ALS) Algorithm
2.2.4. Fluorometric Titration Experiments
2.2.5. Synchronous Fluorescence Experiments
2.2.6. Three-Dimensional (3D) Fluorescence Experiments
2.2.7. CD Spectra
2.2.8. Molecular Docking
2.2.9. Molecular Dynamics (MD) Simulation
2.3. Statistical Analysis
3. Results
3.1. Inhibitory Activity of Hesperetin
3.2. Inhibition Kinetics of Hesperetin
3.3. Fluorescence Spectra of PPO–Hesperetin Interaction
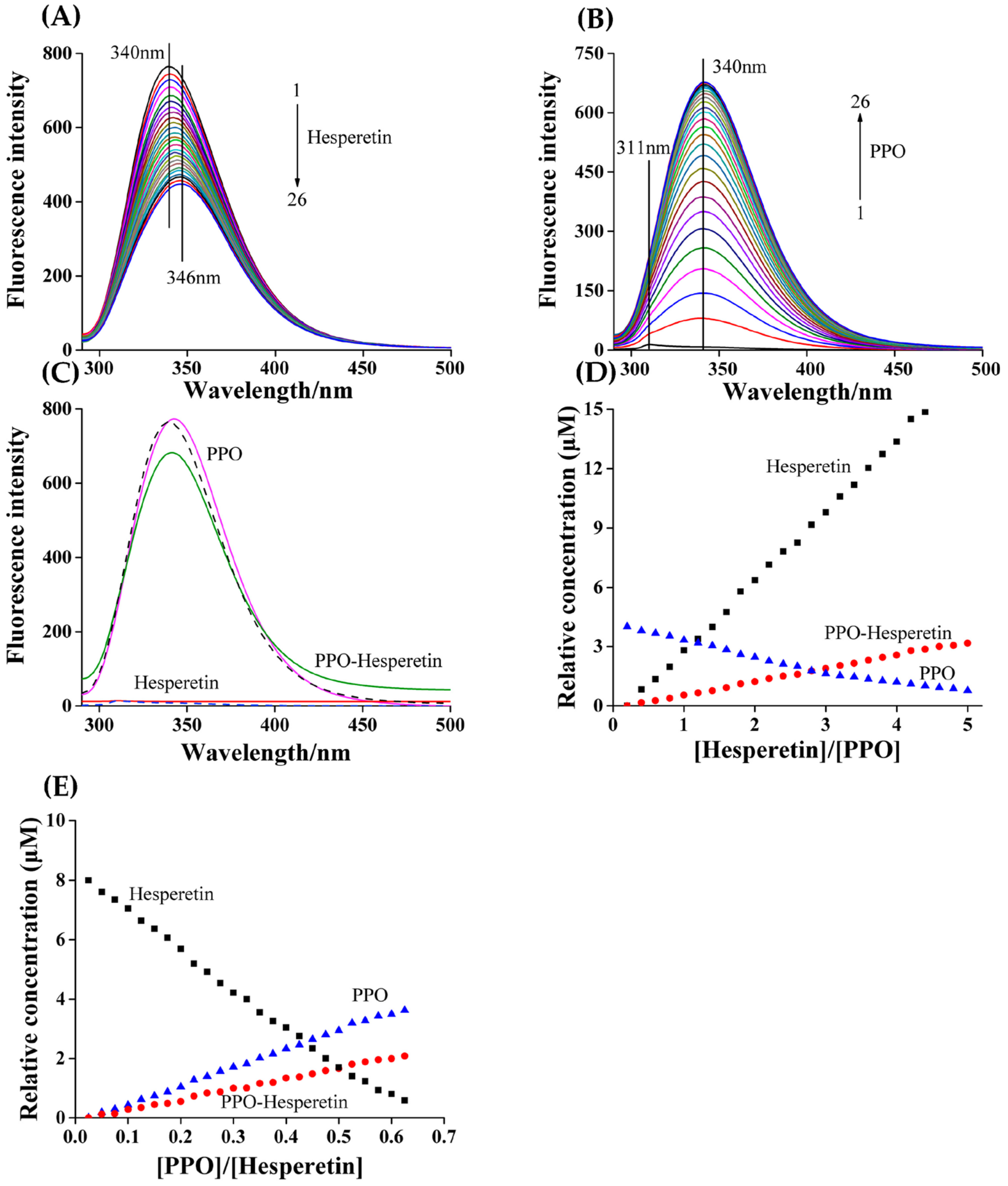
3.4. MCR–ALS Decomposition
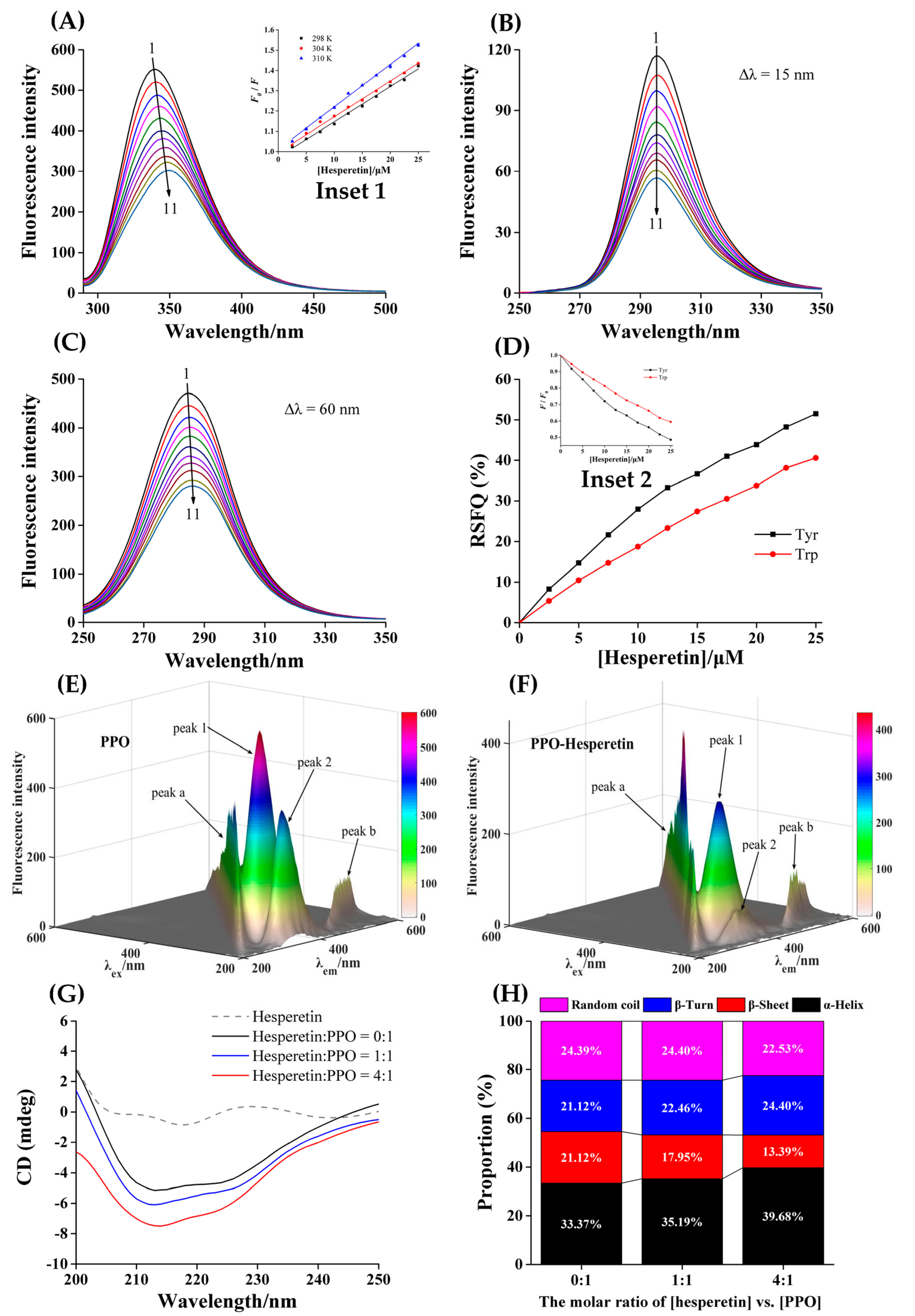
3.5. Intrinsic Fluorescence of PPO Quenched by Hesperetin
3.6. Mechanism of Fluorescence Quenching
3.7. Thermodynamic Parameters and Binding Forces
3.8. Conformational Changes of PPO
3.9. Molecular Docking
3.10. Molecular Dynamics
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| PPO | polyphenol oxidase |
| IC50 | half-maximum inhibitory concentration |
| MCR–ALS | multivariate curve resolution–alternate least squares |
| CD | circular dichroism |
| SVD | singular value decomposition |
| RSFQ | ratios of synchronous fluorescence quenching |
| 3D | three-dimensional |
| MD | molecular dynamics |
| RMSD | root mean square deviation |
| Rg | radius of gyration |
| RMSF | root mean square fluctuation |
| SASA | solvent accessible surface area |
References
- Huang, G.L.; Liu, T.T.; Ma, J.J.; Sun, X.L.; Sui, S.Y.; Quan, X.Y.; Wang, Y.M. Anti-polyphenol oxidase mechanism of oligomeric procyanidins and its application on browning control of “Baiyu” loquat during storage. Food Biosci. 2022, 48, 101791. [Google Scholar] [CrossRef]
- Alizadeh, N.; Sayahi, M.H.; Iraji, A.; Yazdaf, R.; Moazzam, A.; Mobaraki, K.; Adib, M.; Attarroshan, M.; Larijani, B.; Rastegar, H.; et al. Evaluating the effects of disubstituted 3-hydroxy-1H-pyrrol-2(5H)-one analog as novel tyrosinase inhibitors. Bioorg. Chem. 2022, 126, 105876. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, X.; Tao, G.J.; Chen, J.; Zheng, Z.P. Investigating the inhibitory activity and mechanism differences between norartocarpetin and luteolin for tyrosinase: A combinatory kinetic study and computational simulation analysis. Food Chem. 2017, 223, 40–48. [Google Scholar] [CrossRef]
- Xiong, Z.Q.; Zhou, L.; Liu, W. Research progress in the inhibition of Polyphenol oxidase. Sci. Technol. Food Ind. 2015, 21, 394–400. [Google Scholar]
- Zhang, Y.; Sun, Y.; Zhang, H.; Mai, Q.; Zhang, B.; Li, H.; Deng, Z. The degradation rules of anthocyanins from eggplant peel and antioxidant capacity in fortified model food system during the thermal treatments. Food Biosci. 2020, 38, 100701. [Google Scholar] [CrossRef]
