

Comparative Metabolomic Analysis of the Nutritional Aspects from Ten Cultivars of the Strawberry Fruit


Abstract
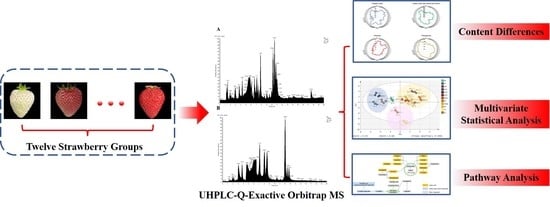
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Reagents
2.3. Sample Preparation for Metabolite Extraction
2.4. Non-Targeted Metabolomics by Ultra-HPLC (UHPLC) Q-Exactive Orbitrap MS
2.5. Data Processing and Statistical Analysis
3. Results and Discussion
3.1. Morphological Differences among Strawberry Cultivars
3.2. Identification of Metabolites in Strawberries
3.3. Multivariate Statistical Analysis of the Identified Metabolites in Different Strawberry Cultivars
3.4. Potential Biomarkers for Distinguishing Different Strawberry Cultivars
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wise, K.; Wedding, T.; Selby-Phanm, J. Application of automated image colour analyses for the early-prediction of strawberry development and quality. Sci. Hortic. 2022, 304, 111316. [Google Scholar] [CrossRef]
- Aaby, K.; Skrede, G.; Wrolstad, R.E. Phenolic composition and antioxidant activities in flesh and achenes of strawberries (Fragaria ananassa). J. Agric. Food Chem. 2005, 53, 4032–4040. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Lim, S.; Lau, H.; Ni, W.; Li, S. Effect of glycinebetaine on metabolite proffles of cold-stored strawberry revealed by 1 H NMR-based metabolomics. Food Chem. 2022, 393, 133452. [Google Scholar] [CrossRef] [PubMed]
- Giampieri, F.; Tulipani, S.; Alvarez-Suarez, J.; Quiles, J.; Mezzetti, B.; Battino, M. The strawberry: Composition, nutritional quality, and impact on human health. Nutrition 2012, 28, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Stürtz, M.; Cerezo, A.B.; Cantos-Villar, E.; Garcia-Parrilla, M.C. Determination of the melatonin content of different varieties of tomatoes (Lycopersicon esculentum) and strawberries (Fragaria ananassa). Food Chem. 2011, 127, 1329–1334. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, T.; Gama, F.; Rodrigues, M.A.; Abadía, J.; Varennes, A.; Pestana, M.; Silva, J.P.; Correia, P.J. Effects of foliar application of organic acids on strawberry plants. Plant Physiol. Biochem. 2022, 188, 12–20. [Google Scholar] [CrossRef]
- Mink, P.J.; Scraffrord, C.G.; Barraj, L.M.; Harnack, L.; Hong, C.; Nettleton, J.A.; Jacobs, D.R. Flavonoid intake and cardiovascular disease mortality: A prospective study in postmenopausal women. Am. J. Clin. Nutr. 2007, 85, 895–909. [Google Scholar] [CrossRef] [Green Version]
- Pinto, M.S.; Carvalho, J.M.; Lajolo, F.M.; Genovese, M.I.; Shetty, K. Evaluation of antiproliferative, anti-type 2 diabetes, and antihypertension Potentials of ellagitannins from strawberries (Fragaria ananassa Duch.) using in vitro models. J. Med. Food 2010, 13, 1027–1035. [Google Scholar] [CrossRef]
- Navarro-Hortal, M.D.; Romero-Marquez, J.M.; Esteban-Munoz, A.; Sanchez-Gonzalez, C.; Rivas-García, L.; Llopis, J.; Cianciosi, D.; Giampieri, F.; Sumalla-Cano, S.; Battino, M.; et al. Strawberry (Fragaria × ananassa cv. Romina) methanolic extract attenuates Alzheimer’s beta amyloid production and oxidative stress by SKN-1/NRF and DAF-16/FOXO mediated mechanisms in C. elegans. Food Chem. 2022, 372, 131272. [Google Scholar] [CrossRef]
- Seeram, N.P.; Adams, L.S.; Zhang, Y.; Lee, R.; Sand, D.; Scheuller, H.S.; Heber, D. Blackberry, black raspberry, blueberry, cranberry, red raspberry, and strawberry extracts inhibit growth and stimulate apoptosis of human cancer cells in vitro. J. Agric. Food Chem. 2006, 54, 9329–9339. [Google Scholar] [CrossRef]
- Macias-Rodriguez, L.; Quero, E.; Lopez, M.G. Carbohydrate differences in strawberry crowns and fruit (Fragaria × ananassa) during plant development. J. Agric. Food Chem. 2002, 50, 3317–3321. [Google Scholar] [CrossRef]
