
Transcriptome Reveals Regulation of Quorum Sensing of Hafnia alvei H4 on the Coculture System of Hafnia alvei H4 and Pseudomonas fluorescens ATCC13525



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Crystal Violet Biofilm Assay
2.3. Extracellular Polysaccharides Assay
2.4. Biogenic Amine Measurement Using HPLC
2.5. Transcriptome Analysis and Quantitative Reverse Transcription (RT-qPCR)
2.6. Statistical Analysis
2.7. Accession Numbers
3. Results and Discussion
3.1. Characteristics under Coculture Conditions
3.1.1. Biofilm Formation
3.1.2. Exopolysaccharide Assay
3.1.3. Biogenic Amine Production
3.2. Transcriptome Analysis under Coculture Conditions
3.2.1. Transcriptional Responses in H. alvei H4 and P. fluorescens ATCC13525
3.2.2. Analysis of Genes Associated with Biofilm Formation
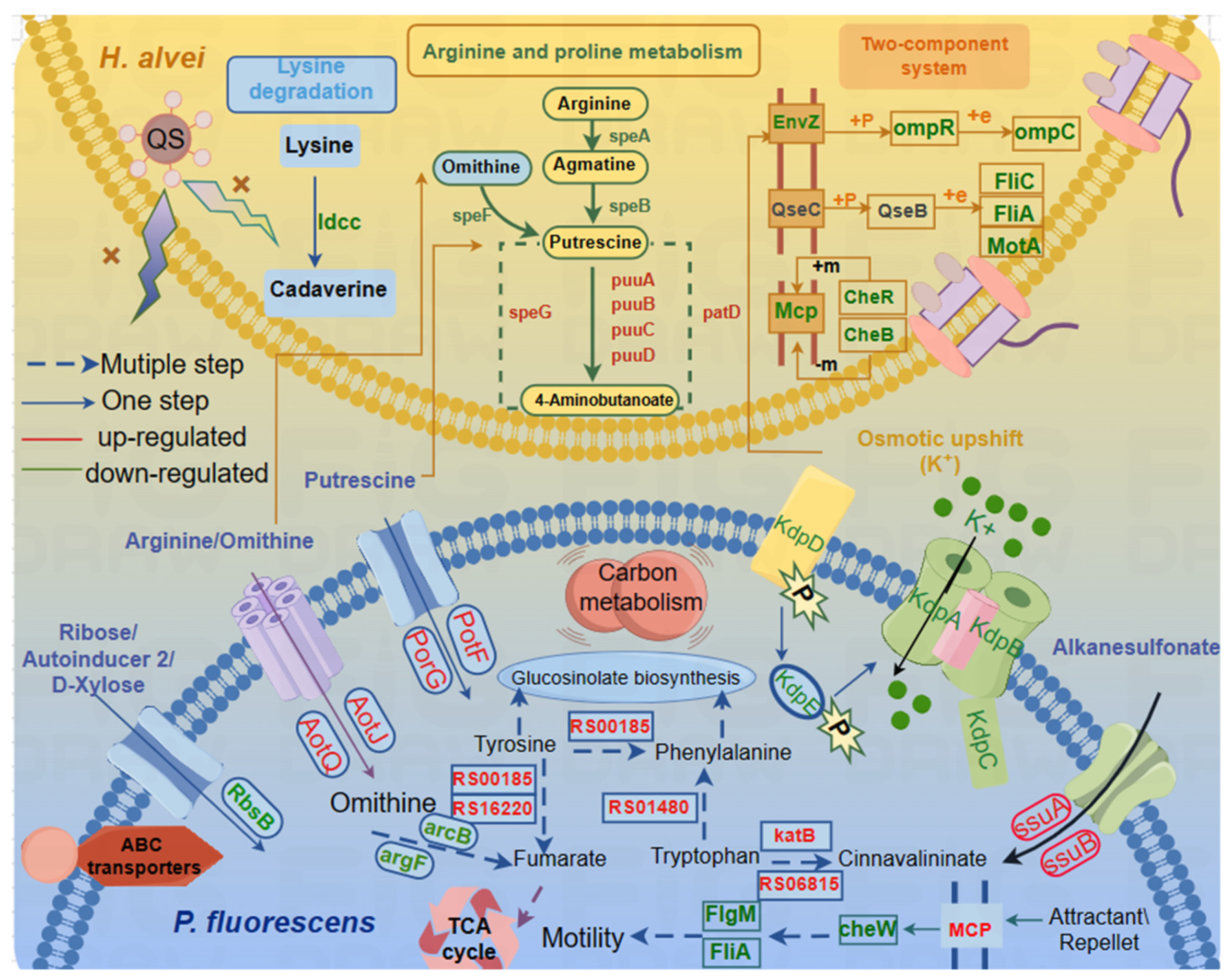
Flagellar Assembly and Bacterial Chemotaxis
Signal Transduction System
3.2.3. Amino Acid Metabolism
3.3. Real-Time Quantitative PCR Validation (RT-qPCR)
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Özogul, F.; Polat, A.; Özogul, Y. The effects of modified atmosphere packaging and vacuum packaging on chemical, sensory and microbiological changes of sardines (Sardina pilchardus). Food Chem. 2004, 85, 49–57. [Google Scholar] [CrossRef]
- Wang, Y.; Feng, L.; Lu, H.; Zhu, J.; Kumar, V.; Liu, X. Transcriptomic analysis of the food spoilers Pseudomonas fluorescens reveals the antibiofilm of carvacrol by interference with intracellular signaling processes. Food Control 2021, 127, 108115. [Google Scholar] [CrossRef]
- Robson, A.A.; Kelly, M.S.; Latchford, J.W. Effect of temperature on the spoilage rate of whole, unprocessed crabs: Carcinus maenas, Necora puber and Cancer pagurus. Food Microbiol. 2007, 24, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.F.; Lin, H.; Sui, J.X.; Cao, L.M. Production and Characterization of Egg Yolk Antibodies (Igy) against Two Specific Spoilage Organisms (SSO) in Aquatic Products. Adv. Mater. Res. 2011, 343–344, 519–529. [Google Scholar] [CrossRef]
- Han, F.; Li, M.; Lin, H.; Wang, J.; Cao, L.; Khan, M.N. The novel Shewanella putrefaciens-infecting bacteriophage Spp001: Genome sequence and lytic enzymes. J. Ind. Microbiol. Biotechnol. 2014, 41, 1017–1026. [Google Scholar] [CrossRef]
- Bruhn, J.B.; Christensen, A.B.; Flodgaard, L.R.; Nielsen, K.F.; Larsen, T.O.; Givskov, M.; Gram, L. Presence of Acylated Homoserine Lactones (AHLs) and AHL-Producing Bacteria in Meat and Potential Role of AHL in Spoilage of Meat. Appl. Environ. Microbiol. 2004, 70, 4293–4302. [Google Scholar] [CrossRef]
- Zhu, Y.L.; Hou, H.M.; Zhang, G.L.; Wang, Y.F.; Hao, H.S. AHLs Regulate Biofilm Formation and Swimming Motility of Hafnia alvei H4. Front. Microbiol. 2019, 10, 1330. [Google Scholar] [CrossRef]