- Zhou, H.L.; Wang, F.H.; Yi, J.J.; Cheng, F.Y.; Yuan, L.; Niu, H.H.; Zhou, L.Y. Research progress on the effect of chemical inhibitors on the properties of polyphenol oxidase in fruits and vegetables. Food Ferment. Ind. 2021, 47, 253–260. [Google Scholar]
- Lee, M.K.; Hwang, Y.H.; Ryu, H.; Lee, A.; Jeong, H.H.; Baek, J.; Kim, M.J.; Lee, J.Y.; Van, J.Y.; Liu, Y.S.; et al. Galla rhois water extract inhibits enzymatic browning in apple juice partly by binding to and inactivating polyphenol oxidase. Food Chem. 2022, 383, 132277. [Google Scholar] [CrossRef]
- Siew, Z.Z.; Chan, E.W.C.; Wong, C.W. Hydrophobic bioactive constituents of cinnamon bark as inhibitor of polyphenol oxidase from Musa acuminata ‘Mas’ peel. Biocatal. Agric. Biotechnol. 2022, 45, 102504. [Google Scholar] [CrossRef]
- Yu, K.B.; Zhou, L.; Sun, Y.F.; Zeng, Z.C.; Chen, H.W.; Liu, J.P.; Zou, L.Q.; Liu, W. Anti-browning effect of Rosa roxburghii on apple juice and identification of polyphenol oxidase inhibitors. Food Chem. 2021, 359, 129855. [Google Scholar] [CrossRef]
- Fan, M.H.; Zhang, G.W.; Hu, X.; Xu, X.M.; Gong, D.M. Quercetin as a tyrosinase inhibitor: Inhibitory activity, conformational change and mechanism. Food Res. Int. 2017, 100, 226–233. [Google Scholar] [CrossRef]
- Fan, M.H.; Ding, H.F.; Zhang, G.W.; Hu, X.; Gong, D.M. Relationships of dietary flavonoid structure with its tyrosinase inhibitory activity and affinity. LWT 2019, 107, 25–34. [Google Scholar] [CrossRef]
- Xiong, Z.Q.; Liu, W.; Zhou, L.; Zou, L.Q.; Chen, J. Mushroom (Agaricus bisporus) polyphenoloxidase inhibited by apigenin: Multi-spectroscopic analyses and computational docking simulation. Food Chem. 2016, 203, 430–439. [Google Scholar] [CrossRef]
- Jiang, H.W.; Zhou, L.; Sun, Y.F.; Yu, K.B.; Yu, W.Z.; Tian, Y.Q.; Liu, J.P.; Zou, L.Q.; Liu, W. Polyphenol oxidase inhibited by 4-hydroxycinnamic acid and naringenin: Multi-spectroscopic analyses and molecular docking simulation at different pH. Food Chem. 2022, 396, 133662. [Google Scholar] [CrossRef]
- Parhiz, H.; Roohbakhsh, A.; Soltani, F.; Rezaee, R.; Iranshahi, M. Antioxidant and anti-inflammatory properties of the citrus flavonoids hesperidin and hesperetin: An updated review of their molecular mechanisms and experimental models. Phytother. Res. 2015, 29, 323–331. [Google Scholar] [CrossRef]
