
The Study of Yak Colostrum Nutritional Content Based on Foodomics
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Sample Collection
2.2. Determination of Fatty Acids with Targeted Metabolism Based on Gas Chromatography−Mass Spectrometer (GC−MS)
2.3. Determination of Amino Acids and Their Derivatives with Targeted Metabolism Based on Liquid Chromatography Tandem Quadrupole Mass Spectrometer (UHPLC−MS)
2.4. Lipids Extraction and MS Data of Non−Targeted Lipidomics Based on UHPLC−MS
2.5. Metabolites Extraction and MS Data of Non−Targeted Metabolomics Based on Ultra High Performance Liquid Chromatography Tandem Quadrupole Time of Flight Mass Spectrometer (UHPLC−TOF−MS)
2.6. Statistical Analysis
3. Results
3.1. Content of Fatty Acids in Yak Colostrum and Mature Milk
3.2. Content of Amino Acids and Their Derivatives in Yak Colostrum and Mature Milk
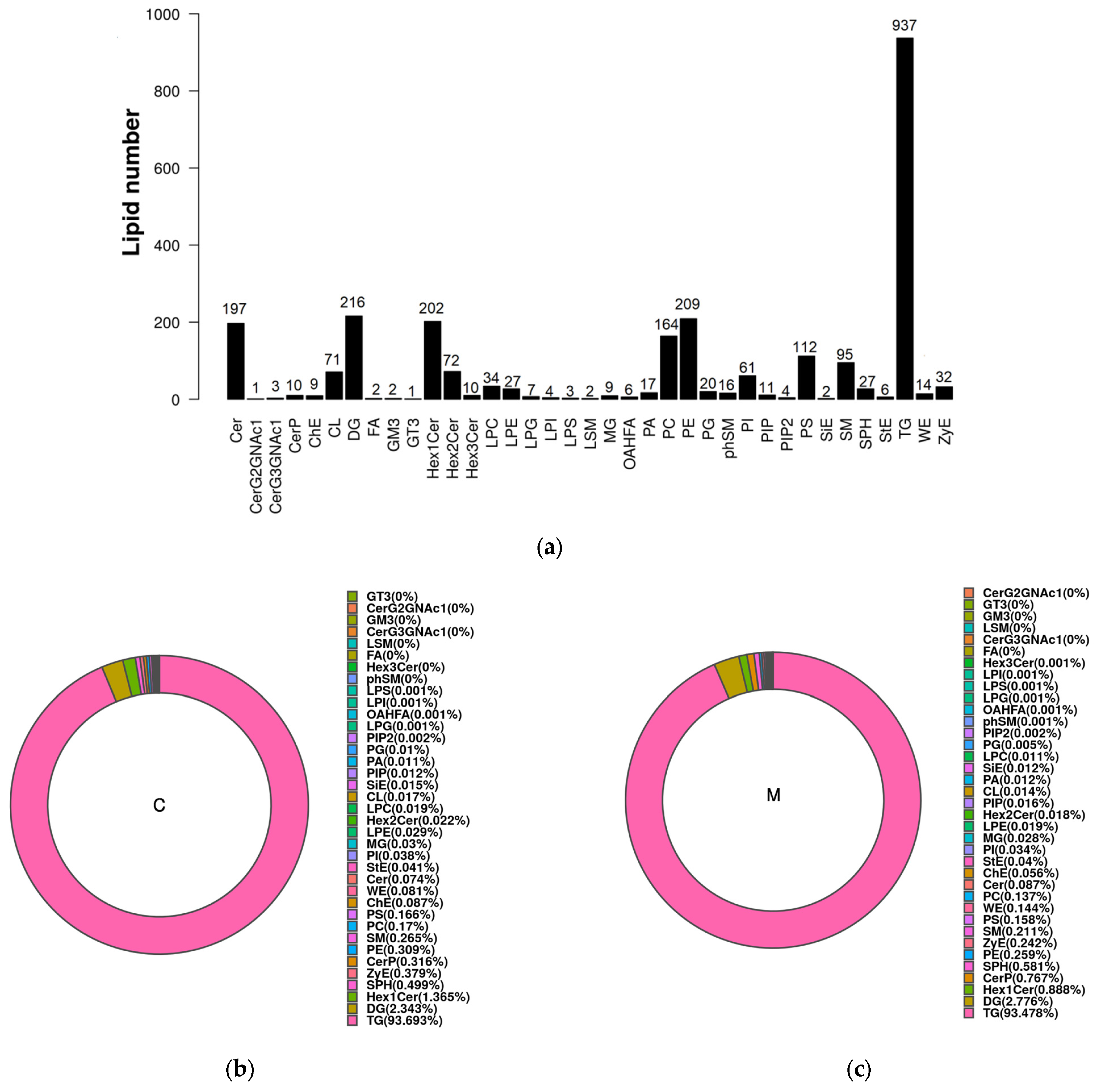
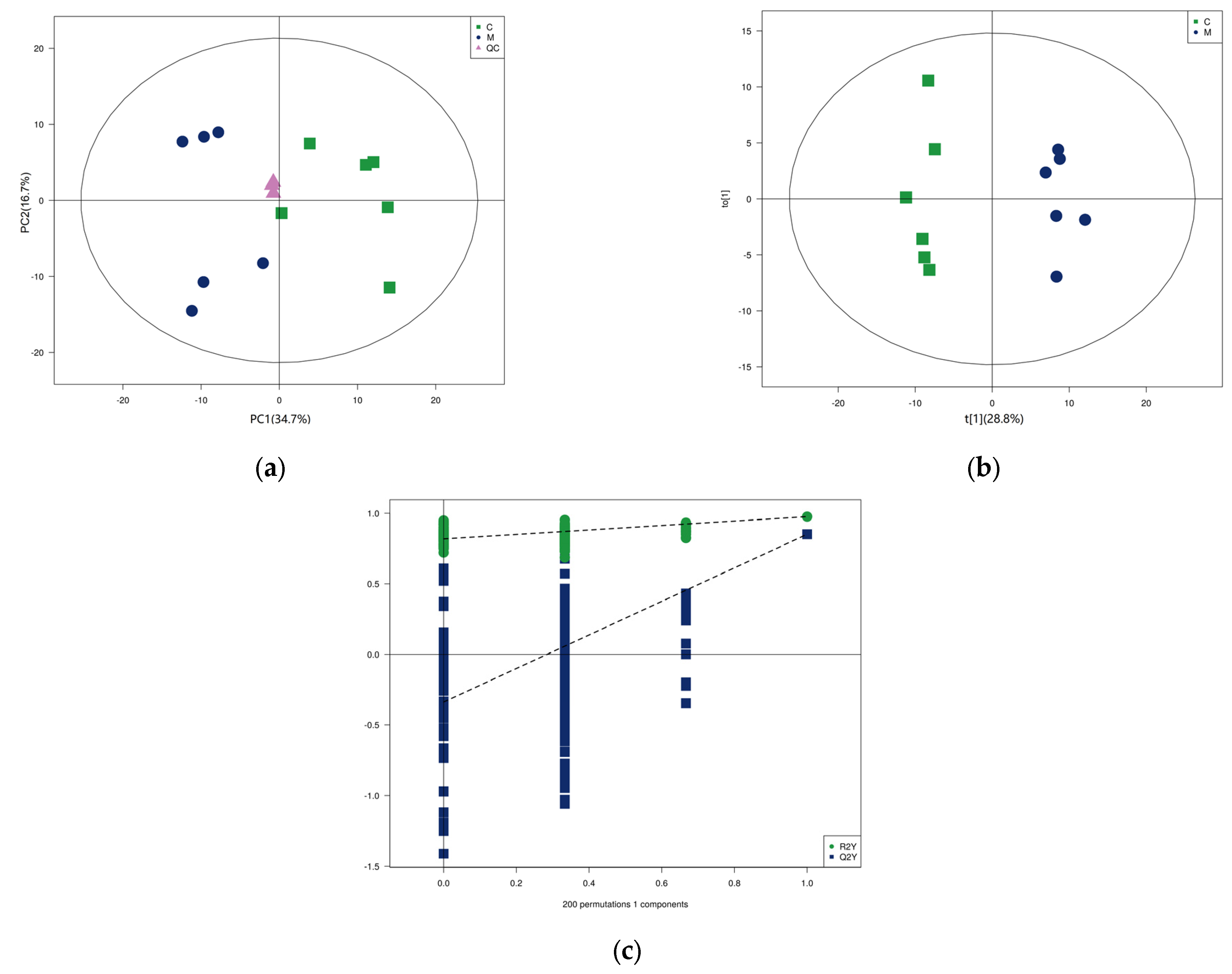
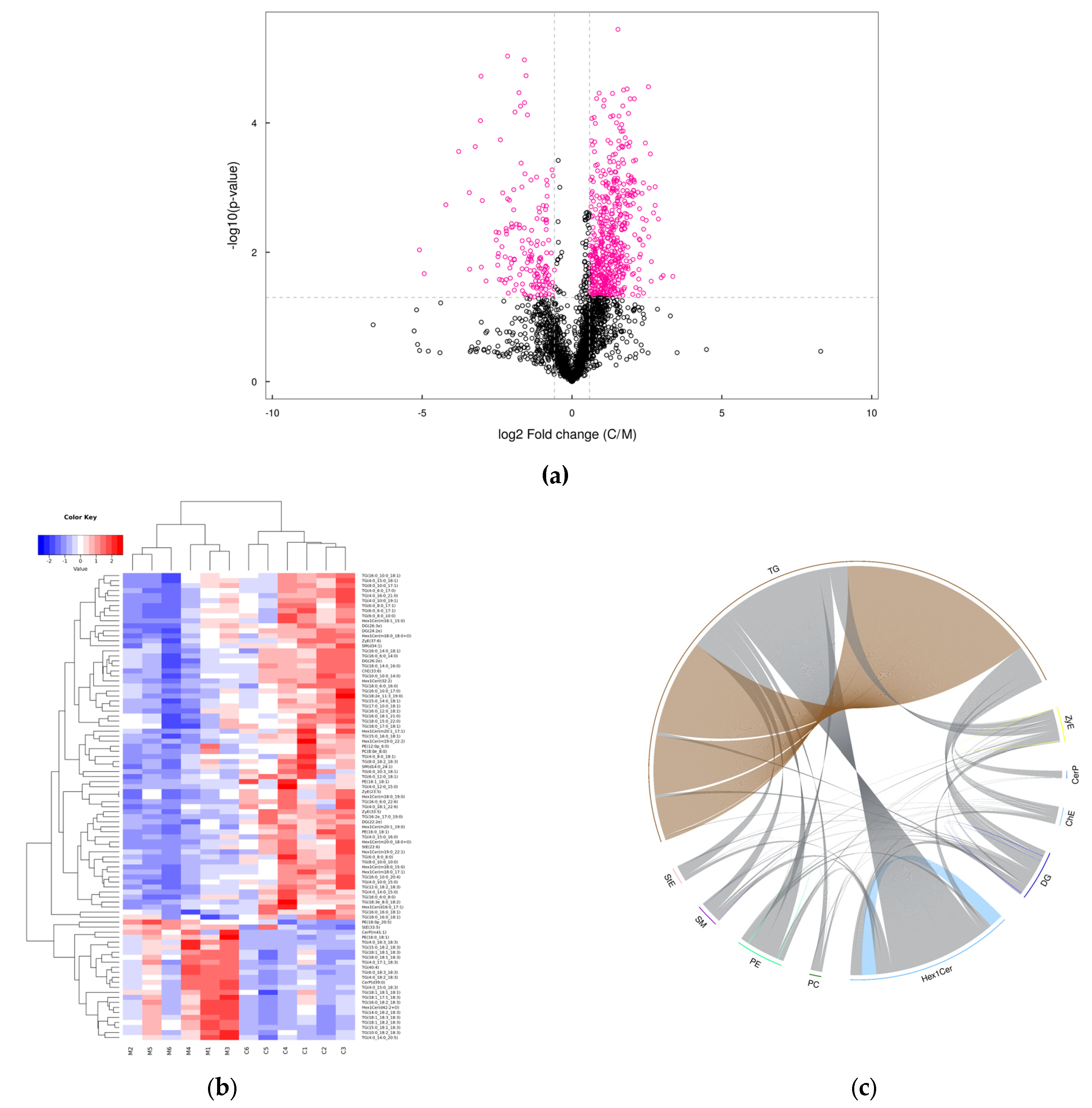
3.3. Lipidomics in Yak Colostrum and Mature Milk
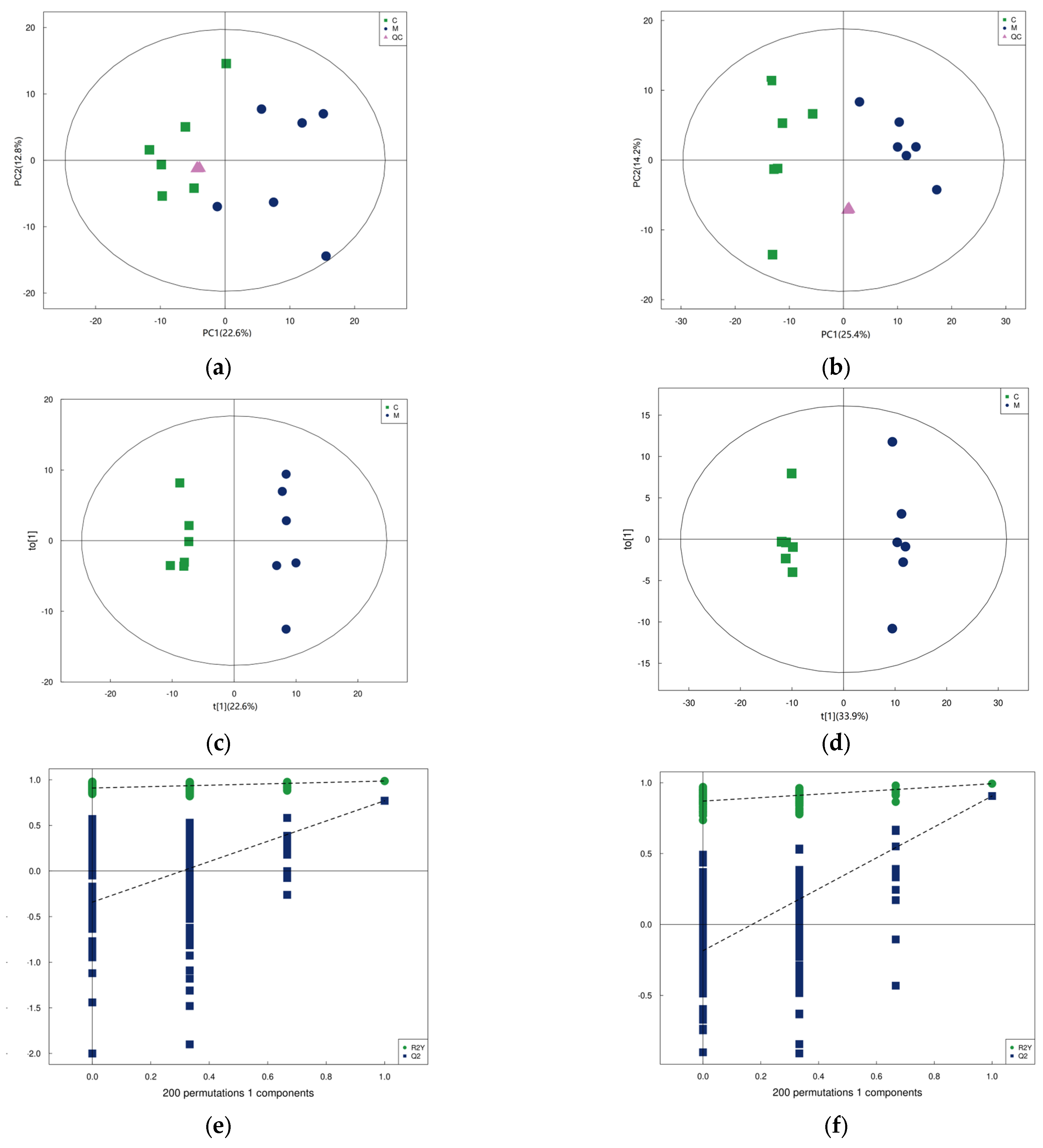
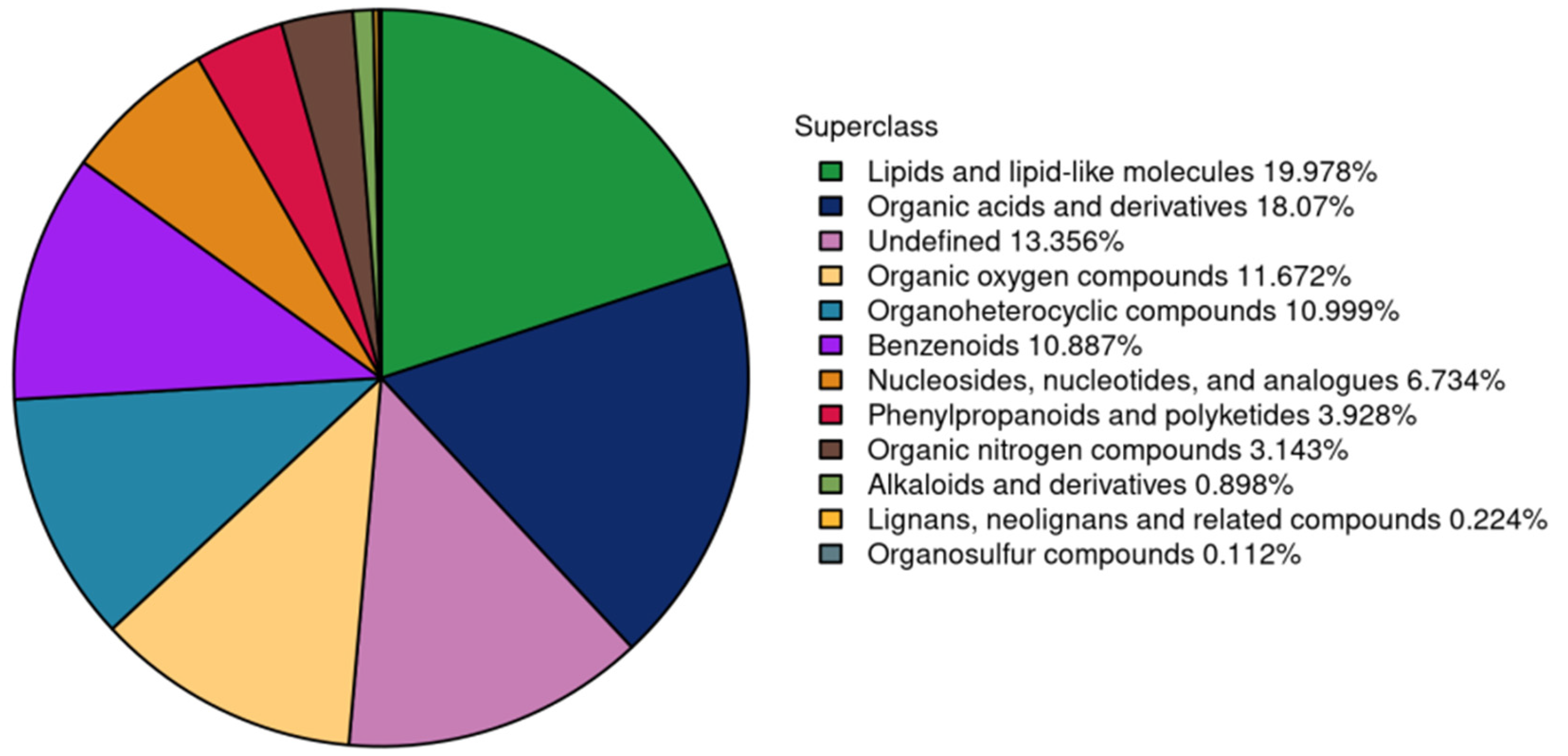
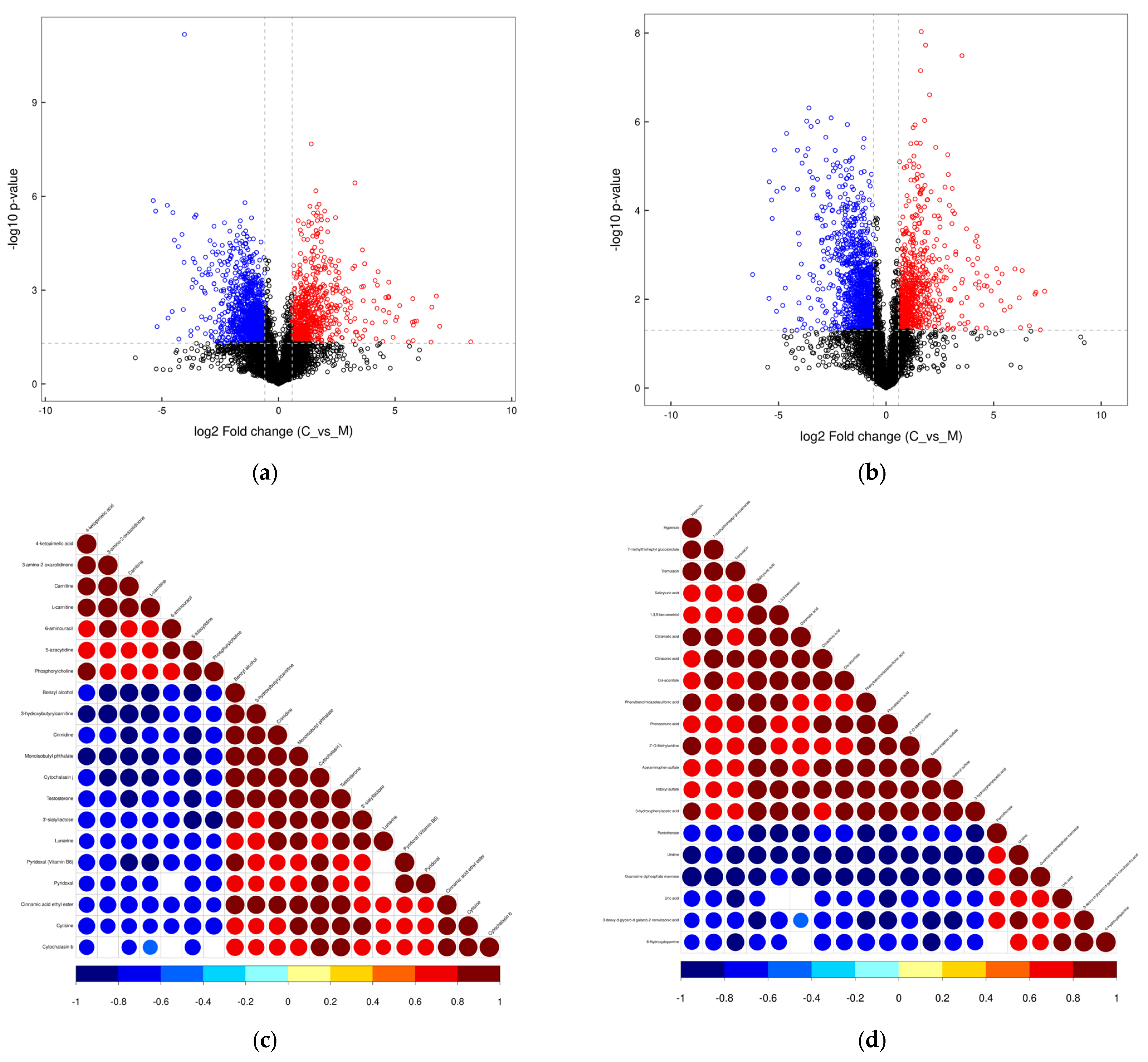
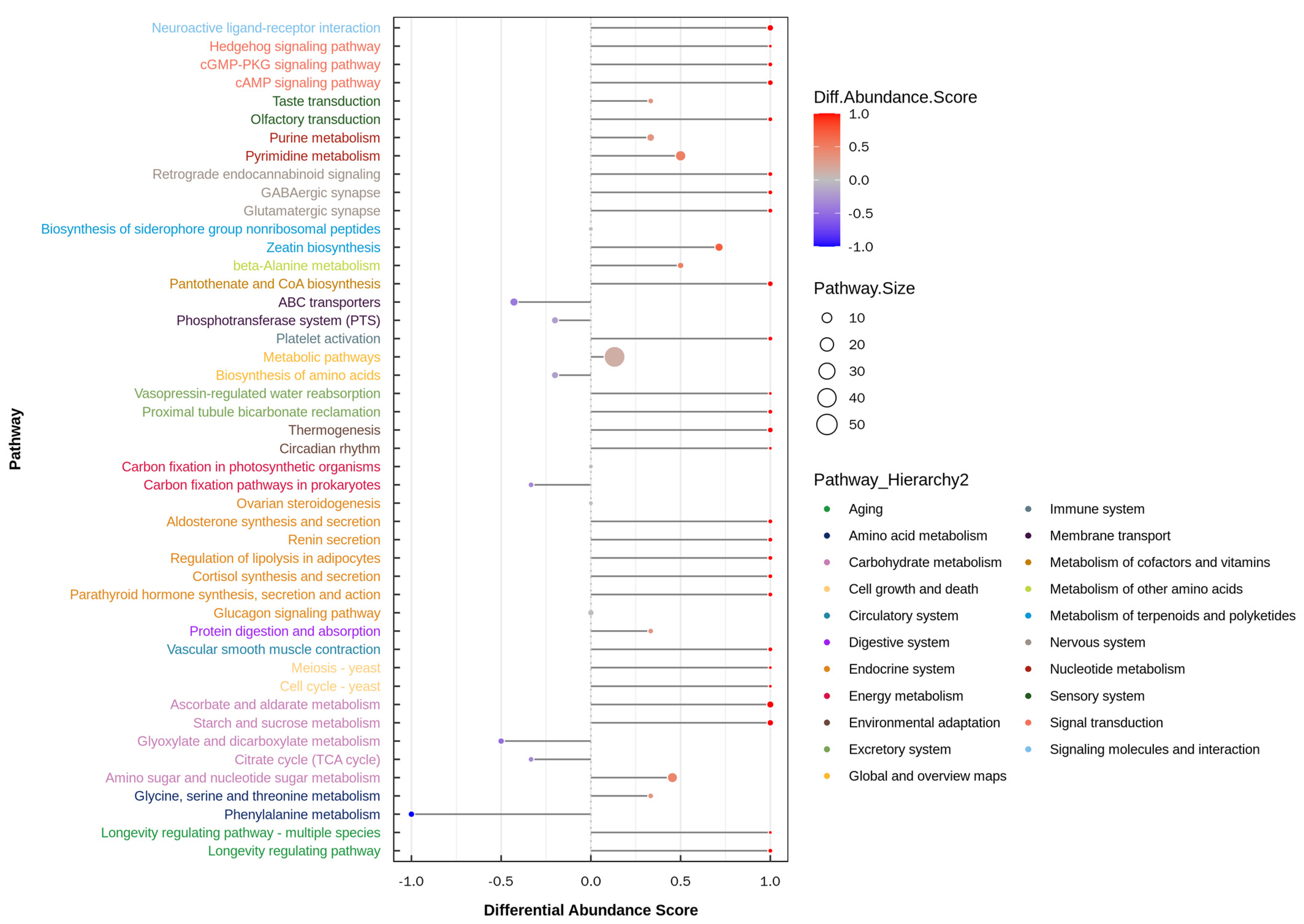
3.4. Metabolome in Yak Colostrum and Mature Milk
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El-fattah, A.M.A.; Rabo, F.H.A.; El-dieb, S.M. Changes in composition of colostrum of Egyptian buffaloes and Holstein cows. BMC Vet. Res. 2012, 8, 19. [Google Scholar] [CrossRef]
- Puppel, K.; Gołębiewski, M.; Grodkowski, G.; Slósarz, J.; Kunowska-Slósarz, M.; Solarczyk, P.; Łukasiewicz, M.; Balcerak, M.; Przysucha, T. Composition and Factors Affecting Quality of Bovine Colostrum: A Review. Animals 2019, 9, 1070. [Google Scholar] [CrossRef] [PubMed]
- Saad, K.; Abo-Elela, M.G.M.; El-Baseer, K.A.A.; Ahmed, A.E.; Ahmad, F.A.; Tawfeek, M.S.K.; El-Houfey, A.A.; Khair, M.D.A.; Abdel-Salam, A.M.; Amir Abo-Elgheit, A.; et al. Effects of bovine colostrum on recurrent respiratory tract infections and diarrhea in children. Medicine 2016, 95, e4560. [Google Scholar] [CrossRef]
- Eslamian, G.; Ardehali, S.H.; Baghestani, A.-R.; Shariatpanahi, Z.V. Effects of early enteral bovine colostrum supplementation on intestinal permeability in critically ill patients: A randomized, double-blind, placebo-controlled study. Nutrition 2018, 60, 106–111. [Google Scholar] [CrossRef]