- Shen, Y.; Cui, F.C.; Wang, D.F.; Li, T.T.; Li, J.R. Quorum Quenching Enzyme (PF-1240) Capable to Degrade AHLs as a Candidate for Inhibiting Quorum Sensing in Food Spoilage Bacterium Hafnia alvei. Foods 2021, 10, 2700. [Google Scholar] [CrossRef]
- Hwang, C.C.; Lee, Y.C.; Huang, Y.R.; Lin, C.M.; Shiau, C.Y.; Hwang, D.F.; Tsai, Y.H. Biogenic amines content, histamine-forming bacteria and adulteration of bonito in tuna candy products. Food Control 2010, 21, 845–850. [Google Scholar] [CrossRef]
- Kanki, M.; Yoda, T.; Ishibashi, M.; Tsukamoto, T. Photobacterium phosphoreum caused a histamine fish poisoning incident. Int. J. Food Microbiol. 2004, 92, 79–87. [Google Scholar] [CrossRef]
- Ge, Y.; Zhu, J.; Ye, X.; Yang, Y. Spoilage potential characterization of Shewanella and Pseudomonas isolated from spoiled large yellow croaker (Pseudosciaena crocea). Lett. Appl. Microbiol. 2017, 64, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cai, L.; Li, Y.; Xu, X.; Zhou, G. Biofilm formation by meat-borne Pseudomonas fluorescens on stainless steel and its resistance to disinfectants. Food Control 2018, 91, 397–403. [Google Scholar] [CrossRef]
- Zarei, M.; Yousefvand, A.; Maktabi, S.; Pourmahdi Borujeni, M.; Mohammadpour, H. Identification, phylogenetic characterisation and proteolytic activity quantification of high biofilm-forming Pseudomonas fluorescens group bacterial strains isolated from cold raw milk. Int. Dairy J. 2020, 109, 104787. [Google Scholar] [CrossRef]
- Mann, E.E.; Wozniak, D.J. Pseudomonas biofilm matrix composition and niche biology. FEMS Microbiol. Rev. 2012, 36, 893–916. [Google Scholar] [CrossRef] [PubMed]
- Delille, A.; Quilès, F.; Humbert, F. In Situ Monitoring of the Nascent Pseudomonas fluorescens Biofilm Response to Variations in the Dissolved Organic Carbon Level in Low-Nutrient Water by Attenuated Total Reflectance-Fourier Transform Infrared Spectroscopy. Appl. Environ. Microbiol. 2007, 73, 5782–5788. [Google Scholar] [CrossRef]
- Puga, C.H.; Orgaz, B.; SanJose, C. Listeria monocytogenes Impact on Mature or Old Pseudomonas fluorescens Biofilms during Growth at 4 and 20 °C. Front. Microbiol. 2016, 7, 00134. [Google Scholar] [CrossRef]
- Li, T.; Yang, B.; Li, X.; Li, J.; Zhao, G.; Kan, J. Quorum sensing system and influence on food spoilage in Pseudomonas fluorescens from turbot. J. Food Sci. Technol. 2018, 55, 3016–3025. [Google Scholar] [CrossRef]
- Chen, T.R.; Wei, Q.K.; Chen, Y.J. Pseudomonas spp and Hafnia alvei growth in UHT milk at cold storage. Food Control 2011, 22, 697–701. [Google Scholar] [CrossRef]
- Liu, M.; Gray, J.M.; Griffiths, M.W. Occurrence of Proteolytic Activity and N-Acyl-Homoserine Lactone Signals in the Spoilage of Aerobically Chill-Stored Proteinaceous Raw Foods. J. Food Prot. 2006, 69, 2729–2737. [Google Scholar] [CrossRef]
- Nychas, G.-J.E.; Skandamis, P.N.; Tassou, C.C.; Koutsoumanis, K.P. Meat spoilage during distribution. Meat Sci. 2008, 78, 77–89. [Google Scholar] [CrossRef]