- Roohbakhsh, A.; Parhiz, H.; Soltani, F.; Rezaee, R.; Iranshahi, M. Molecular mechanisms behind the biological effects of hesperidin and hesperetin for the prevention of cancer and cardiovascular diseases. Life Sci. 2015, 124, 64–74. [Google Scholar] [CrossRef]
- Moghaddam, A.H.; Zare, M. Neuroprotective effect of hesperetin and nano-hesperetin on recognition memory impairment and the elevated oxygen stress in rat model of Alzheimer’s disease. Biomed. Pharmacother. 2018, 97, 1096–1101. [Google Scholar]
- Si, Y.X.; Wang, Z.J.; Park, D.; Chung, H.Y.; Wang, S.F.; Yan, L.; Yang, J.M.; Qian, G.Y.; Yin, S.J.; Park, Y.D. Effect of hesperetin on tyrosinase: Inhibition kinetics integrated computational simulation study. Int. J. Biol. Macromol. 2012, 50, 257–262. [Google Scholar] [CrossRef]
- Xu, H.; Li, X.; Mo, L.; Zou, Y.; Zhao, G. Tyrosinase inhibitory mechanism and the anti-browning properties of piceid and its ester. Food Chem. 2022, 390, 133207. [Google Scholar] [CrossRef]
- Li, G.H.; Lee, Y.Y.; Lu, X.X.; Chen, J.; Liu, N.; Qiu, C.Y.; Wang, Y. Simultaneous loading of (−)-epigallocatechin gallate and ferulic acid in chitosan-based nanoparticles as effective antioxidant and potential skin-whitening agents. Int. J. Biol. Macromol. 2022, 219, 333–345. [Google Scholar] [CrossRef]
- Karakaya, G.; Türe, A.; Ercan, A.; Öncül, S.; Aytemir, M.D. Synthesis, computational molecular docking analysis and effectiveness on tyrosinase inhibition of kojic acid derivatives. Bioorg. Chem. 2019, 88, 102950. [Google Scholar] [CrossRef]
- Ding, H.F.; Wu, X.Q.; Pan, J.H.; Hu, X.; Gong, D.M.; Zhang, G.W. New insights into the inhibition mechanism of betulinic acid on α-glucosidase. J. Agric. Food Chem. 2018, 66, 7065–7075. [Google Scholar] [CrossRef]
- Li, J.; Gong, Y.H.; Li, J.W.; Fan, L.P. In vitro inhibitory effects of polyphenols from Tartary buckwheat on xanthine oxidase: Identification, inhibitory activity, and action mechanism. Food Chem. 2022, 379, 132100. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Ni, M.T.; Zhang, Y.; Zhang, G.W.; Pan, J.H.; Gong, D.M. Comparing the inhibitory abilities of epigallocatechin-3-gallate and gallocatechin gallate against tyrosinase and their combined effects with kojic acid. Food Chem. 2021, 349, 129172. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Hu, X.; Pan, J.H.; Gong, D.M.; Zhang, G.W. Mechanistic insights into the inhibition of pancreatic lipase by apigenin: Inhibitory interaction, conformational change and molecular docking studies. J. Mol. Liq. 2021, 335, 116505. [Google Scholar] [CrossRef]
