
Plant and Arthropod IgE-Binding Papain-like Cysteine Proteases: Multiple Contributions to Allergenicity

 ,
,  ,
, 
Abstract
:1. Introduction
2. PLCP Structural Features
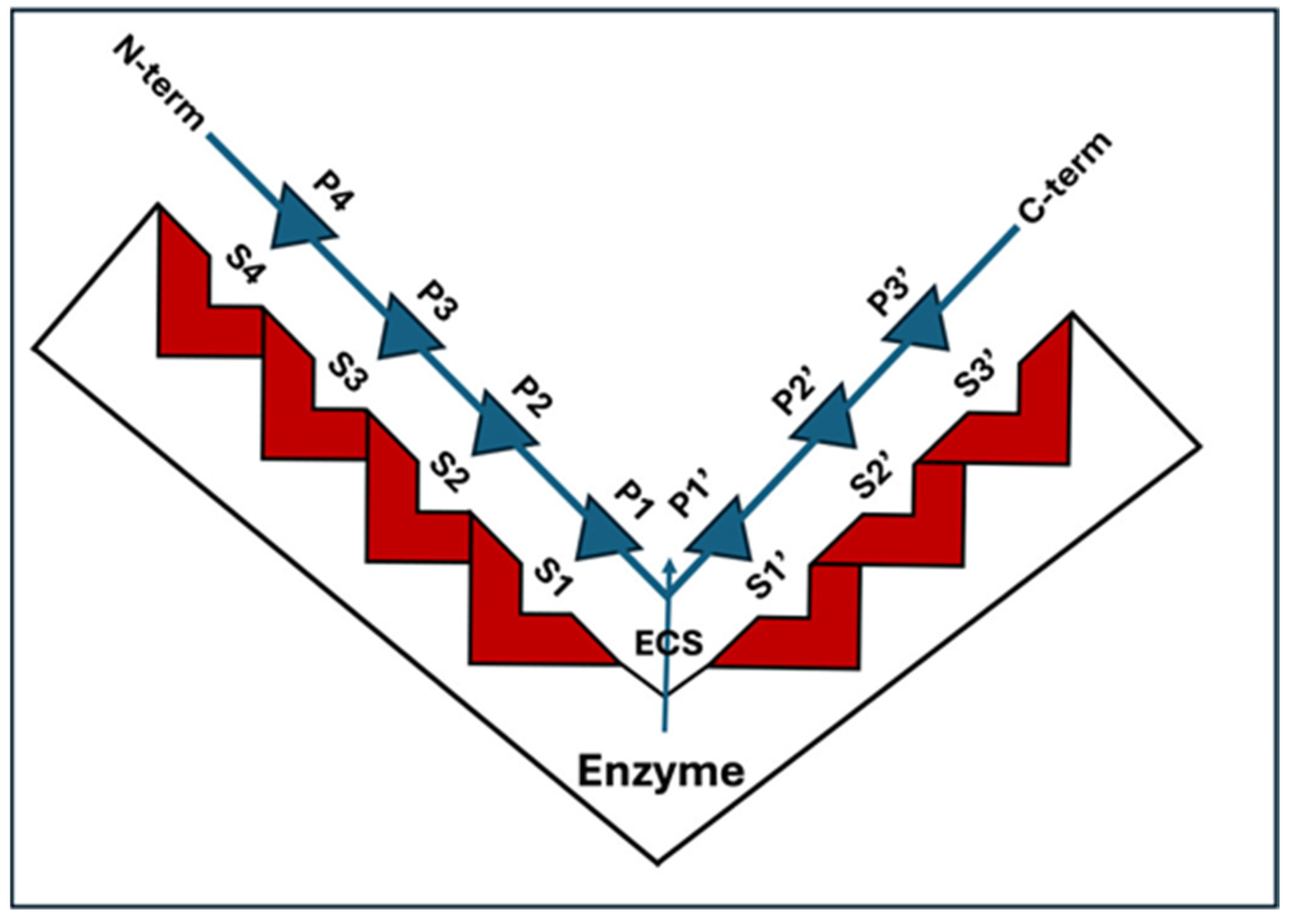
3. PLCP Substrates
4. Inhibition of Enzyme Activity
5. Resistance to Gastrointestinal Digestion
6. PLCP Allergenicity
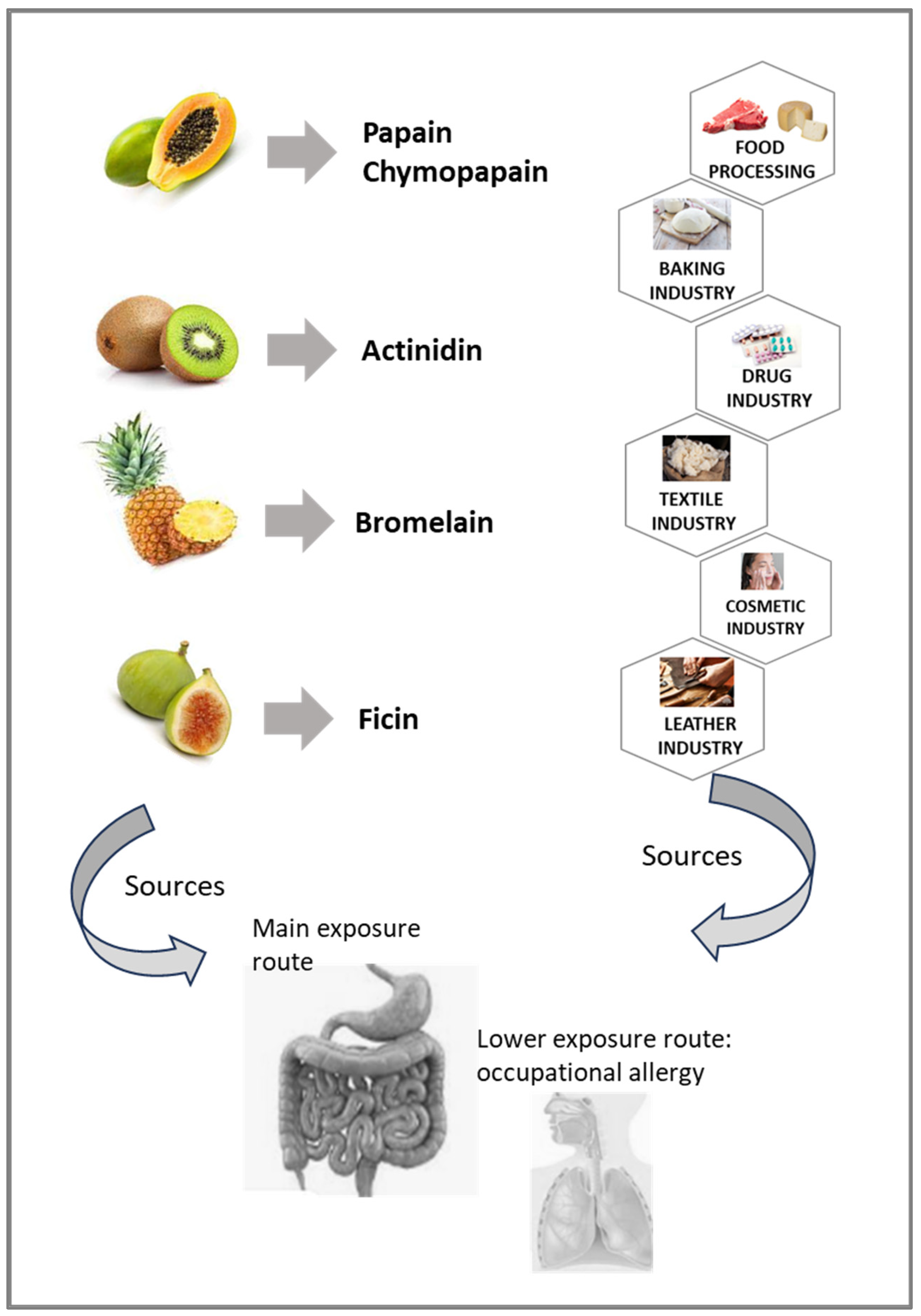
7. Sources of PLCPs Known for Allergenicity and/or IgE Binding
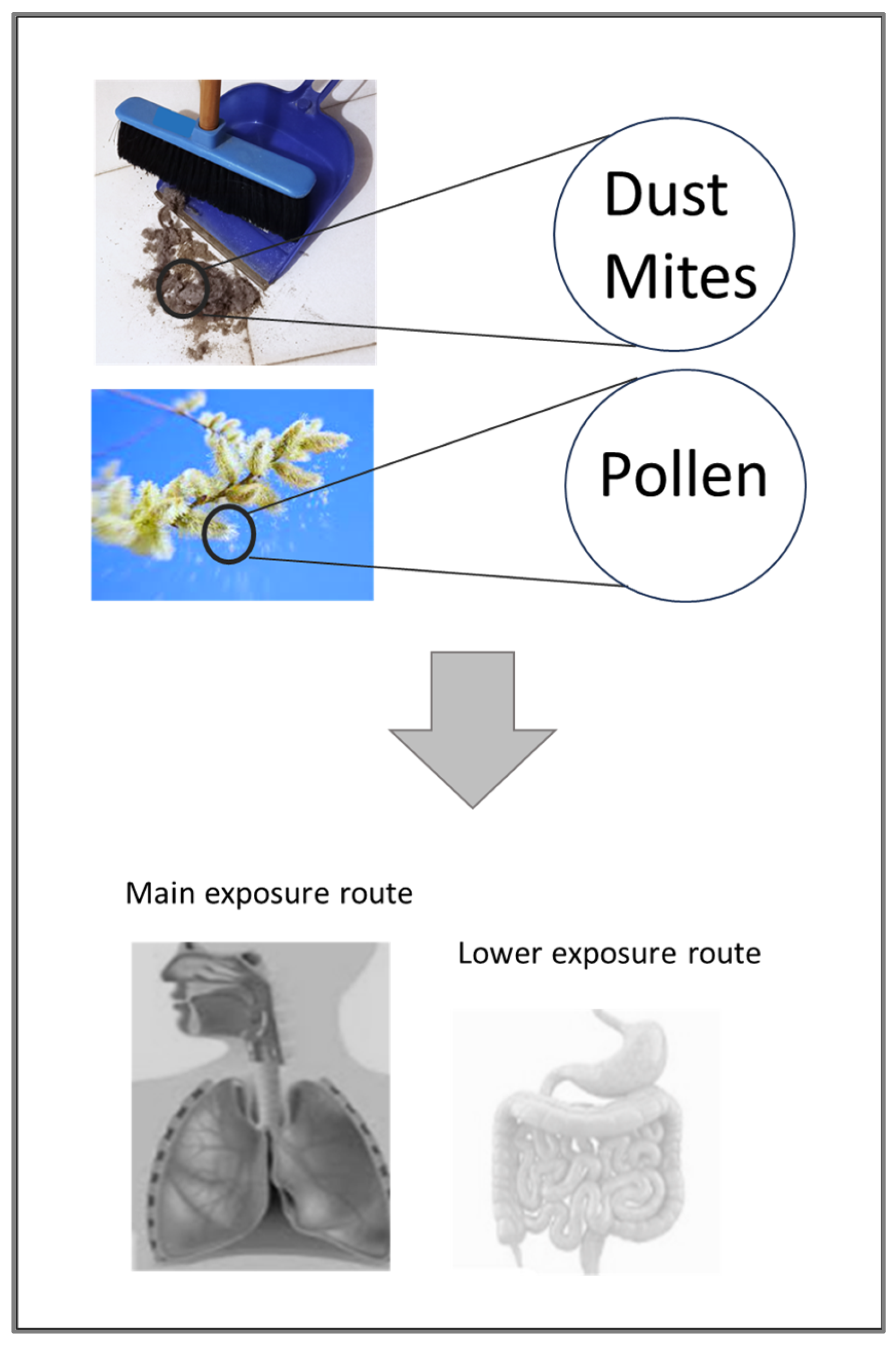
7.1. House Dust Mites (HDM)
7.2. Green Kiwifruit
7.3. Papaya
7.4. Pineapple
7.5. Fig
7.6. Short Ragweed
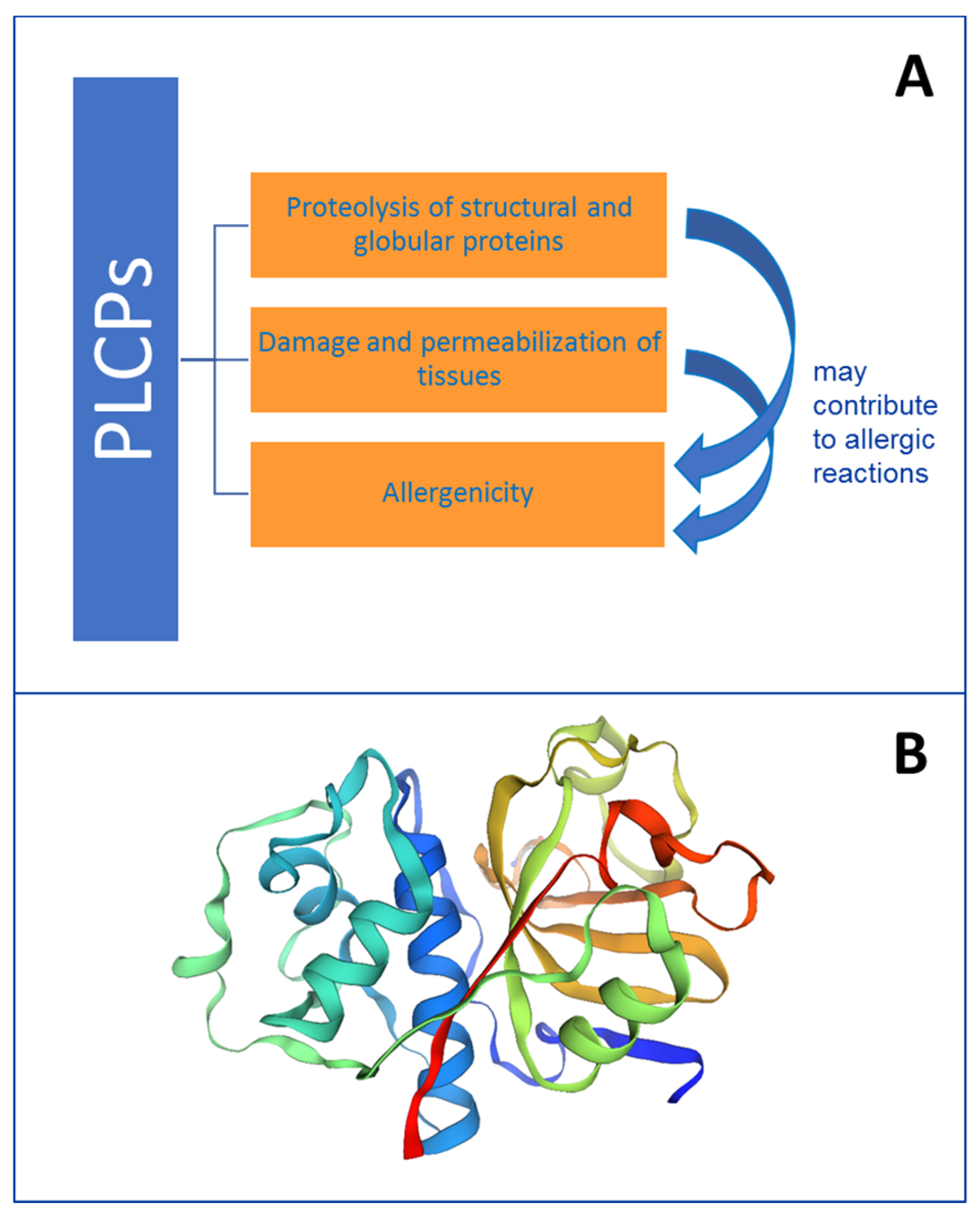
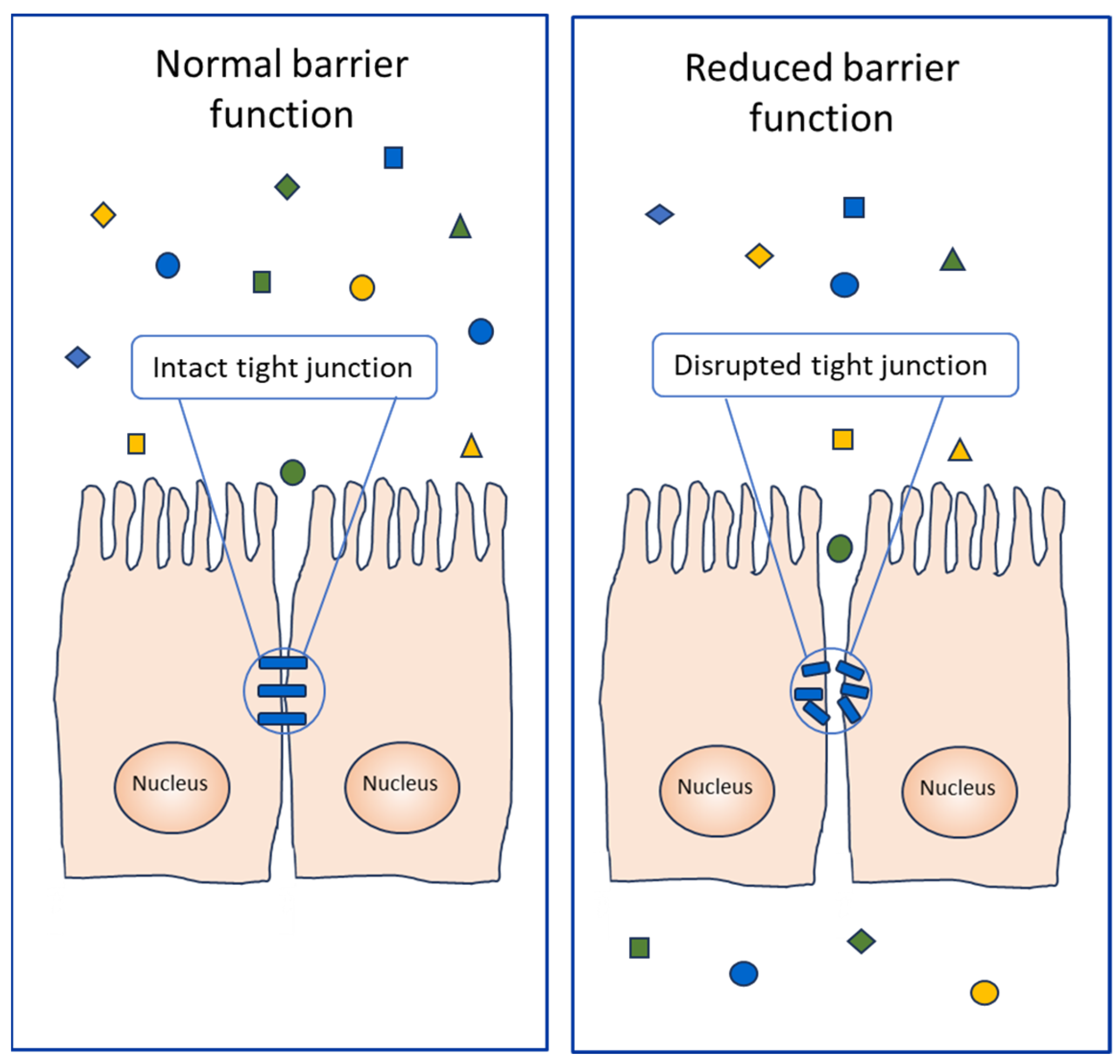
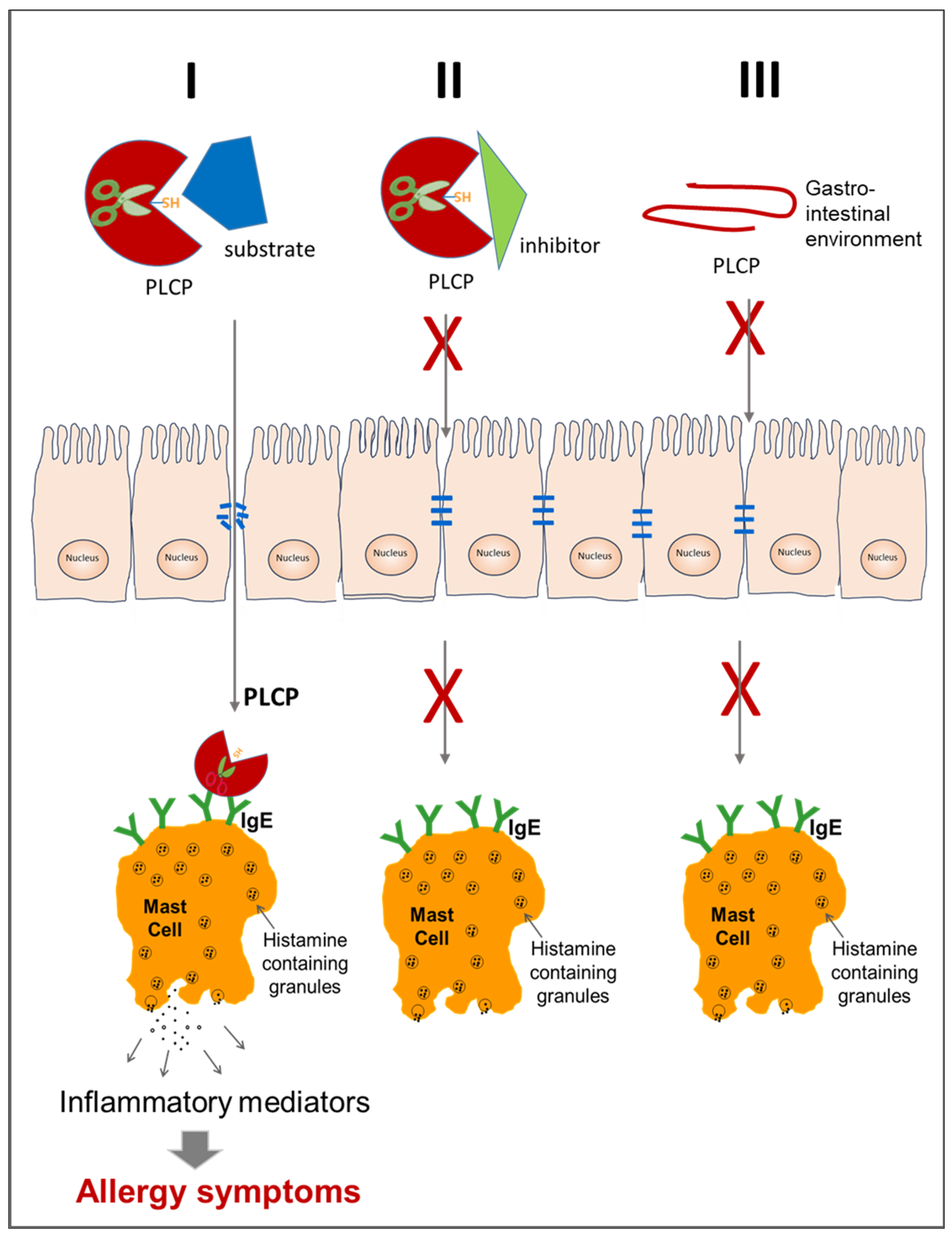
8. Contribution to Allergenicity of Enzyme Activity and Tissue Damage
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rawlings, N.D.; Bateman, A. How to use the MEROPS database and website to help understand peptidase specificity. Protein Sci. 2021, 30, 83–92. [Google Scholar] [CrossRef]
- Barrett, A.J.; Rawlings, N.D. Families and clans of cysteine peptidases. Perspect. Drug Discov. Des. 1996, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.D.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Res. 2018, 46, D624–D632. [Google Scholar] [CrossRef]
- Barrett, A.J.; Rawlings, N.D. Evolutionary lines of cysteine peptidases. Biol. Chem. 2001, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Zhao, W.; Yi, S.; Gao, W.; Zhang, J.; Zhao, J.; Liu, X. A new application of papain: As a peroxidase-like catalyst for fluorometric detection of uric acid. Enzyme Microb. Technol. 2023, 164, 110192. [Google Scholar] [CrossRef] [PubMed]
- Petushkova, A.I.; Savvateeva, L.V.; Zamyatnin, A.A. Structure determinants defining the specificity of papain-like cysteine proteases. Comput. Struct. Biotechnol. J. 2022, 20, 6552–6569. [Google Scholar] [CrossRef]
- Liu, H.; Hu, M.; Wang, Q.; Cheng, L.; Zhang, Z. Role of Papain-Like Cysteine Proteases in Plant Development. Front. Plant Sci. 2018, 9, 1717. [Google Scholar] [CrossRef]
- Verma, S.; Dixit, R.; Pandey, K.C. Cysteine Proteases: Modes of Activation and Future Prospects as Pharmacological Targets. Front. Pharmacol. 2016, 7, 107. [Google Scholar] [CrossRef]
- Giangrieco, I.; Ciardiello, M.A.; Tamburrini, M.; Tuppo, L.; Rafaiani, C.; Mari, A.; Alessandri, C. Comparative Analysis of the Immune Response and the Clinical Allergic Reaction to Papain-like Cysteine Proteases from Fig, Kiwifruit, Papaya, Pineapple and Mites in an Italian Population. Foods 2023, 12, 2852. [Google Scholar] [CrossRef]
- Bairoch, A. The ENZYME database in 2000. Nucleic Acids Res. 2000, 28, 304–305. [Google Scholar] [CrossRef]
- Pastorello, E.A.; Conti, A.; Pravettoni, V.; Farioli, L.; Rivolta, F.; Ansaloni, R.; Ispano, M.; Incorvaia, C.; Giuffrida, M.G.; Ortolani, C. Identification of actinidin as the major allergen of kiwi fruit. J. Allergy Clin. Immunol. 1998, 101, 531–537. [Google Scholar] [CrossRef]
- Grozdanovic, M.M.; Ostojic, S.; Aleksic, I.; Andjelkovic, U.; Petersen, A.; Gavrovic-Jankulovic, M. Active actinidin retains function upon gastro-intestinal digestion and is more thermostable than the E-64-inhibited counterpart. J. Sci. Food Agric. 2014, 94, 3046–3052. [Google Scholar] [CrossRef]
- Bublin, M.; Radauer, C.; Knulst, A.; Wagner, S.; Scheiner, O.; Mackie, A.R.; Mills, E.N.; Breiteneder, H. Effects of gastrointestinal digestion and heating on the allergenicity of the kiwi allergens Act d 1, actinidin, and Act d 2, a thaumatin-like protein. Mol. Nutr. Food Res. 2008, 52, 1130–1139. [Google Scholar] [CrossRef] [PubMed]
- Cavic, M.; Grozdanovic, M.M.; Bajic, A.; Jankovic, R.; Andjus, P.R.; Gavrovic-Jankulovic, M. The effect of kiwifruit (Actinidia deliciosa) cysteine protease actinidin on the occludin tight junction network in T84 intestinal epithelial cells. Food Chem. Toxicol. 2014, 72, 61–68. [Google Scholar] [CrossRef]
- Mostafaie, A.; Bidmeshkipour, A.; Shirvani, Z.; Mansouri, K.; Chalabi, M. Kiwifruit actinidin: A proper new collagenase for isolation of cells from different tissues. Appl. Biochem. Biotechnol. 2008, 144, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Chalabi, M.; Khademi, F.; Yarani, R.; Mostafaie, A. Proteolytic activities of kiwifruit actinidin (Actinidia deliciosa cv. Hayward) on different fibrous and globular proteins: A comparative study of actinidin with papain. Appl. Biochem. Biotechnol. 2014, 172, 4025–4037. [Google Scholar] [CrossRef] [PubMed]
- Mohd Azmi, S.I.; Kumar, P.; Sharma, N.; Sazili, A.Q.; Lee, S.J.; Ismail-Fitry, M.R. Application of Plant Proteases in Meat Tenderization: Recent Trends and Future Prospects. Foods 2023, 12, 1336. [Google Scholar] [CrossRef]
- Gailhofer, G.; Wilders-Truschnig, M.; Smolle, J.; Ludvan, M. Asthma caused by bromelain: An occupational allergy. Clin. Exp. Allergy 1988, 18, 445–450. [Google Scholar] [CrossRef]