- Fu, L.; Wang, C.; Liu, N.; Ma, A.; Wang, Y. Quorum sensing system-regulated genes affect the spoilage potential of Shewanella baltica. Food Res. Int. 2018, 107, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Whiteley, M.; Diggle, S.P.; Greenberg, E.P. Progress in and promise of bacterial quorum sensing research. Nature 2017, 551, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, J.R.; Lee, K.; Richard, B.; Walker, A.R.; Burne, R.A. Direct interactions with commensal streptococci modify intercellular communication behaviors of Streptococcus mutans. ISME J. 2020, 15, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Sikdar, R.; Elias, M. Quorum quenching enzymes and their effects on virulence, biofilm, and microbiomes: A review of recent advances. Expert Rev. Anti-Infect. Ther. 2020, 18, 1221–1233. [Google Scholar] [CrossRef]
- Hou, H.M.; Zhu, Y.L.; Wang, J.Y.; Jiang, F.; Qu, W.Y.; Zhang, G.L.; Hao, H.-S. Characteristics of N-Acylhomoserine Lactones Produced by Hafnia alvei H4 Isolated from Spoiled Instant Sea Cucumber. Sensors 2017, 17, 772. [Google Scholar] [CrossRef]
- Harimawan, A.; Ting, Y.-P. Investigation of extracellular polymeric substances (EPS) properties of P. aeruginosa and B. subtilis and their role in bacterial adhesion. Colloids Surf. B: Biointerfaces 2016, 146, 459–467. [Google Scholar] [CrossRef]
- Sang, X.; Ma, X.; Hao, H.; Bi, J.; Zhang, G.; Hou, H. Evaluation of biogenic amines and microbial composition in the Chinese traditional fermented food grasshopper sub shrimp paste. Lwt 2020, 134, 109979. [Google Scholar] [CrossRef]
- Xue, Z.P.; Cu, X.; Xu, K.; Peng, J.H.; Liu, H.R.; Zhao, R.T.; Wang, Z.; Wang, T.; Xu, Z.S. The effect of glutathione biosynthesis of Streptococcus thermophilus ST-1 on cocultured Lactobacillus delbrueckii ssp. bulgaricus ATCC11842. J. Dairy Sci. 2023, 106, 884–896. [Google Scholar] [CrossRef]
- Winkelströter, L.K.; Teixeira, F.B.d.R.; Silva, E.P.; Alves, V.F.; De Martinis, E.C.P. Unraveling Microbial Biofilms of Importance for Food Microbiology. Microb. Ecol. 2013, 68, 35–46. [Google Scholar] [CrossRef]
- Jahid, I.K.; Ha, S.D. The Paradox of Mixed-Species Biofilms in the Context of Food Safety. Compr. Rev. Food Sci. Food Saf. 2014, 13, 990–1011. [Google Scholar] [CrossRef]
- Zhang, C.; Zhu, F.; Jatt, A.N.; Liu, H.; Niu, L.; Zhang, L.; Liu, Y. Characterization of co-culture of Aeromonas and Pseudomonas bacterial biofilm and spoilage potential on refrigerated grass carp (Ctenopharyngodon idellus). Lett. Appl. Microbiol. 2020, 71, 337–344. [Google Scholar] [PubMed]
- Linares, D.M.; Martín, M.; Ladero, V.; Alvarez, M.A.; Fernández, M. Biogenic Amines in Dairy Products. Crit. Rev. Food Sci. Nutr. 2011, 51, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Rossi, P.; Xing, Q.; Bini, E.; Portaliou, A.G.; Clay, M.C.; Warren, E.M.; Khanra, N.K.; Economou, A.; Kalodimos, C.G. Chaperone Recycling in Late-Stage Flagellar Assembly. J. Mol. Biol. 2023, 435, 167954. [Google Scholar] [CrossRef] [PubMed]