- Ulfman, L.H.; Leusen, J.H.W.; Savelkoul, H.F.J.; Warner, J.O.; Neerven, R.J.J.V. Effects of bovine immunoglobulins on immune function, allergy, and infection. Front. Nutr. 2018, 5, 52. [Google Scholar] [CrossRef]
- Qiu, Q.; Zhang, G.J.; Ma, T.; Qian, W.B.; Wang, J.Y.; Ye, Z.Q.; Cao, C.C.; Hu, Q.J.; Kim, J.; Larkin, D.M.; et al. The yak genome and adaptation to life at high altitude. Nat. Genet. 2012, 44, 946–949. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ma, Y.; Li, Q.; Wang, J.; Cheng, J.; Xue, J.; Shi, J. The chemical composition and nitrogen distribution of Chinese Yak (Maiwa) Milk. Int. J. Mol. Sci. 2011, 12, 4885–4895. [Google Scholar] [CrossRef]
- Lin, K.; Zhang, W.; Han, X.; Xin, L.; Meng, Z.X.; Gong, P.M.; Cheng, D.Y. Yak milk casein as potential precursor of angiotensin I-converting enzyme inhibitory peptides based on in silico proteolysis. Food Chem. 2018, 253, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Braconi, D.; Bernardini, G.; Millucci, L.; Santucci, A. Foodomics for human health: Current status and perspectives. Expert Rev. Proteomic. 2018, 15, 153–164. [Google Scholar] [CrossRef]
- García-Cañas, V.; Simó, C.; Herrero, M.; Ibáñez, E.; Cifuentes, A.; Chem, A. Present and future challenges in food analysis: Foodomics. Anal. Chem. 2012, 84, 10150–10159. [Google Scholar] [CrossRef]
- Li, Q.Q.; Zhao, Y.; Zhu, D.; Pang, X.M.; Liu, Y.; Frew, R.; Chen, G. Lipidomics profiling of goat milk, soymilk and bovine milk by UPLC-Q-Exactive Orbitrap Mass Spectrometry. Food Chem. 2017, 224, 302–309. [Google Scholar] [CrossRef]
- Ten-Domenech, I.; Ramos-Garcia, V.; Moreno-Torres, M.; Parra-Llorca, A.; Gormaz, M.; Vento, M.; Kuligowski, J.; Quintas, G. The effect of Holder pasteurization on the lipid and metabolite composition of human milk. Food Chem. 2022, 384, 132581. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhu, Z.B.; Jia, W. Molecular mechanism associated with the use of magnetic fermentation in modulating the dietary lipid composition and nutritional quality of goat milk. Food Chem. 2022, 366, 130554. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yang, K.; Wei, X.; Qiao, W.; Chen, L. Untargeted metabolomics analysis reveals dynamic changes in co-fermentation with human milk-derived probiotics and Poria cocos. Front. Microbiol. 2022, 13, 1032870. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Mir, M.U.R.; Ganie, S.A.; Mushtaq, S.; Bukhari, S.I.; Alshehri, S.; Rashid, S.M.; Mir, T.M.; Rehman, M.U. Milk-Compositional Study of Metabolites and Pathogens in the Milk of Bovine Animals Affected with Subclinical Mastitis. Molecules 2022, 27, 8631. [Google Scholar] [CrossRef] [PubMed]