- Ricardo, P.C.; Serudo, R.L.; Ţălu, Ş.; Lamarão, C.V.; da Fonseca Filho, H.D.; de Araújo Bezerra, J.; Sanches, E.A.; Campelo, P.H. Encapsulation of Bromelain in Combined Sodium Alginate and Amino Acid Carriers: Experimental Design of Simplex-Centroid Mixtures for Digestibility Evaluation. Molecules 2022, 27, 6364. [Google Scholar] [CrossRef]
- Kumar, V.; Mangla, B.; Javed, S.; Ahsan, W.; Kumar, P.; Garg, V.; Dureja, H. Bromelain: A review of its mechanisms, pharmacological effects and potential applications. Food Funct. 2023, 14, 8101–8128. [Google Scholar] [CrossRef]
- Ataide, J.A.; Cefali, L.C.; Figueiredo, M.C.; Braga, L.E.O.; Ruiz, A.L.T.G.; Foglio, M.A.; Oliveira-Nascimento, L.; Mazzola, P.G. In Vitro performance of free and encapsulated bromelain. Sci. Rep. 2021, 11, 10195. [Google Scholar] [CrossRef]
- Varilla, C.; Marcone, M.; Paiva, L.; Baptista, J. Bromelain, a Group of Pineapple Proteolytic Complex Enzymes. Foods 2021, 10, 2249. [Google Scholar] [CrossRef]
- Bhowmik, M.; Biswas Sarkar, M.; Kanti Sarkar, R.; Dasgupta, A.; Saha, S.; Jana, K.; Sircar, G.; Gupta Bhattacharya, S. Cloning and immunobiochemical analyses on recombinant chymopapain allergen Cari p 2 showing pollen-fruit cross-reaction. Mol. Immunol. 2021, 137, 42–51. [Google Scholar] [CrossRef]
- Nordby, E.J.; Wright, P.H.; Schofield, S.R. Safety of chemonucleolysis. Adverse effects reported in the United States, 1982–1991. Clin. Orthop. Relat. Res. 1993, 293, 122–134. [Google Scholar] [CrossRef]
- Bouley, J.; Groeme, R.; Le Mignon, M.; Jain, K.; Chabre, H.; Bordas-Le Floch, V.; Couret, M.N.; Bussières, L.; Lautrette, A.; Naveau, M.; et al. Identification of the cysteine protease Amb a 11 as a novel major allergen from short ragweed. J. Allergy Clin. Immunol. 2015, 136, 1055–1064. [Google Scholar] [CrossRef]
- Mora, C.; Flores, I.; Montealegre, F.; Díaz, A. Cloning and expression of Blo t 1, a novel allergen from the dust mite Blomia tropicalis, homologous to cysteine proteases. Clin. Exp. Allergy 2003, 33, 28–34. [Google Scholar] [CrossRef]
- Heymann, P.W.; Chapman, M.D.; Platts-Mills, T.A. Antigen Der f I from the dust mite Dermatophagoides farinae: Structural comparison with Der p I from Dermatophagoides pteronyssinus and epitope specificity of murine IgG and human IgE antibodies. J. Immunol. 1986, 137, 2841–2847. [Google Scholar] [CrossRef]
- Sanchez-Borges, M.; Capriles-Hulett, A.; Fernandez-Caldas, E.; Suarez-Chacon, R.; Caballero, F.; Castillo, S.; Sotillo, E. Mite-contaminated foods as a cause of anaphylaxis. J. Allergy Clin. Immunol. 1997, 99, 738–743. [Google Scholar] [CrossRef]
- Warner, A.; Boström, S.; Munir, A.K.; Möller, C.; Schou, C.; Kjellman, N.I. Environmental assessment of Dermatophagoides mite-allergen levels in Sweden should include Der m 1. Allergy 1998, 53, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Chapman, M.D.; Platts-Mills, T.A. Purification and characterization of the major allergen from Dermatophagoides pteronyssinus-antigen P1. J. Immunol. 1980, 125, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.; Mills, K.; Hazell, L.; Hart, B.; Thomas, W. Molecular analysis of the group 1 and 2 allergens from the house dust mite, Euroglyphus maynei. Int. Arch. Allergy Immunol. 1999, 118, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.H.; Tsai, J.J.; Lin, Y.H.; Yu, S.J.; Liao, E.C. Identification the Cross-Reactive or Species-Specific Allergens of Tyrophagus putrescentiae and Development Molecular Diagnostic Kits for Allergic Diseases. Diagnostics 2020, 10, 665. [Google Scholar] [CrossRef]
- Milošević, J.; Janković, B.; Prodanović, R.; Polović, N. Comparative stability of ficin and papain in acidic conditions and the presence of ethanol. Amino Acids 2019, 51, 829–838. [Google Scholar] [CrossRef]
- Aider, M. Potential applications of ficin in the production of traditional cheeses and protein hydrolysates. JDS Commun. 2021, 2, 233–237. [Google Scholar] [CrossRef]
- Tymoszuk, D.; Wiszniewska, M.; Walusiak-Skorupa, J. Papain-induced occupational rhinoconjunctivitis and asthma—A case report. Med. Pr. 2016, 67, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Novey, H.S.; Keenan, W.J.; Fairshter, R.D.; Wells, I.D.; Wilson, A.F.; Culver, B.D. Pulmonary disease in workers exposed to papain: Clinico-physiological and immunological studies. Clin. Allergy 1980, 10, 721–731. [Google Scholar] [CrossRef]
- Sharma, M.; Sharma, V.; Panda, A.K.; Majumdar, D.K. Development of enteric submicron particle formulation of papain for oral delivery. Int. J. Nanomed. 2011, 6, 2097–2111. [Google Scholar] [CrossRef]
- Trevisol, T.C.; Henriques, R.O.; Souza, A.J.A.; Furigo, A. An overview of the use of proteolytic enzymes as exfoliating agents. J. Cosmet. Dermatol. 2022, 21, 3300–3307. [Google Scholar] [CrossRef]
- Chapman, M.D.; Wünschmann, S.; Pomés, A. Proteases as Th2 adjuvants. Curr. Allergy Asthma Rep. 2007, 7, 363–367. [Google Scholar] [CrossRef]