- Pu, P.; Zheng, X.; Jiao, L.N.; Chen, L.; Yang, H.; Zhang, Y.H.; Liang, G.Z. Six flavonoids inhibit the antigenicity of β-lactoglobulin by noncovalent interactions: A spectroscopic and molecular docking study. Food Chem. 2021, 339, 128106. [Google Scholar] [CrossRef]
- Maurya, N.; Maurya, J.K.; Singh, U.K.; Dohare, R.; Zafaryab, M.; Moshahid Alam Rizvi, M.; Kumari, M.; Patel, R. In vitro cytotoxicity and interaction of noscapine with human serum albumin: Effect on structure and esterase activity of HSA. Mol. Pharm. 2019, 16, 952–966. [Google Scholar] [CrossRef]
- Cheng, D.; Wang, X.R.; Tang, J.L.; Zhang, X.Y.; Wang, C.L.; Li, H. Characterization of the binding mechanism and conformational changes of bovine serum albumin upon interaction with aluminum-maltol: A spectroscopic and molecular docking study. Metallomics 2019, 11, 1625–1634. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Huang, L.; Sun, C.Y.; Zhao, D.S.; Tang, H.J. Studies on the structure-activity relationship and interaction mechanism of flavonoids and xanthine oxidase through enzyme kinetics, spectroscopy methods and molecular simulations. Food Chem. 2020, 323, 126807. [Google Scholar] [CrossRef]
- Huang, G.; Jin, H.N.; Liu, G.C.; Yang, S.Y.; Jiang, L.Z.; Zhang, Y.; Sui, X.N. An insight into the changes in conformation and emulsifying properties of soy β-conglycinin and glycinin as affected by EGCG: Multi-spectral analysis. Food Chem. 2022, 394, 133484. [Google Scholar] [CrossRef]
- Murtaza, A.; Iqbal, A.; Linhu, Z.; Liu, Y.; Xu, X.; Pan, S.; Hu, W. Effect of high-pressure carbon dioxide on the aggregation and conformational changes of polyphenol oxidase from apple (Malus domestica) juice. Innov. Food Sci. Emerg. Technol. 2019, 54, 43–50. [Google Scholar] [CrossRef]
- Singh, M.; Thrimawithana, T.; Shukla, R.; Adhikari, B. Inhibition of enzymes associated with obesity by the polyphenol-rich extracts of Hibiscus sabdariffa. Food Biosci. 2022, 50, 101992. [Google Scholar] [CrossRef]
- Wang, C.; Chen, L.; Lu, Y.C.; Liu, J.; Zhao, R.; Sun, Y.H.; Sun, B.Y.; Wang, C.N. pH-Dependent complexation between β-lactoglobulin and lycopene: Multi-spectroscopy, molecular docking and dynamic simulation study. Food Chem. 2021, 362, 130230. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Lv, N.; Ren, G.; Wu, R.; Wang, B.; Cao, Z.; Xie, H. Explore the interaction mechanism between zein and EGCG using multi-spectroscopy and molecular dynamics simulation methods. Food Hydrocoll. 2021, 120, 106906. [Google Scholar] [CrossRef]