- Rocchetti, G.; Becchi, P.P.; Salis, L.; Lucini, L.; Cabiddu, A. Impact of Pasture-Based Diets on the Untargeted Metabolomics Profile of Sarda Sheep Milk. Foods 2022, 12, 143. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Hernández, S.; Esteban-Muñoz, A.; Giménez-Martínez, R.; Aguilar-Cordero, M.J.; Miralles-Buraglia, B.; Olalla-Herrera, M.A. Comparison of Changes in the Fatty Acid Profile of Human Milk of Spanish Lactating Women during the First Month of Lactation Using Gas Chromatography-Mass Spectrometry. A Comparison with Infant Formulas. Nutrients 2019, 11, 3055. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Chen, M.; Luo, R.; Huang, G.; Wu, X.; Zheng, N.; Zhang, Y.; Wang, J. Fatty acid profiles of milk from Holstein cows, Jersey cows, buffalos, yaks, humans, goats, camels, and donkeys based on gas chromatography-mass spectrometry. J. Dairy Sci. 2022, 105, 1687–1700. [Google Scholar] [CrossRef]
- Han, X.X.; Zhang, C.B.; Wang, C.R.; Huang, Y.C.; Liu, Z.Q. Gadolinium inhibits cadmium transport by blocking non-selective cation channels in rice seedling. Ecotox. Environ. Safe. 2019, 179, 160–166. [Google Scholar] [CrossRef]
- Xu, L.P.; Chen, W.J.; Wang, X.Y.; Yu, Z.B.; Han, S.P. Comparative Lipidomic Analyses Reveal Different Protections in Preterm and Term Breast Milk for Infants. Front. Pediatr. 2020, 8, 590. [Google Scholar] [CrossRef]
- Chen, W.J.; Chen, X.H.; Qian, Y.; Wang, X.Y.; Zhou, Y.H.; Yan, X.Y.; Yu, B.S.; Yao, S.W.; Yu, Z.B.; Zhu, J.A.; et al. Lipidomic Profiling of Human Milk Derived Exosomes and Their Emerging Roles in the Prevention of Necrotizing Enterocolitis. Mol. Nutr. Food Res. 2021, 65, e2000845. [Google Scholar] [CrossRef]
- Wang, C.C.; Tong, Y.; Wen, Y.K.; Cai, J.; Guo, H.; Huang, L.F.; Xu, M.; Feng, M.X.; Chen, X.S.; Zhang, J.J.; et al. Hepatocellular Carcinoma-Associated Protein TD26 Interacts and Enhances Sterol Regulatory Element-Binding Protein 1 Activity to Promote Tumor Cell Proliferation and Growth. Hepatology 2018, 68, 1833–1850. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.W.; Wang, Z.; Zhang, K.L.; Chi, Z.X.; Xu, T.; Jiang, D.L.; Chen, S.; Li, W.X.; Yang, X.Y.; Zhang, X.; et al. One-Carbon Metabolism Supports S-Adenosylmethionine and Histone Methylation to Drive Inflammatory Macrophages. Mol. Cell 2019, 75, 1147–1160. [Google Scholar] [CrossRef] [PubMed]