- Topham, C.M.; Srinivasan, N.; Thorpe, C.J.; Overington, J.P.; Kalsheker, N.A. Comparative modelling of major house dust mite allergen Der p I: Structure validation using an extended environmental amino acid propensity table. Protein Eng. 1994, 7, 869–894. [Google Scholar] [CrossRef]
- Denessiouk, K.; Uversky, V.N.; Permyakov, S.E.; Permyakov, E.A.; Johnson, M.S.; Denesyuk, A.I. Papain-like cysteine proteinase zone (PCP-zone) and PCP structural catalytic core (PCP-SCC) of enzymes with cysteine proteinase fold. Int. J. Biol. Macromol. 2020, 165, 1438–1446. [Google Scholar] [CrossRef]
- Chruszcz, M.; Chapman, M.D.; Vailes, L.D.; Stura, E.A.; Saint-Remy, J.M.; Minor, W.; Pomés, A. Crystal structures of mite allergens Der f 1 and Der p 1 reveal differences in surface-exposed residues that may influence antibody binding. J. Mol. Biol. 2009, 386, 520–530. [Google Scholar] [CrossRef]
- Matsumoto, K.; Mizoue, K.; Kitamura, K.; Tse, W.C.; Huber, C.P.; Ishida, T. Structural basis of inhibition of cysteine proteases by E-64 and its derivatives. Biopolymers 1999, 51, 99–107. [Google Scholar] [CrossRef]
- Chan, S.K.; Pomés, A.; Hilger, C.; Davies, J.M.; Mueller, G.; Kuehn, A.; Lopata, A.L.; Gadermaier, G.; van Hage, M.; Raulf, M.; et al. Keeping Allergen Names Clear and Defined. Front. Immunol. 2019, 10, 2600. [Google Scholar] [CrossRef]
- SMITH, E.L. Active site of papain and covalent high-energy bonds of proteins. J. Biol. Chem. 1958, 233, 1392–1397. [Google Scholar] [CrossRef]
- Liu, T.Y.; Stein, W.H.; Moore, S.; Elliott, S.D. The sequence of amino acid residues around the sulfhydryl group at the active site of streptococcal proteinase. J. Biol. Chem. 1965, 240, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- van Duijnen, P.T.; Thole, B.T.; Hol, W.G. On the role of the active site helix in papain, an ab initio molecular orbital study. Biophys. Chem. 1979, 9, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Schechter, I.; Berger, A. On the size of the active site in proteases. I. Papain. Biochem. Biophys. Res. Commun. 1967, 27, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Turk, D.; Turk, B.; Turk, V. Papain-like lysosomal cysteine proteases and their inhibitors: Drug discovery targets? Biochem. Soc. Symp. 2003, 70, 15–30. [Google Scholar]
- Rawlings, N.D.; Barrett, A.J. Families of cysteine peptidases. Methods Enzymol. 1994, 244, 461–486. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.L.; Backes, B.J.; Leonetti, F.; Mahrus, S.; Ellman, J.A.; Craik, C.S. Rapid and general profiling of protease specificity by using combinatorial fluorogenic substrate libraries. Proc. Natl. Acad. Sci. USA 2000, 97, 7754–7759. [Google Scholar] [CrossRef]
- Lapek, J.D.; Jiang, Z.; Wozniak, J.M.; Arutyunova, E.; Wang, S.C.; Lemieux, M.J.; Gonzalez, D.J.; O’Donoghue, A.J. Quantitative Multiplex Substrate Profiling of Peptidases by Mass Spectrometry. Mol. Cell. Proteom. 2019, 18, 968–981. [Google Scholar] [CrossRef]
- Otto, H.H.; Schirmeister, T. Cysteine Proteases and Their Inhibitors. Chem. Rev. 1997, 97, 133–172. [Google Scholar] [CrossRef] [PubMed]
- Varughese, K.I.; Su, Y.; Cromwell, D.; Hasnain, S.; Xuong, N.H. Crystal structure of an actinidin-E-64 complex. Biochemistry 1992, 31, 5172–5176. [Google Scholar] [CrossRef] [PubMed]
- Hanada, K.; Tamai, M.; Yamagishi, M.; Ohmura, S.; Sawada, J.; Tanaka, I. Isolation and Characterization of E–64, a New Thiol Protease Inhibitor. Agric. Biol. Chem. 1978, 42, 523–528. [Google Scholar] [CrossRef]
- Grozdanović, M.M.; Drakulić, B.J.; Gavrović-Jankulović, M. Conformational mobility of active and E-64-inhibited actinidin. Biochim. Biophys. Acta 2013, 1830, 4790–4799. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Wang, Q.; Su, H.; Shao, Q.; Zhao, W.; Chen, G.; Li, M.; Xu, Y. Identification of Cysteine 270 as a Novel Site for Allosteric Modulators of SARS-CoV-2 Papain-Like Protease. Angew. Chem. Int. Ed. Engl. 2022, 61, e202212378. [Google Scholar] [CrossRef] [PubMed]
- Hernández Alvarez, L.; Barreto Gomes, D.E.; Hernández González, J.E.; Pascutti, P.G. Dissecting a novel allosteric mechanism of cruzain: A computer-aided approach. PLoS ONE 2019, 14, e0211227. [Google Scholar] [CrossRef] [PubMed]
- Stremnitzer, C.; Manzano-Szalai, K.; Willensdorfer, A.; Starkl, P.; Pieper, M.; König, P.; Mildner, M.; Tschachler, E.; Reichart, U.; Jensen-Jarolim, E. Papain Degrades Tight Junction Proteins of Human Keratinocytes In Vitro and Sensitizes C57BL/6 Mice via the Skin Independent of its Enzymatic Activity or TLR4 Activation. J. Investig. Dermatol. 2015, 135, 1790–1800. [Google Scholar] [CrossRef]
- Díez-Gómez, M.L.; Quirce, S.; Aragoneses, E.; Cuevas, M. Asthma caused by Ficus benjamina latex: Evidence of cross-reactivity with fig fruit and papain. Ann. Allergy Asthma Immunol. 1998, 80, 24–30. [Google Scholar] [CrossRef]
- Gratacós Gómez, A.R.; Gómez Torrijos, E. Eosinophilic Esophagitis due to Aeroallergens: A Systematic Review and Update. J. Investig. Allergol. Clin. Immunol. 2022, 32, 438–450. [Google Scholar] [CrossRef] [PubMed]