- Liu, L.L.; Li, J.D.; Zhang, L.L.; Wei, S.D.; Qin, Z.Y.; Liang, D.D.; Ding, B.M.; Chen, H.; Song, W. Conformational changes of tyrosinase caused by pentagalloylglucose binding: Implications for inhibitory effect and underlying mechanism. Food Res. Int. 2022, 157, 111312. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.M.; Zheng, Y.Y.; Lin, A.T.; Zhang, X.; Wu, X.Z.; Lin, J.; Xu, X.T.; Xiong, Z. Design, synthesis and evaluation of indole-based bisacylhydrazone derivatives as α-glucosidase inhibitors. J. Mol. Struct. 2023, 1271, 134124. [Google Scholar] [CrossRef]
- Mahdavi, A.; Mohammadsadeghi, N.; Mohammadi, F.; Saadati, F.; Nikfard, S. Evaluation of inhibitory effects of some novel phenolic derivatives on the mushroom tyrosinase activity: Insights from spectroscopic analyses, molecular docking and in vitro assays. Food Chem. 2022, 387, 132938. [Google Scholar] [CrossRef]
- Zheng, L.; Lee, J.; Yue, L.M.; Lim, G.T.; Yang, J.M.; Ye, Z.M.; Park, Y.D. Inhibitory effect of pyrogallol on α-glucosidase: Integrating docking simulations with inhibition kinetics. Int. J. Biol. Macromol. 2018, 112, 686–693. [Google Scholar] [CrossRef]
- Santa Rosa, L.N.; de Paula Rezende, J.; Coelho, Y.L.; Mendes, T.A.O.; da Silva, L.H.M.; dos Santos Pires, A.C. β-lactoglobulin conformation influences its interaction with caffeine. Food Biosci. 2021, 44, 101418. [Google Scholar] [CrossRef]
- Wen, Y.X.; Zhou, X.; Huo, D.; Chen, J.C.; Weng, L.M.; Li, B.; Wu, Z.Q.; Zhang, X.; Li, L. Optimization for the extraction of polysaccharides from Huidouba and their in vitro α-glucosidase inhibition mechanism. Food Biosci. 2022, 49, 101910. [Google Scholar] [CrossRef]
- Fu, J.J.; Sun, C.; Tan, Z.F.; Zhang, G.Y.; Chen, G.B.; Song, L. Nanocomplexes of curcumin and glycated bovine serum albumin: The formation mechanism and effect of glycation on their physicochemical properties. Food Chem. 2022, 368, 130651. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.J.; Chen, W.; Pan, F.; Zhao, P.P.; He, Y.T.; Yu, R.; Fu, R.X.; Munkh-Amgalan, G.; Jiang, Z.M. Characterization of the binding behavior, structure and foaming properties of bovine α-lactalbumin combined with saponin by the multi-spectroscopic and silico approaches. Food Hydrocoll. 2022, 124, 107259. [Google Scholar] [CrossRef]
