Effects of Dark Chocolate Intake on Brain Electrical Oscillations in Healthy People


Abstract
:1. Introduction
2. Materials and Methods
2.1. Baseline EEG Recording
2.2. Experimental Maneuver
2.3. Statistical Mnalysis
3. Results
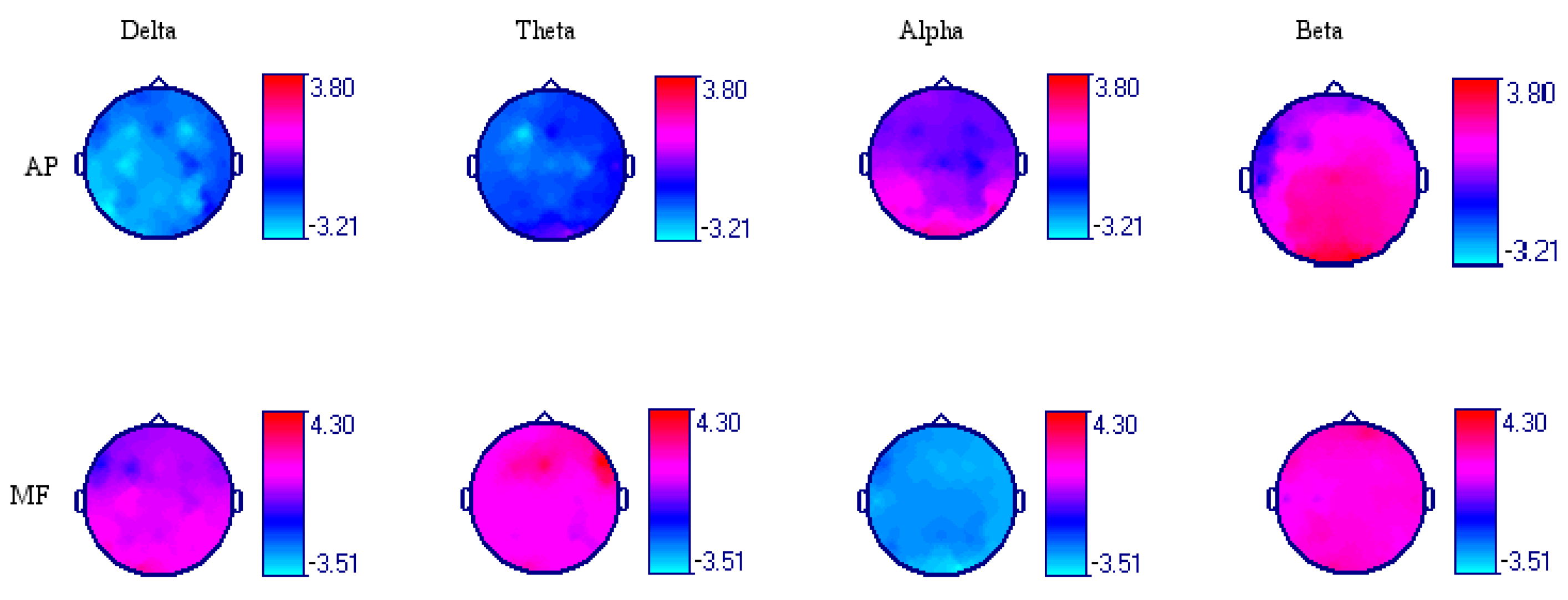
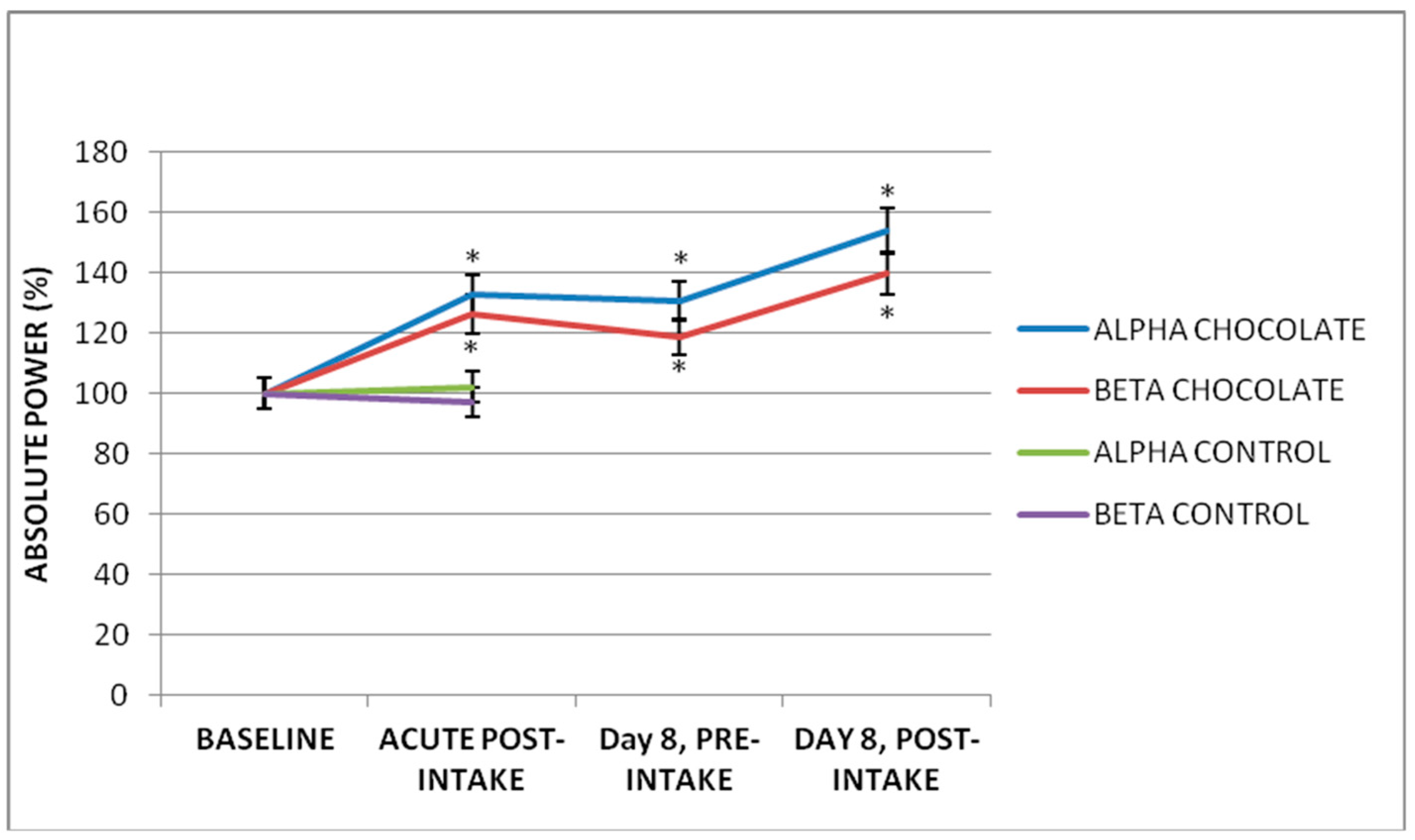
3.1. Acute Effect of Dark Chocolate Intake
3.2. Acute Effect of Low-Calorie Milk Intake
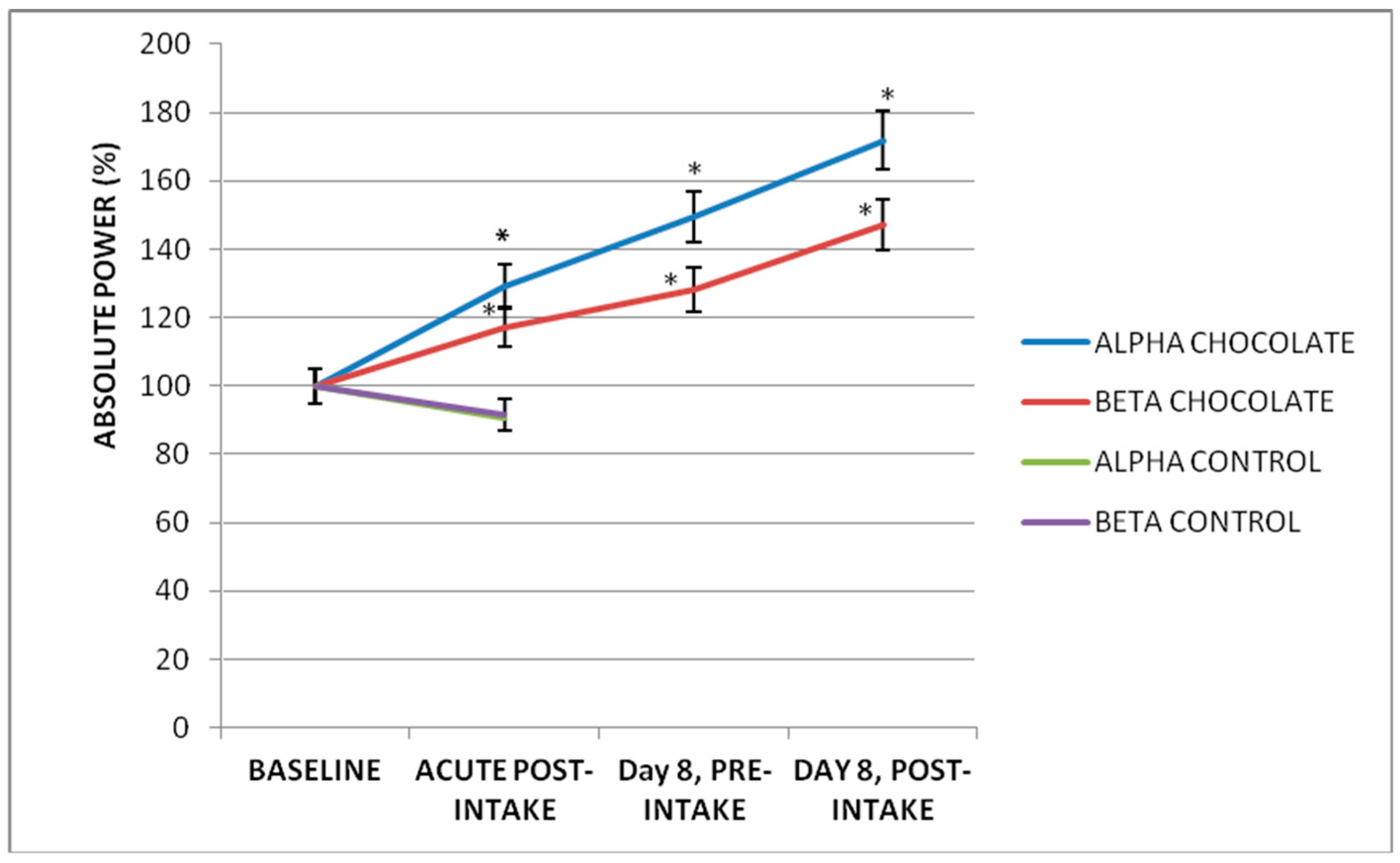
3.3. Subchronic Effect of Seven-Day Dark Chocolate Intake
3.4. Acute Effect on a Subchronic Effect of Dark Chocolate Intake
3.5. Modifications of Mean Frequency
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kalaria, R.N.; Maestre, G.E.; Arizaga, R.; Friedland, R.P.; Galasko, D.; Hall, K.; Luchsinger, J.A.; Ogunniyi, A.; Perry, E.K.; Potocnik, F.; et al. Alzheimer’s disease and vascular dementia in developing countries: Prevalence, management, and risk factors. Lancet Neurol. 2008, 7, 812–826. [Google Scholar] [CrossRef]
- Linsell, L.; Malouf, R.; Morris, J.; Kurinczuk, J.J.; Marlow, N. Prognostic factors for poor cognitive development in children born very preterm or with very low birth weight: A systematic review. JAMA Pediatr. 2015, 169, 1162–1172. [Google Scholar] [CrossRef] [PubMed]
- Kehagia, A.A. A neurological perspective on the enhancement debate: Lessons learned from Parkinson’s disease. J. Psychopharmacol. 2016, 30, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Magrone, T.; Russo, M.A.; Jirillo, E. Cocoa and dark chocolate polyphenols: From biology to clinical applications. Front. Immunol. 2017, 8, 677. [Google Scholar] [CrossRef] [PubMed]