- Visaggi, P.; Savarino, E.; Del Corso, G.; Hunter, H.; Baiano Svizzero, F.; Till, S.J.; Dunn, J.; Wong, T.; de Bortoli, N.; Zeki, S. Six-Food Elimination Diet Is Less Effective during Pollen Season in Adults with Eosinophilic Esophagitis Sensitized to Pollens. Am. J. Gastroenterol. 2023, 118, 1957–1962. [Google Scholar] [CrossRef] [PubMed]
- Koken, G.; Ertoy Karagol, H.I.; Polat Terece, S.; Cavdar, Z.; Cetin, K.; Egritas Gurkan, O.; Sari, S.; Dalgic, B.; Bakirtas, A. Food-induced immediate response of the esophagus in pediatric eosinophilic esophagitis. Allergy 2023, 78, 3235–3240. [Google Scholar] [CrossRef] [PubMed]
- Fuiano, N.; Incorvaia, C.; Riario-Sforza, G.G.; Casino, G. Anaphylaxis to honey in pollinosis to mugwort: A case report. Eur. Ann. Allergy Clin. Immunol. 2006, 38, 364–365. [Google Scholar] [PubMed]
- Li, J.D.; Cui, L.; Xu, Y.Y.; Guan, K. A Case of Anaphylaxis Caused by Major Royal Jelly Protein 3 of Royal Jelly and Its Cross-Reactivity with Honeycomb. J. Asthma Allergy 2021, 14, 1555–1557. [Google Scholar] [CrossRef]
- Tulic, M.K.; Vivinus-Nébot, M.; Rekima, A.; Rabelo Medeiros, S.; Bonnart, C.; Shi, H.; Walker, A.; Dainese, R.; Boyer, J.; Vergnolle, N.; et al. Presence of commensal house dust mite allergen in human gastrointestinal tract: A potential contributor to intestinal barrier dysfunction. Gut 2016, 65, 757–766. [Google Scholar] [CrossRef]
- Soh, W.T.; Zhang, J.; Hollenberg, M.D.; Vliagoftis, H.; Rothenberg, M.E.; Sokol, C.L.; Robinson, C.; Jacquet, A. Protease allergens as initiators-regulators of allergic inflammation. Allergy 2023, 78, 1148–1168. [Google Scholar] [CrossRef]
- Al-Ghouleh, A.; Johal, R.; Sharquie, I.K.; Emara, M.; Harrington, H.; Shakib, F.; Ghaemmaghami, A.M. The glycosylation pattern of common allergens: The recognition and uptake of Der p 1 by epithelial and dendritic cells is carbohydrate dependent. PLoS ONE 2012, 7, e33929. [Google Scholar] [CrossRef]
- van der Heide, S.; De Monchy, J.G.; De Vries, K.; Dubois, A.E.; Kauffman, H.F. Seasonal differences in airway hyperresponsiveness in asthmatic patients: Relationship with allergen exposure and sensitization to house dust mites. Clin. Exp. Allergy 1997, 27, 627–633. [Google Scholar] [CrossRef]
- Baur, X.; Fruhmann, G. Allergic reactions, including asthma, to the pineapple protease bromelain following occupational exposure. Clin. Allergy 1979, 9, 443–450. [Google Scholar] [CrossRef]
- Sagona, M.A.; Bruszer, G.V.; Nelson, J.C.; Mascoli, C.; Serkes, K. Reactivity of IgE and IgG serum levels to chymopapain after chemonucleolysis. J. Allergy Clin. Immunol. 1985, 75, 659–662. [Google Scholar] [CrossRef] [PubMed]
- Nettis, E.; Napoli, G.; Ferrannini, A.; Tursi, A. IgE-mediated allergy to bromelain. Allergy 2001, 56, 257–258. [Google Scholar] [CrossRef] [PubMed]
- Tuppo, L.; Giangrieco, I.; Tamburrini, M.; Alessandri, C.; Mari, A.; Ciardiello, M.A. Detection of Allergenic Proteins in Foodstuffs: Advantages of the Innovative Multiplex Allergen Microarray-Based Immunoassay Compared to Conventional Methods. Foods 2022, 11, 878. [Google Scholar] [CrossRef] [PubMed]
- Alessandri, C.; Giangrieco, I.; Tuppo, L.; Ferrara, R.; Zennaro, D.; Bernardi, M.L.; Ciancamerla, M.; Rafaiani, C.; Tamburrini, M.; Mari, A.; et al. Are peas a safe food for lipid transfer protein allergic patients? Allergy 2021, 76, 2587–2589. [Google Scholar] [CrossRef]
- Urbani, S.; Aruanno, A.; Nucera, E. Adverse reaction to. Clin. Mol. Allergy 2020, 18, 9. [Google Scholar] [CrossRef] [PubMed]
- Widmer, F.; Hayes, P.J.; Whittaker, R.G.; Kumar, R.K. Substrate preference profiles of proteases released by allergenic pollens. Clin. Exp. Allergy 2000, 30, 571–576. [Google Scholar] [CrossRef]
- Gunawan, H.; Takai, T.; Ikeda, S.; Okumura, K.; Ogawa, H. Protease activity of allergenic pollen of cedar, cypress, juniper, birch and ragweed. Allergol. Int. 2008, 57, 83–91. [Google Scholar] [CrossRef]
- Wiederstein, M.; Baumgartner, S.; Lauter, K. Soybean (Glycinemax) allergens—A Review on an Outstanding Plant Food with Allergenic Potential. ACS Food Sci. Technol. 2023, 3, 363–378. [Google Scholar] [CrossRef]
- Sharma, A.; Vashisht, S.; Mishra, R.; Gaur, S.N.; Prasad, N.; Lavasa, S.; Batra, J.K.; Arora, N. Molecular and immunological characterization of cysteine protease from Phaseolus vulgaris and evolutionary cross-reactivity. J. Food Biochem. 2022, 46, e14232. [Google Scholar] [CrossRef]
- Lee, A.J.; Machell, J.; Van Den Broek, A.H.; Nisbet, A.J.; Miller, H.R.; Isaac, R.E.; Huntley, J.F. Identification of an antigen from the sheep scab mite, Psoroptes ovis, homologous with house dust mite group I allergens. Parasite Immunol. 2002, 24, 413–422. [Google Scholar] [CrossRef]
- Acevedo, N.; Zakzuk, J.; Caraballo, L. House Dust Mite Allergy Under Changing Environments. Allergy Asthma Immunol. Res. 2019, 11, 450–469. [Google Scholar] [CrossRef] [PubMed]
- Thomas, W.R.; Smith, W. House-dust-mite allergens. Allergy 1998, 53, 821–832. [Google Scholar] [CrossRef]