- Saijo-Hamano, Y.; Uchida, N.; Namba, K.; Oosawa, K. In Vitro Characterization of FlgB, FlgC, FlgF, FlgG, and FliE, Flagellar Basal Body Proteins of Salmonella. J. Mol. Biol. 2004, 339, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Altarriba, M.; Merino, S.; Gavín, R.; Canals, R.o.; Rabaan, A.; Shaw, J.G.; Tomás, J.M. A polar flagella operon (flg) of Aeromonas hydrophila contains genes required for lateral flagella expression. Microb. Pathog. 2003, 34, 249–259. [Google Scholar] [CrossRef]
- Deme, J.C.; Johnson, S.; Vickery, O.; Aron, A.; Monkhouse, H.; Griffiths, T.; James, R.H.; Berks, B.C.; Coulton, J.W.; Stansfeld, P.J.; et al. Structures of the stator complex that drives rotation of the bacterial flagellum. Nat. Microbiol. 2020, 5, 1553–1564. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.A.; Chacko, S.J.; Vénien-Bryan, C.; Wadhams, G.H.; Armitage, J.P. Regulation of Flagellum Number by FliA and FlgM and Role in Biofilm Formation by Rhodobacter sphaeroides. J. Bacteriol. 2011, 193, 4010–4014. [Google Scholar] [CrossRef]
- Hazelbauer, G.L. Bacterial Chemotaxis: The Early Years of Molecular Studies. Annu. Rev. Microbiol. 2012, 66, 285–303. [Google Scholar] [CrossRef]
- Karmakar, R. State of the art of bacterial chemotaxis. J. Basic Microbiol. 2021, 61, 366–379. [Google Scholar] [CrossRef]
- Porter, S.L.; Wadhams, G.H.; Armitage, J.P. Signal processing in complex chemotaxis pathways. Nat. Rev. Microbiol. 2011, 9, 153–165. [Google Scholar] [CrossRef]
- Minamino, T.; Kinoshita, M.; Namba, K. Directional Switching Mechanism of the Bacterial Flagellar Motor. Comput. Struct. Biotechnol. J. 2019, 17, 1075–1081. [Google Scholar] [CrossRef]
- Kentner, D.; Sourjik, V. Spatial organization of the bacterial chemotaxis system. Curr. Opin. Microbiol. 2006, 9, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Y.; Han, S.; Qin, M.; Wu, X.; Niu, W.; Gao, C.; Wang, H. N-acyl-homoserine lactones-mediated quorum sensing promotes intestinal colonization of Aeromonas veronii through facilitating cheA gene expression. Aquaculture 2024, 579, 740189. [Google Scholar] [CrossRef]
- Sieuwerts, S.; Molenaar, D.; van Hijum, S.A.F.T.; Beerthuyzen, M.; Stevens, M.J.A.; Janssen, P.W.M.; Ingham, C.J.; de Bok, F.A.M.; de Vos, W.M.; van Hylckama Vlieg, J.E.T. Mixed-Culture Transcriptome Analysis Reveals the Molecular Basis of Mixed-Culture Growth in Streptococcus thermophilus and Lactobacillus bulgaricus. Appl. Environ. Microbiol. 2010, 76, 7775–7784. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.C.; Morgan, L.K.; Godakumbura, P.; Kenney, L.J.; Anand, G.S. The inner membrane histidine kinase EnvZ senses osmolality via helix-coil transitions in the cytoplasm. EMBO J. 2012, 31, 2648–2659. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Yang, Z.; Xie, J. Comparative Transcriptome Analysis of Shewanella putrefaciens WS13 Biofilms Under Cold Stress. Front. Cell. Infect. Microbiol. 2022, 12, 851521. [Google Scholar] [CrossRef]