- Desideri, G.; Kwik-Uribe, C.; Grassi, D.; Necozione, S.; Ghiadoni, L.; Mastroiacovo, D.; Raffaele, A.; Ferri, L.; Bocale, R.; Lechiara, M.C.; et al. Benefits in cognitive function, blood pressure, and insulin resistance through cocoa flavanol cognition and aging (CoCoA) study. Hypertension 2012, 60, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Brickman, A.M.; Khan, U.A.; Provenzano, F.A.; Yeung, L.K.; Suzuki, W.; Schroeter, H. Enhancing dentate gyrus function with dietary flavanols improves cognition in older adults. Nat. Neurosci. 2014, 17, 1798–1803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Socci, V.; Tempesta, D.; Desideri, G.; De Gennaro, L.; Ferrara, M. Enhancing human cognition with cocoa flavonoids. Front. Nutr. 2017, 16, 4–19. [Google Scholar] [CrossRef] [PubMed]
- John, E.R.; Karmel, B.Z.; Corning, W.C.; Easton, P.; Brown, D.; Ahn, H. Neurometrics. Science 1977, 196, 1393–1410. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, R.W.; North, D.M.; Biver, C.J. Intelligence and EEG phase reset: A two compartmental model of phase shift and lock. Neuroimage 2008, 42, 1639–1653. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, R.W.; Palmero-Soler, E.; North, D.M.; Biver, C.J. Intelligence and EEG measuresof information flow: Efficiency and homeostatic neuroplasticity. Sci. Rep. 2016, 6, 38890. [Google Scholar] [CrossRef] [PubMed]