- Takai, T.; Kato, T.; Hatanaka, H.; Inui, K.; Nakazawa, T.; Ichikawa, S.; Mitsuishi, K.; Ogawa, H.; Okumura, K. Modulation of allergenicity of major house dust mite allergens Der f 1 and Der p 1 by interaction with an endogenous ligand. J. Immunol. 2009, 183, 7958–7965. [Google Scholar] [CrossRef]
- Chevigné, A.; Barumandzadeh, R.; Groslambert, S.; Cloes, B.; Dehareng, D.; Filée, P.; Marx, J.C.; Frère, J.M.; Matagne, A.; Jacquet, A.; et al. Relationship between propeptide pH unfolding and inhibitory ability during ProDer p 1 activation mechanism. J. Mol. Biol. 2007, 374, 170–185. [Google Scholar] [CrossRef] [PubMed]
- Rayapudi, M.; Mavi, P.; Zhu, X.; Pandey, A.K.; Abonia, J.P.; Rothenberg, M.E.; Mishra, A. Indoor insect allergens are potent inducers of experimental eosinophilic esophagitis in mice. J. Leukoc. Biol. 2010, 88, 337–346. [Google Scholar] [CrossRef]
- Simon, D.; Straumann, A.; Dahinden, C.; Simon, H.U. Frequent sensitization to Candida albicans and profilins in adult eosinophilic esophagitis. Allergy 2013, 68, 945–948. [Google Scholar] [CrossRef]
- Béné, J.; Ley, D.; Roboubi, R.; Gottrand, F.; Gautier, S. Eosinophilic esophagitis after desensitization to dust mites with sublingual immunotherapy. Ann. Allergy Asthma Immunol. 2016, 116, 583–584. [Google Scholar] [CrossRef]
- Liccardi, G.; D’Amato, M.; D’Amato, G. Oral allergy syndrome after ingestion of salami in a subject with monosensitization to mite allergens. J. Allergy Clin. Immunol. 1996, 98, 850–852. [Google Scholar] [CrossRef]
- Sánchez-Borges, M.; Fernandez-Caldas, E. Hidden allergens and oral mite anaphylaxis: The pancake syndrome revisited. Curr. Opin Allergy Clin. Immunol. 2015, 15, 337–343. [Google Scholar] [CrossRef]
- Lombardero, M.; Heymann, P.W.; Platts-Mills, T.A.; Fox, J.W.; Chapman, M.D. Conformational stability of B cell epitopes on group I and group II Dermatophagoides spp. allergens. Effect of thermal and chemical denaturation on the binding of murine IgG and human IgE antibodies. J. Immunol. 1990, 144, 1353–1360. [Google Scholar] [CrossRef]
- Chevigné, A.; Jacquet, A. Emerging roles of the protease allergen Der p 1 in house dust mite-induced airway inflammation. J. Allergy Clin. Immunol. 2018, 142, 398–400. [Google Scholar] [CrossRef]
- Jacquet, A. The HDM allergen orchestra and its cysteine protease maestro: Stimulators of kaleidoscopic innate immune responses. Mol. Immunol. 2023, 156, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Hirasawa, Y.; Takai, T.; Mitsuishi, K.; Okuda, M.; Kato, T.; Okumura, K.; Ikeda, S.; Ogawa, H. Reduction of skin barrier function by proteolytic activity of a recombinant house dust mite major allergen Der f 1. J. Investig. Dermatol. 2006, 126, 2719–2723. [Google Scholar] [CrossRef]
- Ferreira, F.; Hawranek, T.; Gruber, P.; Wopfner, N.; Mari, A. Allergic cross-reactivity: From gene to the clinic. Allergy 2004, 59, 243–267. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.; Iraola, V.; Leonor, J.R.; Bartra, J.; Rodríguez, F.; Boquete, M.; Huertas, Á.; Paniagua, M.J.; Pinto, H.; Carnés, J. Different sensitization to storage mites depending on the co-exposure to house dust mites. Ann. Allergy Asthma Immunol. 2015, 114, 36–42.e31. [Google Scholar] [CrossRef]
- Sánchez-Borges, M.; Suárez-Chacón, R.; Capriles-Hulett, A.; Caballero-Fonseca, F. An update on oral anaphylaxis from mite ingestion. Ann. Allergy Asthma Immunol. 2005, 94, 216–220; quiz 220–212, 306. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Machín, I.; Glez-Paloma Poza, R.; Iglesias-Souto, J.; Iraola, V.; Matheu, V. Oral mite anaphylaxis. Allergy 2010, 65, 1345–1347. [Google Scholar] [CrossRef] [PubMed]
- Bell, C.H. 15—Pest control of stored food products: Insects and mites. In Hygiene in Food Processing; Lelieveld, H.L.M., Holah, J.T., Napper, D., Eds.; Woodhead Publishing: Cambridge, UK, 2014. [Google Scholar] [CrossRef]
- Brückner, A.; Heethoff, M. Scent of a mite: Origin and chemical characterization of the lemon-like flavor of mite-ripened cheeses. Exp. Appl. Acarol. 2016, 69, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Bublin, M.; Mari, A.; Ebner, C.; Knulst, A.; Scheiner, O.; Hoffmann-Sommergruber, K.; Breiteneder, H.; Radauer, C. IgE sensitization profiles toward green and gold kiwifruits differ among patients allergic to kiwifruit from 3 European countries. J. Allergy Clin. Immunol. 2004, 114, 1169–1175. [Google Scholar] [CrossRef]
- Tuppo, L.; Giangrieco, I.; Palazzo, P.; Bernardi, M.L.; Scala, E.; Carratore, V.; Tamburrini, M.; Mari, A.; Ciardiello, M.A. Kiwellin, a modular protein from green and gold kiwi fruits: Evidence of in vivo and in vitro processing and IgE binding. J. Agric. Food Chem. 2008, 56, 3812–3817. [Google Scholar] [CrossRef]
- Nieuwenhuizen, N.J.; Beuning, L.L.; Sutherland, P.W.; Sharma, N.N.; Cooney, J.M.; Bieleski, L.R.F.; Schröder, R.; MacRae, E.A.; Atkinson, R.G. Identification and characterisation of acidic and novel basic forms of actinidin, the highly abundant cysteine protease from kiwifruit. Funct. Plant Biol. 2007, 34, 946–961. [Google Scholar] [CrossRef]