- Playford, R.J.; Weiser, M.J. Bovine Colostrum: Its Constituents and Uses. Nutrients 2021, 13, 265. [Google Scholar] [CrossRef] [PubMed]
- Delplanque, B.; Gibson, R.; Koletzko, B.; Lapillonne, A.; Strandvik, B. Lipid quality in infant nutrition: Current knowledge and future opportunities. J. Pediatr. Gastr. Nutr. 2015, 61, 8–17. [Google Scholar] [CrossRef]
- Contarini, G.; Povolo, M.; Pelizzola, V.; Monti, L.; Bruni, A.; Passolungo, L.; Abeni, F.; Degano, L. Bovine colostrum: Changes in lipid constituents in the first 5 days after parturition. J. Dairy Sci. 2014, 97, 5065–5072. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, S.; Spener, F. Conjugated linoleic acids as functional food: An insight into their health benefits. Nutr. Metab. 2009, 6, 36. [Google Scholar] [CrossRef]
- Hammad, S.; Pu, S.; Jones, P.J. Current Evidence Supporting the Link Between Dietary Fatty Acids and Cardiovascular Disease. Lipids 2016, 51, 507–517. [Google Scholar] [CrossRef]
- Khalid, W.; Gill, P.; Arshad, M.S.; Ali, A.; Ranjha, M.M.A.N.; Mukhtar, S.; Afzal, F.; Maqbool, Z. Functional behavior of DHA and EPA in the formation of babies brain at different stages of age, and protect from different brain-related diseases. Int. J. Food Prop. 2022, 25, 1021–1044. [Google Scholar] [CrossRef]
- Zaloga, G.P. Narrative review of n-3 polyunsaturated fatty acid supplementation upon immune functions, resolution molecules and lipid peroxidation. Nutrients 2021, 13, 662. [Google Scholar] [CrossRef]
- Lukas, S.; Georg, H. Monounsaturated fatty acids, olive oil and health status: A systematic review and meta-analysis of cohort studies. Lipids Health Dis. 2014, 13, 154. [Google Scholar] [CrossRef]
- Wilms, J.N.; Hare, K.S.; Fischer-Tlustos, A.J.; Vahmani, P.; Dugan, M.E.R.; Leal, L.N.; Steele, M.A. Fatty acid profile characterization in colostrum, transition milk, and mature milk of primi- and multiparous cows during the first week of lactation. J. Dairy Sci. 2022, 105, 2612–2630. [Google Scholar] [CrossRef] [PubMed]
- Li, M.H.; Li, W.X.; Kong, F.H.; Kang, S.M.; Liang, X.N.; Han, H.J.; Wu, J.R.; Zheng, Y.; Li, Q.L.; Yue, X.Q.; et al. Metabolomics methods to analyze full spectrum of amino acids in different domains of bovine colostrum and mature milk. Eur. Food Res. Technol. 2020, 246, 213–224. [Google Scholar] [CrossRef]