- Hernández, J.L.; Valdés, P.; Biscay, R.; Virues, T.; Szava, S.; Bosch, J. A global scale factor in brain topography. Int. J. Neurosci. 1994, 76, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Camfield, P.; Camfield, C.; Pohlmann-Eden, B. Transition from pediatric to adult epilepsy care: A difficult process marked by medical and social crisis. Epilepsy Curr. 2012, 12, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Francis, S.T.; Head, K.; Morris, P.G.; Macdonald, I.A. The effect of flavanol-rich cocoa on the fMRI response to a cognitive task in healthy young people. J. Cardiovasc. Pharmacol. 2006, 47 (Suppl. 2), S215–S220. [Google Scholar] [CrossRef] [PubMed]
- Decroix, L.; Tonoli, C.; Soares, D.D.; Tagougui, S.; Heyman, E.; Meeusen, R. Acute cocoa flavanol improves cerebral oxygenation without enhancing executive function at rest or after exercise. Appl. Physiol. Nutr. Metab. 2016, 41, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Scholey, A.B.; French, S.J.; Morris, P.J.; Kennedy, D.O.; Milne, A.L.; Haskell, C.F. Consumption of cocoa flavanols results in improvements in mood and cognitive performance during sustained mental effort. J. Psychopharmacol. 2010, 24, 1505–1514. [Google Scholar] [CrossRef] [PubMed]
- Field, D.T.; Williams, C.M.; Butler, L.T. Consumption of cocoa flavanols results in an improvement in visual and cognitive functions. Physiol. Behav. 2011, 103, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Pase, M.P.; Scholey, A.B.; Pipingas, A.; Kras, M.; Nolidin, K.; Gibbs, A. Cocoa polyphenols enhance positive mood states but not cognitive performance: A randomized, placebo-controlled trial. J. Psychopharmacol. 