- Mellado-Mojica, E.; Calvo-Gómez, O.; Jofre-Garfifias, A.E.; Dávalos-González, P.A.; Desjardins, V.; López, M.G. Fructooligosaccharides as molecular markers of geographic origin, growing region, genetic background and prebiotic potential in strawberries: A TLC, HPAEC-PAD and FTIR study. Food Chem. Adv. 2022, 1, 100064. [Google Scholar] [CrossRef]
- Samykanno, K.; Pang, E.; Marriott, P.J. Chemical characterisation of two Australian-grown strawberry varieties by using comprehensive two-dimensional gas chromatography–mass spectrometry. Food Chem. 2013, 142, 11075–11080. [Google Scholar] [CrossRef]
- Ono, E.; Mizutani, M.F.; Nakamura, N.; Fukui, Y.; Sakakibara, K.Y.; Yamaguchi, M.; Nakayama, T.; Tanaka, T.; Kusumi, T.; Tanaka, Y. Yellow Flowers Generated by Expression of the Aurone Biosynthetic Pathway. Proc. Natl. Acad. Sci. USA 2006, 103, 1997–2005. [Google Scholar] [CrossRef] [Green Version]
- Duan, W.; Shao, W.; Lin, W.; Yuan, L.; Chen, L.; Zagorchev, L.; Li, J. Integrated metabolomics and transcriptomics reveal the differences in fruit quality of the red and white Fragaria pentaphylla morphs. Food Biosci. 2021, 40, 100896. [Google Scholar] [CrossRef]
- Sirijan, M.; Drapal, M.; Chaiprasart, P.; Fraser, P.D. Characterisation of thai strawberry (Fragaria × ananassa Duch.) cultivars food research international with RAPD markers and metabolite profiling techniques. Phytochemistry 2020, 180, 112522. [Google Scholar] [CrossRef]
- Dubrow, G.A.; Forero, D.P.; Peterson, D.G. Identification of volatile compounds correlated with consumer acceptability of strawberry preserves: Untargeted GC–MS analysis. Food Chem. 2022, 378, 132043. [Google Scholar] [CrossRef]
- Kim, A.N.; Lee, K.Y.; Han, C.Y.; Kim, H.J.; Choi, S.G. Effect of an oxygen-free atmosphere during heating on anthocyanin, organic acid, and color of strawberry puree. J. Food Biosci. 2022, 50, 102065. [Google Scholar] [CrossRef]
- Shen, J.; Shao, W.; Du, Z.; Lu, H.; Li, J. Integrated metabolomic and transcriptomic analyses reveal differences in the biosynthetic pathway of anthocyanins in Fragaria nilgerrensis and Fragaria pentaphylla. Sci. Hortic. 2020, 271, 109476. [Google Scholar] [CrossRef]
- Kårlund, A.; Hanhineva, K.; Lehtonen, M.; McDougall, G.J.; Stewart, D.; Karjalainen, R.O. Non-targeted metabolite profiling highlights the potential of strawberry leaves as a resource for specific bioactive compounds. J. Agric. Food Chem. 2017, 97, 2182–2190. [Google Scholar] [CrossRef] [Green Version]
- Lou, Z.; Wang, H.; Zhu, S.; Ma, C.; Wang, Z. Antibacterial activity and mechanism of action of chlorogenic acid. J. Food Sci. 2017, 76, M398–M403. [Google Scholar] [CrossRef] [PubMed]
- Pincemail, J.; Kevers, C.; Tabart, J.; Defraigne, J.O.; Dommes, J. Cultivars, Culture Conditions, and Harvest Time Influence Phenolic and Ascorbic Acid Contents and Antioxidant Capacity of Strawberry (Fragaria x ananassa). J. Food Sci. 2012, 77, 205–207. [Google Scholar] [CrossRef] [PubMed]
- Odriozola-Serrano, I.; Garde-Cerda´n, T.; Soliva-Fortuny, R.; Martı´n-Belloso, O. Differences in free amino acid profile of non-thermally treated tomato and strawberry juices. J. Food Compos. Anal. 2013, 32, 51–58. [Google Scholar] [CrossRef]
- Bingo¨l, L.; Türkyılmaz, M.; Ozkan, M. Increase in thermal stability of strawberry anthocyanins with amino acid copigmentation. Food Chem. 2022, 384, 132518. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.; Tong, Z.; Dong, X.; Sun, M.; Gao, T.; Duan, J. Simultaneous determination of 98 pesticide residues in strawberries using UPLC-MS/MS and GC-MS/MS. Microchem. J. 2020, 156, 104975. [Google Scholar] [CrossRef]