- Tabatabaie, L.; Klomp, L.W.; Berger, R. L-Serine synthesis in the central nervous system: A review on serine deficiency disorders. Mol. Genet. Metab. 2010, 99, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Stallcup, M.R. Minireview: Protein arginine methylation of nonhistone proteins in transcriptional regulation. Mol. Endocrinol. 2009, 23, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Müller-Tidow, C.; Wang, W.; Idos, G.E.; Diederichs, S.; Yang, R.; Readhead, C.; Berdel, W.E.; Serve, H.; Saville, M.; Watson, R.; et al. Cyclin A1 directly interacts with B-myb and cyclin A1/cdk2 phosphorylate B-myb at functionally important serine and threonine residues: Tissue-specific regulation of B-myb function. Blood 2001, 97, 2091–2097. [Google Scholar] [CrossRef]
- Tochitani, S. Taurine: A Maternally Derived Nutrient Linking Mother and Offspring. Metabolites 2022, 12, 228. [Google Scholar] [CrossRef] [PubMed]
- Buchman, A. Choline deficiency during parenteral nutrition in humans. Nutr. Clin. Pract. 2003, 18, 353–358. [Google Scholar] [CrossRef]
- Yao, Y.P.; Zhao, G.Z.; Xiang, J.Y.; Zou, X.Q.; Jin, Q.Z.; Wang, X.G. Lipid composition and structural characteristics of bovine, caprine and human milk fat globules. Int. Dairy J. 2016, 56, 64–73. [Google Scholar] [CrossRef]
- Tan, S.T.; Ramesh, T.; Toh, X.R.; Nguyen, L.N. Emerging roles of lysophospholipids in health and disease. Prog. Lipid Res. 2020, 80, 101068. [Google Scholar] [CrossRef] [PubMed]
- Anto, L.; Warykas, S.W.; Torres-gonzalez, M.; Blesso, C.N. Milk Polar Lipids: Underappreciated Lipids with Emerging Health Benefits. Nutrients 2020, 12, 1001. [Google Scholar] [CrossRef]
- Sarbu, M.; Zamfir, A.D. Modern separation techniques coupled to high performance mass spectrometry for glycolipid analysis. Electrophoresis 2018, 39, 1155–1170. [Google Scholar] [CrossRef]
- Alexandropoulou, I.; Grammatikopoulou, M.G.; Gkouskou, K.K.; Pritsa, A.A.; Vassilakou, T.; Rigopoulou, E.; Lindqvist, H.M.; Bogdanos, D.P. Ceramides in Autoimmune Rheumatic Diseases: Existing Evidence and Therapeutic Considerations for Diet as an Anticeramide Treatment. Nutrients 2023, 15, 229. [Google Scholar] [CrossRef]
- Jouan, P.N.; Pouliot, Y.; Gauthier, S.F.; Laforest, J.P. Hormones in bovine milk and milk products A survey. Int. Dairy J. 2006, 16, 1408–1414. [Google Scholar] [CrossRef]
- Yong, J.K. Pivotal roles of prolactin and other hormones in lactogenesis and the nutritional composition of human milk. Clin. Exp. Immunol. 2020, 63, 312–313. [Google Scholar] [CrossRef]