- Sadeghi-Kaji, S.; Shareghi, B.; Saboury, A.A.; Farhadian, S. Investigating the interaction of porcine pancreatic elastase and propanol: A spectroscopy and molecular simulation study. Int. J. Biol. Macromol. 2020, 146, 687–691. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.H.; Hu, X.; Pan, J.H.; Gong, D.M.; Zhang, G.W. Effects of interaction between hesperetin/hesperidin and glutenin on the structure and functional properties of glutenin. LWT 2022, 155, 112983. [Google Scholar] [CrossRef]
- Ross, P.D.; Subramanian, S. Thermodynamics of protein association reactions: Forces contributing to stability. Biochemistry 1981, 20, 3096–3102. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Peng, S.S.; Zhu, F.W.; Lei, X.L.; Xiao, Q.; Su, W.; Liu, Y.; Huang, C.S.; Zhang, L.X. Multispectroscopic investigation of the interaction between two ruthenium (II) arene complexes of curcumin analogs and human serum albumin. Biol. Trace Elem. Res. 2016, 169, 189–203. [Google Scholar] [CrossRef]
- Yu, Z.Y.; Xu, K.; Wang, X.; Wen, Y.T.; Wang, L.J.; Huang, D.Q.; Chen, X.X.; Chai, W.M. Punicalagin as a novel tyrosinase and melanin inhibitor: Inhibitory activity and mechanism. LWT 2022, 161, 113318. [Google Scholar] [CrossRef]
- Greenfield, N.J. Applications of circular dichroism in protein and peptide analysis. TrAC-Trends Anal. Chem. 1999, 18, 236–244. [Google Scholar] [CrossRef]
- Zhou, H.L.; Wang, F.H.; Niu, H.H.; Yuan, L.; Tian, J.; Cai, S.B.; Bi, X.F.; Zhou, L.Y. Structural studies and molecular dynamic simulations of polyphenol oxidase treated by high pressure processing. Food Chem. 2022, 372, 131243. [Google Scholar] [CrossRef]
- Feng, Y.; Sun, Y.; Meng, Z.; Sui, X.; Zhang, D.; Yan, H.; Wang, Q. S-Ethyl thioacetate as a natural anti-browning agent can significantly inhibit the browning of fresh-cut potatoes by decreasing polyphenol oxidase activity. Sci. Hortic. 2022, 305, 111427. [Google Scholar] [CrossRef]
- Das, S.; Hazarika, Z.; Sarmah, S.; Baruah, K.; Rohman, M.A.; Paul, D.; Jha, A.N.; Roy, A.S. Exploring the interaction of bioactive kaempferol with serum albumin, lysozyme and hemoglobin: A biophysical investigation using multi-spectroscopic, docking and molecular dynamics simulation studies. J. Photochem. Photobiol. B Biol. 2020, 205, 111825. [Google Scholar] [CrossRef]
- Kooravand, M.; Asadpour, S.; Haddadi, H.; Farhadian, S. An insight into the interaction between malachite green oxalate with human serum albumin: Molecular dynamic simulation and spectroscopic approaches. J. Hazard. Mater. 2021, 407, 124878. [Google Scholar] [CrossRef]