- Akhatou, I.; Domínguez, R.G.; Recamales, A.F. Investigation of the effect of genotype and agronomic conditions on metabolomic profiles of selected strawberry cultivars with different sensitivity to environmental stress. Plant Physiol. Biochem. 2016, 101, 14–22. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, W.; Wang, L.; Liu, H.; Liu, H. Regulating effects of the biophilic environment with strawberry plants on psychophysiological health and cognitive performance in small spaces. Build. Environ. 2022, 212, 108801. [Google Scholar] [CrossRef]
- Wang, Q.; Wei, Y.; Jiang, S.; Wang, X.; Xu, F.; Wang, H.; Shao, X. Flavor development in peach fruit treated with 1-methylcyclopropene during shelf storage. Food Res. Int. 2020, 137, 109653. [Google Scholar] [CrossRef]
- Byeon, S.E.; Lee, J. Differential responses of fruit quality and major targeted metabolites in three different cultivars of cold-stored figs (Ficus carica L.). Sci. Hortic. 2020, 260, 108877. [Google Scholar] [CrossRef]
- Cadi, H.E.; Cadi, A.E.; Kounnoun, A.; Majdoub, Y.O.E.; Lovillo, M.P.; Brigui, J.; Dugo, P.; Mondello, L.; Cacciola, F. Wild strawberry (Arbutus unedo): Phytochemical screening and antioxidant properties of fruits collected in northern Morocco. Arabian J. Chem. 2020, 13, 6299–6311. [Google Scholar] [CrossRef]
- Thomas-Valdés, S.; Theoduloz, C.; Jiménez-Aspee, F. Effect of simulated gastrointestinal digestion on polyphenols and bioactivity of the native Chilean red strawberry (Fragaria chiloensis ssp. chiloensis f. patagonica). Food Res. Int. 2019, 123, 106–114. [Google Scholar] [CrossRef]
- Cerezo, A.B.; Cuevas, E.; Winterhalter, P.; Parrilla, M.C.; Troncoso, A.M. Isolation, identification, and antioxidant activity of anthocyanin compounds in Camarosa strawberry. Food Chem. 2010, 123, 574–582. [Google Scholar] [CrossRef]
- Chen, W.; Xie, C.; He, Q.; Sun, J.; Bai, W. Improvement in color expression and antioxidant activity of strawberry juice fermented with lactic acid bacteria: A phenolic-based research. Food Chem. 2023, 17, 100535. [Google Scholar] [CrossRef]
- Cao, Y.; Zhao, B.; Li, Y.; Gao, H.; Xia, Q. Investigation of the difference in color enhancement effect on cyanidin-3-Oglucoside by different phenolic acids and the interaction mechanism. Food Chem. 2023, 411, 135409. [Google Scholar] [CrossRef] [PubMed]
- Ortega, R.H.; Fernández, M.A.; Cerezo, A.B.; Troncoso, A.M.; Parrilla, M.C. Influence of storage conditions on the anthocyanin profile and colour of an innovative beverage elaborated by gluconic fermentation of strawberry. J. Funct. Foods. 2016, 23, 198–209. [Google Scholar] [CrossRef]
- Garcia, C.; Blesso, C.N. Antioxidant properties of anthocyanins and their mechanism of action in atherosclerosis. Free Radical Biol. Med. 2021, 172, 152–166. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Wu, L.; Qiu, J.; Qian, Y.; Wang, M. Comparative Metabolomic Analysis of the Nutritional Aspects from Ten Cultivars of the Strawberry Fruit. Foods 2023, 12, 1153. https://doi.org/10.3390/foods12061153
Wang X, Wu L, Qiu J, Qian Y, Wang M. Comparative Metabolomic Analysis of the Nutritional Aspects from Ten Cultivars of the Strawberry Fruit. Foods. 2023; 12(6):1153. https://doi.org/10.3390/foods12061153
Chicago/Turabian StyleWang, Xinlu, Linxia Wu, Jing Qiu, Yongzhong Qian, and Meng Wang. 2023. "Comparative Metabolomic Analysis of the Nutritional Aspects from Ten Cultivars of the Strawberry Fruit" Foods 12, no. 6: 1153. https://doi.org/10.3390/foods12061153
APA StyleWang, X., Wu, L., Qiu, J., Qian, Y., & Wang, M. (2023). Comparative Metabolomic Analysis of the Nutritional Aspects from Ten Cultivars of the Strawberry Fruit. Foods, 12(6), 1153. https://doi.org/10.3390/foods12061153