- Zhang, B.Z.; Jiang, C.S.; Cao, H.; Zeng, W.; Ren, J.P.; Hu, Y.F.; Li, W.T.; He, Q.G. Transcriptome analysis of heat resistance regulated by quorum sensing system in Glaesserella parasuis. Front. Microbiol. 2022, 13, 968460. [Google Scholar] [CrossRef]
- Xie, Z.; Jian, H.; Jin, Z.; Xiao, X.; Kelly, R.M. Enhancing the Adaptability of the Deep-Sea Bacterium Shewanella piezotolerans WP3 to High Pressure and Low Temperature by Experimental Evolution under H2O2 Stress. Appl. Environ. Microbiol. 2018, 84, e02342-17. [Google Scholar] [CrossRef]
- Zhang, P.Y.; Xu, P.P.; Xia, Z.J.; Wang, J.; Xiong, J.; Li, Y.Z. Combined treatment with the antibiotics kanamycin and streptomycin promotes the conjugation of Escherichia coli. FEMS Microbiol. Lett. 2013, 348, 149–156. [Google Scholar] [CrossRef]
- Armbruster, C.E.; Pang, B.; Murrah, K.; Juneau, R.A.; Perez, A.C.; Weimer, K.E.D.; Swords, W.E. RbsB (NTHI_0632) mediates quorum signal uptake in nontypeable Haemophilus influenzae strain 86-028NP. Mol. Microbiol. 2011, 82, 836–850. [Google Scholar] [CrossRef]
- Li, J.; Zhou, L.; Feng, W.; Cheng, H.; Muhammad, A.I.; Ye, X.; Zhi, Z. Comparison of Biogenic Amines in Chinese Commercial Soy Sauces. Molecules 2019, 24, 1522. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Zhang, Y.; Lin, X.; Zhang, S.; Chen, Y.; Ji, C. Inhibition Mechanism of Lactiplantibacillus plantarum on the Growth and Biogenic Amine Production in Morganella morganii. Foods 2023, 12, 3625. [Google Scholar] [CrossRef] [PubMed]
- Jijakli, K.; Jensen, P.A.; Whiteson, K.L. Metabolic Modeling of Streptococcus mutans Reveals Complex Nutrient Requirements of an Oral Pathogen. mSystems 2019, 4, 419507. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zheng, L.; Lu, Z.; Zhou, L.; Meng, F.; Shi, C.; Bie, X. Biochemical and molecular regulatory mechanism of the pgpH gene on biofilm formation in Listeria monocytogenes. J. Appl. Microbiol. 2023, 134, lxac086. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | log2 Radio | Gene Description |
|---|---|---|
| flagellar assembly | ||
| fliL | −4.32515228 | flagellar basal body-associated protein FliL |
| fliN | −5.766718961 | flagellar motor switch protein FliN |
| fliM | −2.837097495 | flagellar motor switch protein FliM |
| fliJ | −2.408896556 | flagellar export protein FliJ |
| ERL64_RS17280 | −1.339639572 | flagellar hook-length control protein FliK |
| fliS | −2.016760592 | flagellar export chaperone FliS |
| motA | −2.425518042 | flagellar motor stator protein MotA |
| flgB | −3.035717278 | flagellar basal body rod protein FlgB |
| flgC | −3.475398667 | flagellar basal body rod protein FlgC |
| flgD | −3.258609823 | flagellar hook assembly protein FlgD |
| flgE | −3.074676572 | flagellar hook protein FlgE |
| ERL64_RS17360 | −3.436893106 | flagellar basal body rod protein FlgF |