- Bublin, M.; Dennstedt, S.; Buchegger, M.; Antonietta Ciardiello, M.; Bernardi, M.L.; Tuppo, L.; Harwanegg, C.; Hafner, C.; Ebner, C.; Ballmer-Weber, B.K.; et al. The performance of a component-based allergen microarray for the diagnosis of kiwifruit allergy. Clin. Exp. Allergy 2011, 41, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Grozdanovic, M.; Popovic, M.; Polovic, N.; Burazer, L.; Vuckovic, O.; Atanaskovic-Markovic, M.; Lindner, B.; Petersen, A.; Gavrovic-Jankulovic, M. Evaluation of IgE reactivity of active and thermally inactivated actinidin, a biomarker of kiwifruit allergy. Food Chem. Toxicol. 2012, 50, 1013–1018. [Google Scholar] [CrossRef] [PubMed]
- Ciardiello, M.A.; Meleleo, D.; Saviano, G.; Crescenzo, R.; Carratore, V.; Camardella, L.; Gallucci, E.; Micelli, S.; Tancredi, T.; Picone, D.; et al. Kissper, a kiwi fruit peptide with channel-like activity: Structural and functional features. J. Pept. Sci. 2008, 14, 742–754. [Google Scholar] [CrossRef]
- Grozdanovic, M.M.; Čavić, M.; Nešić, A.; Andjelković, U.; Akbari, P.; Smit, J.J.; Gavrović-Jankulović, M. Kiwifruit cysteine protease actinidin compromises the intestinal barrier by disrupting tight junctions. Biochim. Biophys. Acta 2016, 1860, 516–526. [Google Scholar] [CrossRef] [PubMed]
- Bekhit, A.A.; Hopkins, D.L.; Geesink, G.; Franks, P. Exogenous proteases for meat tenderization. Crit. Rev. Food Sci. Nutr. 2014, 54, 1012–1031. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Bekhit, A.E.-D.A.; Carne, A.; Hopkins, D.L. Characterisation of commercial papain, bromelain, actinidin and zingibain protease preparations and their activities toward meat proteins. Food Chem. 2012, 134, 95–105. [Google Scholar] [CrossRef]
- Palacin, A.; Rodriguez, J.; Blanco, C.; Lopez-Torrejon, G.; Sánchez-Monge, R.; Varela, J.; Jiménez, M.A.; Cumplido, J.; Carrillo, T.; Crespo, J.F.; et al. Immunoglobulin E recognition patterns to purified Kiwifruit (Actinidinia deliciosa) allergens in patients sensitized to Kiwi with different clinical symptoms. Clin. Exp. Allergy 2008, 38, 1220–1228. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.S.; Nieuwenhuizen, N.J.; Atkinson, R.G.; Macrae, E.A.; Cochrane, S.A.; Warner, J.O.; Hourihane, J.O. Kiwifruit allergy: Actinidin is not a major allergen in the United Kingdom. Clin. Exp. Allergy 2007, 37, 1340–1348. [Google Scholar] [CrossRef]
- Ciardiello, M.A.; Giangrieco, I.; Tuppo, L.; Tamburrini, M.; Buccheri, M.; Palazzo, P.; Bernardi, M.L.; Ferrara, R.; Mari, A. Influence of the natural ripening stage, cold storage, and ethylene treatment on the protein and IgE-binding profiles of green and gold kiwi fruit extracts. J. Agric. Food Chem. 2009, 57, 1565–1571. [Google Scholar] [CrossRef]
- Gavrovic-Jankulovic, M.; Polovic, N.; Prisic, S.; Jankov, R.M.; Atanaskovic-Markovic, M.; Vuckovic, O.; Velickovic, T.C. Allergenic potency of kiwi fruit during fruit development. Food Agric. Immunol. 2005, 16, 117–128. [Google Scholar] [CrossRef]
- Gall, H.; Kalveram, K.J.; Forck, G.; Sterry, W. Kiwi fruit allergy: A new birch pollen-associated food allergy. J. Allergy Clin. Immunol. 1994, 94, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Yin, J.; Wen, L. Papain Induced Occupational Asthma with Kiwi and Fig Allergy. Allergy Asthma Immunol. Res. 2016, 8, 170–173. [Google Scholar] [CrossRef]
- Eagle, H.; Harris, T.N. Studies in blood coagulation: V. the coagulation of blood by proteolytic enzymes (trypsin, papain). J. Gen. Physiol. 1937, 20, 543–560. [Google Scholar] [CrossRef]
- Drenth, J.; Jansonius, J.N.; Koekoek, R.; Swen, H.M.; Wolthers, B.G. Structure of papain. Nature 1968, 218, 929–932. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sharma, A.; Niewiara, M.J.; Singh, R.; Ming, R.; Yu, Q. Papain-like cysteine proteases in Carica papaya: Lineage-specific gene duplication and expansion. BMC Genom. 2018, 19, 26. [Google Scholar] [CrossRef] [PubMed]
- Drugs and Lactation Database (LactMed®); National Institute of Child Health and Human Development: Bethesda, MD, USA, 2006. Available online: https://www.ncbi.nlm.nih.gov/books/NBK501922/ (accessed on 10 January 2024).
- Dando, P.M.; Sharp, S.L.; Buttle, D.J.; Barrett, A.J. Immunoglobulin E antibodies to papaya proteinases and their relevance to chemonucleolysis. Spine 1995, 20, 981–985. [Google Scholar] [CrossRef]
- Maes, D.; Bouckaert, J.; Poortmans, F.; Wyns, L.; Looze, Y. Structure of chymopapain at 1.7 A resolution. Biochemistry 1996, 35, 16292–16298. [Google Scholar] [CrossRef]
- Cogen, F.C.; Goldstein, M.; Zweiman, B. Skin testing in chymopapain anaphylaxis. J. Allergy Clin. Immunol. 1985, 75, 728–730. [Google Scholar] [CrossRef]
- Freddi, G.; Mossotti, R.; Innocenti, R. Degumming of silk fabric with several proteases. J. Biotechnol. 2003, 106, 101–112. [Google Scholar] [CrossRef]