- Adgent, M.A.; Umbach, D.M.; Zemel, B.S.; Kelly, A.; Schall, J.I.; Ford, E.G.; James, K.; Darge, K.; Botelho, J.C.; Hubert, W.; et al. A longitudinal study of estrogen-responsive tissues and hormone concentrations in infants fed soy formula. J. Endocrinol. Metab. 2018, 103, 1899–1909. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Yu, M.M.; Zhou, C.J.; Qi, H.; Wen, X.P.; Hou, X.M.; Li, M.; Gao, X.J. FABP5 is a critical regulator of methionine- and estrogen-induced SREBP-1c gene expression in bovine mammary epithelial cells. J. Cell Physiol. 2018, 234, 537–549. [Google Scholar] [CrossRef]
- Martyniak, A.; Tomasik, P.J. A New Perspective on the Renin-Angiotensin System. Diagnostics 2022, 13, 16. [Google Scholar] [CrossRef]
- Shahid, M.; Rehman, K.; Akash, M.S.H.; Suhail, S.; Kamal, S.; Imran, M.; Assiri, M.A. Genetic Polymorphism in Angiotensinogen and Its Association with Cardiometabolic Diseases. Metabolites 2022, 12, 1291. [Google Scholar] [CrossRef] [PubMed]
- Garimella, P.S.; Toit, C.D.; Le, N.N.; Padmanabhan, S. A genomic deep field view of hypertension. Kidney Int. 2023, 103, 42–52. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Absolute Content (Mean ± SD, g/L) | Relative Content (Mean ± SD, %) | ||
|---|---|---|---|---|
| Colostrum | Mature Milk | Colostrum | Mature Milk | |
| C4:0 | 0.009 ± 0.005 * | 0.004 ± 0.001 | 0.015 ± 0.008 | 0.008 ± 0.001 |
| C6:0 | 0.005 ± 0.001 | 0.004 ± 0.001 | 0.008 ± 0.002 | 0.007 ± 0.002 |
| C8:0 | 0.011 ± 0.004 | 0.010 ± 0.003 | 0.02 ± 0.01 | 0.02 ± 0.01 |
| C10:0 | 0.13 ± 0.05 * | 0.53 ± 0.27 | 0.22 ± 0.08 * | 1.04 ± 0.22 |
| C11:0 | 0.003 ± 0.001 * | 0.008 ± 0.003 | 0.005 ± 0.002 * | 0.015 ± 0.005 |
| C12:0 | 0.66 ± 0.16 * | 1.38 ± 0.28 | 1.06 ± 0.24 * | 2.74 ± 0.51 |
| C13:0 | 0.04 ± 0.01 | 0.05 ± 0.01 | 0.07 ± 0.02 | 0.09 ± 0.02 |
| C14:0 | 5.33 ± 0.51 | 5.88 ± 0.50 | 8.64 ± 0.65 * | 11.66 ± 0.78 |
| C14:1n5 | 0.16 ± 0.04 | 0.18 ± 0.03 | 0.27 ± 0.06 * | 0.36 ± 0.05 |
| C15:0 | 1.27 ± 0.21 | 1.21 ± 0.12 | 2.06 ± 0.33 | 2.39 ± 0.19 |
| C15:1n5 | 0.08 ± 0.04 | 0.04 ± 0.01 | 0.13 ± 0.07 | 0.09 ± 0.02 |
| C16:0 | 15.79 ± 2.26 | 13.97 ± 1.92 | 25.60 ± 3.45 | 27.73 ± 3.59 |
| C16:1n7 | 1.19 ± 0.27 * | 1.62 ± 0.27 | 1.92 ± 0.40 * | 3.23 ± 0.61 |
| C17:0 | 0.82 ± 0.08 * | 1.00 ± 0.02 | 1.33 ± 0.15 * | 1.98 ± 0.07 |
| C17:1n7 | 0.39 ± 0.09 | 0.42 ± 0.09 | 0.63 ± 0.13 | 0.84 ± 0.20 |
| C18:0 | 11.26 ± 1.32 * | 7.05 ± 0.53 | 18.32 ± 2.44 * | 14.00 ± 0.93 |