- Ni, M.T.; Hu, X.; Gong, D.M.; Zhang, G.W. Inhibitory mechanism of vitexin on α-glucosidase and its synergy with acarbose. Food Hydrocoll. 2020, 105, 105782. [Google Scholar] [CrossRef]
- Lu, Y.; Zhao, R.; Wang, C.; Zhang, X.; Wang, C. Deciphering the non-covalent binding patterns of three whey proteins with rosmarinic acid by multi-spectroscopic, molecular docking and molecular dynamics simulation approaches. Food Hydrocoll. 2022, 132, 107895. [Google Scholar] [CrossRef]
- Zhou, H.; Bie, S.; Li, J.; Yuan, L.; Zhou, L. Comparison on inhibitory effect and mechanism of inhibitors on sPPO and mPPO purified from ‘Lijiang snow’ peach by combining multispectroscopic analysis, molecular docking and molecular dynamics simulation. Food Chem. 2023, 400, 134048. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T (K) | Ksv (×104 L mol−1) | Ra | Ka (×104 L mol−1) | Rb | n | ΔH° (kJ mol−1) | ΔG° (kJ mol−1) | ΔS° (J mol−1K−1) |
|---|---|---|---|---|---|---|---|---|
| 298 | 1.71 ± 0.01 a | 0.9982 | 1.98 ± 0.01 a | 0.9988 | 1.23 ± 0.03 | 17.47 | −24.49 | 107.22 |
| 304 | 1.76 ± 0.02 b | 0.9976 | 2.05 ± 0.02 b | 0.9914 | 1.08 ± 0.01 | −25.13 | ||
| 310 | 2.09 ± 0.02 c | 0.9979 | 2.22 ± 0.05 c | 0.9987 | 0.99 ± 0.04 | −25.77 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, X.; Luo, X.; Wang, L.; Gong, D.; Zhang, G. New Insights into the Inhibition of Hesperetin on Polyphenol Oxidase: Inhibitory Kinetics, Binding Characteristics, Conformational Change and Computational Simulation. Foods 2023, 12, 905. https://doi.org/10.3390/foods12040905
Hong X, Luo X, Wang L, Gong D, Zhang G. New Insights into the Inhibition of Hesperetin on Polyphenol Oxidase: Inhibitory Kinetics, Binding Characteristics, Conformational Change and Computational Simulation. Foods. 2023; 12(4):905. https://doi.org/10.3390/foods12040905
Chicago/Turabian StyleHong, Xinyue, Xiaoqiao Luo, Langhong Wang, Deming Gong, and Guowen Zhang. 2023. "New Insights into the Inhibition of Hesperetin on Polyphenol Oxidase: Inhibitory Kinetics, Binding Characteristics, Conformational Change and Computational Simulation" Foods 12, no. 4: 905. https://doi.org/10.3390/foods12040905
APA StyleHong, X., Luo, X., Wang, L., Gong, D., & Zhang, G. (2023). New Insights into the Inhibition of Hesperetin on Polyphenol Oxidase: Inhibitory Kinetics, Binding Characteristics, Conformational Change and Computational Simulation. Foods, 12(4), 905. https://doi.org/10.3390/foods12040905