- Seaf, M.; Ben-Zimra, M.; Mankuta, D.; Dayan, N.; Levi-Schaffer, F. Papain Activates Human Mast Cells to Release Proinflammatory Mediators via its Enzymatic Activity. J. Investig. Dermatol. 2016, 136, 1523–1525. [Google Scholar] [CrossRef]
- Perner, C.; Flayer, C.H.; Zhu, X.; Aderhold, P.A.; Dewan, Z.N.A.; Voisin, T.; Camire, R.B.; Chow, O.A.; Chiu, I.M.; Sokol, C.L. Substance P Release by Sensory Neurons Triggers Dendritic Cell Migration and Initiates the Type-2 Immune Response to Allergens. Immunity 2020, 53, 1063–1077.e1067. [Google Scholar] [CrossRef] [PubMed]
- Miike, S.; Kita, H. Human eosinophils are activated by cysteine proteases and release inflammatory mediators. J. Allergy Clin. Immunol. 2003, 111, 704–713. [Google Scholar] [CrossRef]
- Flindt, M.L. Respiratory hazards from papain. Lancet 1978, 1, 430–432. [Google Scholar] [CrossRef] [PubMed]
- Goeminne, P.C.; Adams, E.; Deschepper, K.; Valcke, Y.; Nemery, B. Papain-induced asthma: A man with dyspnea from dawn till dust. Acta Clin. Belg. 2013, 68, 132–134. [Google Scholar] [CrossRef] [PubMed]
- van Kampen, V.; Merget, R.; Brüning, T. Occupational allergies to papain. Pneumologie 2005, 59, 405–410. [Google Scholar] [CrossRef]
- Mansfield, L.E.; Bowers, C.H. Systemic reaction to papain in a nonoccupational setting. J. Allergy Clin. Immunol. 1983, 71, 371–374. [Google Scholar] [CrossRef]
- Arshad, Z.I.; Amid, A.; Yusof, F.; Jaswir, I.; Ahmad, K.; Loke, S.P. Bromelain: An overview of industrial application and purification strategies. Appl. Microbiol. Biotechnol. 2014, 98, 7283–7297. [Google Scholar] [CrossRef]
- Galleguillos, F.; Rodriguez, J.C. Asthma caused by bromelin inhalation. Clin. Allergy 1978, 8, 21–24. [Google Scholar] [CrossRef]
- Baur, X. Studies on the specificity of human IgE-antibodies to the plant proteases papain and bromelain. Clin. Allergy 1979, 9, 451–457. [Google Scholar] [CrossRef]
- Platts-Mills, T.A.; Hilger, C.; Jappe, U.; van Hage, M.; Gadermaier, G.; Spillner, E.; Lidholm, J.; Keshavarz, B.; Aalberse, R.C.; van Ree, R.; et al. Carbohydrate epitopes currently recognized as targets for IgE antibodies. Allergy 2021, 76, 2383–2394. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, S.; Arai, S.; Watanabe, M. Modification of wheat flour with bromelain and baking hypoallergenic bread with added ingredients. Biosci. Biotechnol. Biochem. 1996, 60, 1269–1272. [Google Scholar] [CrossRef] [PubMed]
- van Kampen, V.; Merget, R.; Brüning, T. Occupational allergies to bromelain. Pneumologie 2007, 61, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Haesaerts, S.; Rodriguez Buitrago, J.A.; Loris, R.; Baeyens-Volant, D.; Azarkan, M. Crystallization and preliminary X-ray analysis of four cysteine proteases from Ficus carica latex. Acta Crystallogr. F Struct. Biol. Commun. 2015, 71, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Raskovic, B.; Lazic, J.; Polovic, N. Characterisation of general proteolytic, milk clotting and antifungal activity of Ficus carica latex during fruit ripening. J. Sci. Food Agric. 2016, 96, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Dechamp, C.; Bessot, J.C.; Pauli, G.; Deviller, P. First report of anaphylactic reaction after fig (Ficus carica) ingestion. Allergy 1995, 50, 514–516. [Google Scholar] [CrossRef]
- Hemmer, W.; Focke, M.; Götz, M.; Jarisch, R. Sensitization to Ficus benjamina: Relationship to natural rubber latex allergy and identification of foods implicated in the Ficus-fruit syndrome. Clin. Exp. Allergy 2004, 34, 1251–1258. [Google Scholar] [CrossRef]
- Gandolfo, M.; Baeza, M.; De Barrio, M. Anaphylaxis after eating figs. Allergy 2001, 56, 462–463. [Google Scholar] [CrossRef]
- Englund, P.T.; King, T.P.; Craig, L.C.; Walti, A. Studies on ficin. I. Its isolation and characterization. Biochemistry 1968, 7, 163–175. [Google Scholar] [CrossRef]
- Morellon-Sterling, R.; El-Siar, H.; Tavano, O.L.; Berenguer-Murcia, Á.; Fernández-Lafuente, R. Ficin: A protease extract with relevance in biotechnology and biocatalysis. Int. J. Biol. Macromol. 2020, 162, 394–404. [Google Scholar] [CrossRef]
- Liburdi, K.; Boselli, C.; Giangolini, G.; Amatiste, S.; Esti, M. An Evaluation of the Clotting Properties of Three Plant Rennets in the Milks of Different Animal Species. Foods 2019, 8, 600. [Google Scholar] [CrossRef]
- Mariani, M.; Camagna, M.; Tarditi, L.; Seccamani, E. A new enzymatic method to obtain high-yield F(ab)2 suitable for clinical use from mouse IgGl. Mol. Immunol. 1991, 28, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Ling, X.J.; Zhou, Y.J.; Yang, Y.S.; Xu, Z.Q.; Wang, Y.; Sun, J.L.; Zhu, Y.; Wei, J.F. A new cysteine protease allergen from Ambrosia trifida pollen: Proforms and mature forms. Mol. Immunol. 2022, 147, 170–179. [Google Scholar] [CrossRef]
- Höllbacher, B.; Schmitt, A.O.; Hofer, H.; Ferreira, F.; Lackner, P. Identification of Proteases and Protease Inhibitors in Allergenic and Non-Allergenic Pollen. Int. J. Mol. Sci. 2017, 18, 1199. [Google Scholar] [CrossRef]