| t−C18:1n9 | 4.80 ± 1.03 * | 1.41 ± 0.32 | 7.77 ± 1.53 * | 2.82 ± 0.70 |
| C18:1n9 | 14.85 ±1.51 * | 12.21 ± 0.36 | 24.08 ± 2.21 | 24.28 ± 1.20 |
| t,t−C18:2n6 | 0.137 ± 0.024 * | 0.036 ± 0.004 | 0.22 ± 0.04 * | 0.07 ± 0.01 |
| C18:2n6 (LA) | 2.03 ± 0.43 * | 1.32 ± 0.20 | 3.29 ± 0.67 | 2.63 ± 0.44 |
| C18:3n6 | 0.013 ± 0.002 * | 0.008 ± 0.002 | 0.021 ± 0.004 | 0.017 ± 0.003 |
| C18:3n3 (ALA) | 1.36 ± 0.27 * | 0.67 ± 0.11 | 2.21 ± 0.45 * | 1.34 ± 0.23 |
| C20:0 | 0.43 ± 0.08 | 0.42 ± 0.04 | 0.70 ± 0.14 | 0.83 ± 0.06 |
| C20:1n9 | 0.08 ± 0.01 | 0.09 ± 0.02 | 0.12 ± 0.02 * | 0.18 ± 0.04 |
| C20:2n6 | 0.024 ± 0.004 * | 0.020 ± 0.003 | 0.039 ± 0.01 | 0.040 ± 0.04 |
| C21:0 | 0.07 ± 0.01 | 0.08 ± 0.02 | 0.11 ± 0.03 | 0.16 ± 0.03 |
| C20:3n6 | 0.012 ± 0.003 | 0.017 ± 0.003 | 0.02 ± 0.01 * | 0.03 ± 0.01 |
| C20:4n6 (ARA) | 0.10 ± 0.03 * | 0.17 ± 0.04 | 0.16 ± 0.05 * | 0.34 ± 0.07 |
| C20:3n3 | 0.02 ± 0.003 | 0.02 ± 0.003 | 0.03 ± 0.01 | 0.03 ± 0.01 |
| C22:0 | 0.14 ± 0.03 | 0.15 ± 0.03 | 0.22 ± 0.05 | 0.30 ± 0.04 |
| C20:5n3 (EPA) | 0.06 ± 0.01 | 0.06 ± 0.01 | 0.09 ± 0.02 | 0.11 ± 0.02 |
| C22:1n9 | 0.025 ± 0.005 | 0.023 ± 0.004 | 0.041 ± 0.01 | 0.046 ± 0.01 |
| C22:2n6 | 0.003 ± 0.0004 | 0.002 ± 0.004 | 0.004 ± 0.001 | 0.003 ± 0.001 |
| C23:0 | 0.07 ± 0.01 | 0.07 ± 0.01 | 0.11 ± 0.03 | 0.14 ± 0.03 |
| C22:4n6 | 0.013 ± 0.002 | 0.013 ± 0.003 | 0.020 ± 0.003 | 0.026 ± 0.006 |
| C22:5n6 | 0.002 ± 0.001 | 0.002 ± 0.001 | 0.003 ± 0.002 | 0.005 ± 0.002 |
| C24:0 | 0.07 ± 0.01 | 0.07 ± 0.01 | 0.11 ± 0.02 | 0.13 ± 0.02 |
| C22:5n3 | 0.16 ± 0.03 | 0.11 ± 0.03 | 0.26 ± 0.05 | 0.22 ± 0.06 |
| C24:1n9 | 0.008 ± 0.002 | 0.010 ± 0.004 | 0.014 ± 0.003 | 0.021 ± 0.007 |
| C22:6n3 (DHA) | 0.04 ± 0.01 * | 0.02 ± 0.01 | 0.06 ± 0.01 | 0.04 ± 0.02 |
| ΣTFAs | 61.64 ± 2.36 * | 50.35 ± 1.33 | - | - |
| ΣSFAs | 36.10 ± 2.79 * | 31.87 ± 2.31 | 58.61 ± 4.35 | 63.23 ± 3.36 |
| ΣUFAs | 25.53 ± 3.02 * | 18.48 ± 1.34 | 41.40 ± 4.35 | 36.77 ± 3.36 |
| ΣMUFAs | 21.57 ± 2.38 * | 16.01 ± 1.00 | 34.97 ± 3.31 | 31.85 ± 2.65 |
| ΣPUFAs | 3.96 ± 0.77 * | 2.47 ± 0.37 | 6.43 ± 1.23 * | 4.91 ± 0.79 |
| Σn−3PUFAs | 2.34 ± 0.49 * | 1.59 ± 0.25 | 2.64 ± 0.49 * | 1.74 ± 0.27 |
| Σn−6PUFAs | 1.63 ± 0.29 * | 0.88 ± 0.12 | 3.79 ± 0.77 | 3.17 ± 0.52 |
| Σn−6/n−3PUFAs | 1.43 ± 0.11 * | 1.82 ± 0.06 | 1.43 ± 0.11 * | 1.82 ± 0.06 |
| ΣPUFAs/ΣSFAs | 0.11 ± 0.03 * | 0.08 ± 0.02 | 0.11 ± 0.03 * | 0.08 ± 0.02 |