| flgG | −3.336673997 | flagellar basal body rod protein FlgG |
| flgK | −1.888983693 | flagellar hook-associated protein FlgK |
| flgL | −2.226392467 | flagellar hook-associated protein FlgL |
| ERL64_RS17345 | −1.831507544 | flagellar basal body P-ring protein FlgI |
| bacterial chemotaxis | ||
| ERL64_RS14790 | −2.265739966 | methyl-accepting chemotaxis protein |
| Tsr | −2.265395717 | methyl-accepting chemotaxis protein |
| ERL64_RS14800 | −2.424604914 | chemotaxis response regulator protein |
| cheR | −2.098189509 | protein-glutamate O-methyltransferase CheR |
| ERL64_RS14800 | −2.424604914 | chemotaxis response regulator protein-glutamate methylesterase |
| two-component system | ||
| envZ | −0.966750293 | two-component system sensor histidine kinase EnvZ |
| ompR | −0.459688448 | two-component system response regulator OmpR |
| phoR | −1.146705289 | phosphate regulon sensor histidine kinase PhoR |
| phoQ | 0.940798155 | two-component system sensor histidine kinase PhoQ |
| phoP | 1.035332663 | two-component system response regulator PhoP |
| rpoN | −1.164441481 | RNA polymerase factor sigma-54 |
| BA metabolism | ||
| ldcc | −3.212727378 | lysine decarboxylase |
| speF | −0.860138669 | ornithine decarboxylase SpeF |
| speA | −0.82353036 | biosynthetic arginine decarboxylase |
| speB | −0.669157809 | agmatinase |
| puuA | 3.06324221 | glutamine synthetase family protein |
| puuB | 2.140552542 | FAD-binding oxidoreductase |
| puuC | 2.239399155 | aldehyde dehydrogenase PuuC |
| puuD | 2.663951099 | gamma-glutamyl-gamma-aminobutyrate hydrolase |
| patD | 1.429802723 | amino butyraldehyde dehydrogenase |
| ERL64_RS21280 | 3.401803121 | amma-amino butyraldehyde dehydrogenase |
| ERL64_RS02325 | 3.06324221 | glutamine synthetase family protein |
| speG | 0.419542916 | spermidine N1-acetyltransferase |
| potF | −0.987904791 | spermidine/putrescine ABC transporter substrate-binding protein PotF |
| potI | −2.362303979 | putrescine ABC transporter permease PotI |
| potH | −2.568324997 | putrescine ABC transporter permease PotH |
| potG | −0.987904791 | putrescine ABC transporter ATP-binding subunit PotG |
| Gene Name | log2 Radio | Gene Description |
|---|---|---|
| biofilm formation | ||
| fliL | −0.925171985 | flagellar basal body-associated protein FliL |
| flgM | −0.699499627 | flagellar biosynthesis anti-sigma factor FlgM |
| fliA | −0.523949137 | RNA polymerase sigma factor FliA |
| CPH89_RS16845 | −1.029022096 | OmpA family protein |
| cheW | −0.506899146 | chemotaxis protein CheW |
| CPH89_RS23015 | −0.547698862 | methyl-accepting chemotaxis protein |
| tssC | −1.370116898 | type VI secretion system contractile sheath large subunit |
| tssG | 2.543096874 | type VI secretion system baseplate subunit TssG |
| tssH | 0.939045148 | type VI secretion system ATPase TssH |
| CPH89_RS09505 | 1.808730303 | type VI secretion system tip protein VgrG |