- Gaspar, R.; de Matos, M.R.; Cortes, L.; Nunes-Correia, I.; Todo-Bom, A.; Pires, E.; Veríssimo, P. Pollen Proteases Play Multiple Roles in Allergic Disorders. Int. J. Mol. Sci. 2020, 21, 3578. [Google Scholar] [CrossRef]
- Wan, H.; Winton, H.L.; Soeller, C.; Tovey, E.R.; Gruenert, D.C.; Thompson, P.J.; Stewart, G.A.; Taylor, G.W.; Garrod, D.R.; Cannell, M.B.; et al. Der p 1 facilitates transepithelial allergen delivery by disruption of tight junctions. J. Clin. Investig. 1999, 104, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Turk, V.; Bode, W. The cystatins: Protein inhibitors of cysteine proteinases. FEBS Lett. 1991, 285, 213–219. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Allergen | Biochemical Name/EC Number | Allergen Source | MW (SDS-PAGE) | Route of Exposure | Gastro-Intestinal Stability | Substrate (from Enzyme Nomenclature Database [10] or from the Indicated Bibliography) a | Examples of Biotechnological Applications b |
|---|---|---|---|---|---|---|---|---|
| ALLERGENIC PLCP FROM FOODS, POLLEN AND MITES | ||||||||
| Actinidia deliciosa (green kiwifruit) | Act d 1 [11] | Actinidin/EC 3.4.22.14 | Food | 30 | Food | Stable [12,13] | Specificity close to that of papain. It disrupts the epithelial barrier of human intestinal T84 cells by degrading the tight junction protein occludin [14]. It hydrolyses collagen and fibrinogen [15,16]. | Meat tenderisation [17]. Isolation of cells following collagenolytic digestion [15]. |
| Ananas comosus (pineapple) | Ana c 2 [18] | Bromelain/EC 3.4.22.32 | Food | 22.8 | Food, inhalation [18] | Unstable [19] | Hydrolysis of proteins with broad specificity for peptide bonds. Details can be found at www.enzyme-database.org/query.php?ec=3.4.22.32, accessed on 2 January 2024 | Therapeutic and cosmetic use [17,20,21,22]. Meat tenderisation [17]. |
| Carica papaya (papaya) | Cari p 2 [23] | Chymopapain/EC 3.4.22.6 | Food | 28 | Food, inhalation [23] | ND | Specificity similar to that of papain | Allergenicity limits its application [24]. |
| Ambrosia artemisiifolia (short ragweed) | Amb a 11 [25] | Cysteine protease | Pollen | 37 kDa (mature protein), 52 kDa (zymogen) | Airway | ND | Not found | |
| Blomia tropicalis (storage mite) | Blo t 1 [26] | Cysteine protease | Mite | 39 | Airway | ND | Not found | |
| Dermatophagoides farinae (american house dust mite) | Der f 1 [27] | Cysteine protease/EC 3.4.22.65 | Mite | 27 | Airway, ingestion [28] | ND | Not found | |
| Dermatophagoides microceras (house dust mite) | Der m 1 [29] | Cysteine protease | Mite | 25 | Airway | ND | Refers to Der p 1 | |
| Dermatophagoides pteronyssinus (european house dust mite) | Der p 1 [30] | Cysteine protease/EC 3.4.22.65 | Mite | 24 | Airway, ingestion [28] | ND | Details are available at www.enzyme-database.org/query.php?ec=3.4.22.65, accessed on 2 January 2024 | |
| Euroglyphus maynei (house dust mite) | Eur m 1 [31] | Cysteine protease/EC 3.4.22.65 | Mite | Airway | ND | Not found | ||
| Tyrophagus putrescentiae (storage mite) | Tyr p 1 [32] | Cysteine protease | Mite | 25 | Airway | ND | Not found | |
| POTENTIALLY ALLERGENIC PLCP FROM FOODS, NOT (YET) REGISTERED BY WHO-IUIS | ||||||||
| Ficus carica (fig) | Fic c Ficin [9] | Ficin/EC 3.4.22.3 | Food | 24 [33] | Food, occupational | Unstable [33] need encapsulation | Specificity similar to that of papain | Meat tenderisation [17]. Cheese and milk protein hydrolysates for special foods production [34]. |
| Carica papaya (papaya) | Cari p papain [9] | Papain/EC 3.4. 22.2 | Food | 24 [33] | Food, inhalation, skin contact [35,36] | Unstable [37] need encapsulation | Hydrolysis of proteins with broad specificity for peptide bonds, but preference for an amino acid bearing a large hydrophobic side chain at the P2 position. Does not accept Val in P1′. | Meat tenderization [17,38]. Exfoliantig agent [38]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giangrieco, I.; Ciardiello, M.A.; Tamburrini, M.; Tuppo, L.; Mari, A.; Alessandri, C. Plant and Arthropod IgE-Binding Papain-like Cysteine Proteases: Multiple Contributions to Allergenicity. Foods 2024, 13, 790. https://doi.org/10.3390/foods13050790
Giangrieco I, Ciardiello MA, Tamburrini M, Tuppo L, Mari A, Alessandri C. Plant and Arthropod IgE-Binding Papain-like Cysteine Proteases: Multiple Contributions to Allergenicity. Foods. 2024; 13(5):790. https://doi.org/10.3390/foods13050790
Chicago/Turabian StyleGiangrieco, Ivana, Maria Antonietta Ciardiello, Maurizio Tamburrini, Lisa Tuppo, Adriano Mari, and Claudia Alessandri. 2024. "Plant and Arthropod IgE-Binding Papain-like Cysteine Proteases: Multiple Contributions to Allergenicity" Foods 13, no. 5: 790. https://doi.org/10.3390/foods13050790
APA StyleGiangrieco, I., Ciardiello, M. A., Tamburrini, M., Tuppo, L., Mari, A., & Alessandri, C. (2024). Plant and Arthropod IgE-Binding Papain-like Cysteine Proteases: Multiple Contributions to Allergenicity. Foods, 13(5), 790. https://doi.org/10.3390/foods13050790