| Compound | Colostrum (Mean ± SD) | Mature Milk (Mean ± SD) |
|---|---|---|
| Lysine (g/L) | 8.44 ± 0.65 * | 4.17 ± 0.21 |
| Tryptophan (g/L) | 1.87 ± 0.33 * | 0.81 ± 0.06 |
| Phenylalanine (g/L) | 4.53 ± 0.77 * | 2.27 ± 0.23 |
| Methionine (g/L) | 2.07 ± 0.29 * | 1.36 ± 0.20 |
| Threonine (g/L) | 3.24 ± 0.28 * | 2.29 ± 0.30 |
| Leucine (g/L) | 6.56 ± 0.67 * | 5.06 ± 0.36 |
| Isoleucine (g/L) | 3.49 ± 0.55 * | 2.34 ± 0.27 |
| Valine (g/L) | 3.26 ± 0.51 * | 2.56 ± 0.22 |
| Histidine (g/L) | 3.00 ± 0.37 * | 1.77 ± 0.17 |
| Glutamic acid (g/L) | 19.69 ± 2.33 * | 11.03 ± 0.84 |
| Glycine (g/L) | 2.70 ± 0.51 | 2.77 ± 0.16 |
| Aspartic acid (g/L) | 4.64 ± 0.50 | 4.12 ± 0.34 |
| Arginine (g/L) | 2.19 ± 0.40 * | 1.48 ± 0.28 |
| Serine (g/L) | 3.61 ± 0.36 * | 2.50 ± 0.39 |
| Tyrosine (g/L) | 1.93 ± 0.21 | 1.61 ± 0.20 |
| Proline (g/L) | 2.88 ± 0.67 | 2.18 ± 0.26 |
| Alanine (g/L) | 2.17 ± 0.29 | 1.83 ± 0.45 |
| Cysteine (g/L) | 1.35 ± 0.28 * | 0.61 ± 0.08 |
| Cystine (g/L) | 0.39 ± 0.09 | 0.31 ± 0.06 |
| Ornithine (mg/L) | 6.50 ± 0.90 | 5.93 ± 0.61 |
| Taurine (mg/L) | 53.77 ± 5.53 * | 22.62 ± 4.10 |
| Choline (mg/L) | 766.82 ± 39.43 * | 220.54 ± 20.83 |
| Aminoadipic acid (mg/L) | 2.10 ± 0.43 * | 10.77 ± 1.42 |
| Hydroxyproline (mg/L) | 3.27 ± 0.71 * | 8.56 ± 1.13 |
| Spermidine (mg/L) | 9.71 ± 1.47 | 9.72 ± 0.86 |
| EAAs (g/L) | 36.44 ± 1.38 * | 22.63 ± 0.92 |
| NEAAs (g/L) | 41.56 ± 2.15 * | 28.42 ± 1.11 |
| TAAs (g/L) | 78.00 ± 3.16 * | 50.89 ± 2.26 |
| EAAs/NEAAs | 0.88 ± 0.04 * | 0.79 ± 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, L.; Pei, J.; Bao, P.; Wang, X.; Guo, S.; Cao, M.; Kang, Y.; Yan, P.; Guo, X. The Study of Yak Colostrum Nutritional Content Based on Foodomics. Foods 2023, 12, 1707. https://doi.org/10.3390/foods12081707
Xiong L, Pei J, Bao P, Wang X, Guo S, Cao M, Kang Y, Yan P, Guo X. The Study of Yak Colostrum Nutritional Content Based on Foodomics. Foods. 2023; 12(8):1707. https://doi.org/10.3390/foods12081707
Chicago/Turabian StyleXiong, Lin, Jie Pei, Pengjia Bao, Xingdong Wang, Shaoke Guo, Mengli Cao, Yandong Kang, Ping Yan, and Xian Guo. 2023. "The Study of Yak Colostrum Nutritional Content Based on Foodomics" Foods 12, no. 8: 1707. https://doi.org/10.3390/foods12081707
APA StyleXiong, L., Pei, J., Bao, P., Wang, X., Guo, S., Cao, M., Kang, Y., Yan, P., & Guo, X. (2023). The Study of Yak Colostrum Nutritional Content Based on Foodomics. Foods, 12(8), 1707. https://doi.org/10.3390/foods12081707