| amino acid metabolism | ||
| CPH89_RS22860 | −1.373666561 | methionine gamma-lyase |
| CPH89_RS03900 | −1.088510925 | L-serine ammonia-lyase |
| CPH89_RS10590 | −0.592860749 | homoserine kinase |
| mmsB | −1.524151588 | 3-hydroxyisobutyrate dehydrogenase |
| CPH89_RS06815 | 0.574998597 | catalase |
| katB | 1.644147633 | catalase KatB |
| CPH89_RS08005 | 2.317725314 | D-amino acid dehydrogenase |
| CPH89_RS00185 | 0.475801482 | aspartate/tyrosine/aromatic aminotransferase |
| CPH89_RS21450 | 1.152619749 | aspartate aminotransferase family protein |
| CPH89_RS16850 | 0.716263227 | argininosuccinate synthase |
| CPH89_RS21350 | 0.422007256 | glutamine synthetase family protein |
| gltB | 0.609812179 | glutamate synthase large subunit |
| CPH89_RS00200 | 0.920911912 | aldehyde dehydrogenase family protein |
| CPH89_RS27825 | 1.897595222 | CoA-acylating methylmalonate-semialdehyde dehydrogenase |
| ABC transport | ||
| RbsB | −0.923885631 | sugar ABC transporter substrate-binding protein |
| CPH89_RS28860 | −1.034779256 | ABC transporter ATP-binding protein |
| CPH89_RS10545 | −0.845831876 | MetQ/NlpA family ABC transporter substrate-binding protein |
| CPH89_RS24050 | −2.801510174 | ABC transporter permease |
| CPH89_RS19375 | −1.480685108 | ABC transporter ATP-binding protein |
| CPH89_RS16095 | −1.459658475 | amino acid ABC transporter permease |
| CPH89_RS23690 | 2.876670285 | carbohydrate ABC transporter permease |
| urtD | 2.872150064 | urea ABC transporter ATP-binding protein UrtD |
| CPH89_RS08665 | 1.314199994 | sulfonate ABC transporter substrate-binding protein |
| ssuB | 0.972531529 | aliphatic sulfonates ABC transporter ATP-binding protein |
| CPH89_RS15185 | 0.63533128 | glycine betaine ABC transporter substrate-binding protein |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, X.; Zhang, G.; Bi, J.; Hou, H. Transcriptome Reveals Regulation of Quorum Sensing of Hafnia alvei H4 on the Coculture System of Hafnia alvei H4 and Pseudomonas fluorescens ATCC13525. Foods 2024, 13, 336. https://doi.org/10.3390/foods13020336
Wang Y, Li X, Zhang G, Bi J, Hou H. Transcriptome Reveals Regulation of Quorum Sensing of Hafnia alvei H4 on the Coculture System of Hafnia alvei H4 and Pseudomonas fluorescens ATCC13525. Foods. 2024; 13(2):336. https://doi.org/10.3390/foods13020336
Chicago/Turabian StyleWang, Yanan, Xue Li, Gongliang Zhang, Jingran Bi, and Hongman Hou. 2024. "Transcriptome Reveals Regulation of Quorum Sensing of Hafnia alvei H4 on the Coculture System of Hafnia alvei H4 and Pseudomonas fluorescens ATCC13525" Foods 13, no. 2: 336. https://doi.org/10.3390/foods13020336
APA StyleWang, Y., Li, X., Zhang, G., Bi, J., & Hou, H. (2024). Transcriptome Reveals Regulation of Quorum Sensing of Hafnia alvei H4 on the Coculture System of Hafnia alvei H4 and Pseudomonas fluorescens ATCC13525. Foods, 13(2), 336. https://doi.org/10.3390/foods13020336