2013, 27, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Grassi, D.; Lippi, C.; Necozione, S.; Desideri, G.; Ferri, C. Short-term administration of dark chocolate is followed by a significant increase in insulin sensitivity and a decrease in blood pressure in healthy persons. Am. J. Clin. Nutr. 2005, 81, 611–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datla, K.P.; Christidou, M.; Widmer, W.W.; Rooprai, H.K.; Dexter, D.T. Tissue distribution and neuroprotective effects of citrus flavonoid tangeretin in a rat model of Parkinson’s disease. Neuroreport 2001, 12, 3871–3875. [Google Scholar] [CrossRef] [PubMed]
- Fernell, M.; Swinton, C.; Lukowiak, K. Epicatechin, a component of dark chocolate, enhances memory formation if applied during the memory consolidation period. Commun. Integr. Biol. 2016, 9, 1205772–1205774. [Google Scholar] [CrossRef] [PubMed]
- Cicvaric, A.; Bulat, T.; Bormann, D.; Yang, J.; Auer, B.; Milenkovic, I. Sustained consumption of cocoa-based dark chocolate enhances seizure-like events in the mouse hippocampus. Food Funct. 2018, 9, 1532–1544. [Google Scholar] [CrossRef] [PubMed]
- Jensen, O.; Gelfand, J.; Kounios, J.; Lisman, J.E. Oscillations in the alpha band (9–12 Hz) increase with memory load during retention in a short-term memory task. Cereb. Cortex 2002, 12, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Poch, C.; Valdivia, M.; Capilla, A.; Hinojosa, J.A.; Campo, P. Suppression of no-longer relevant information in working memory: An alpha-power related mechanism? Biol. Psychol. 2018, 135, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Mierau, A.; Klimesch, W.; Lefebvre, J. State-dependent alpha peak frequency shifts: Experimental evidence, potential mechanisms and functional implications. Neuroscience 2017, 360, 146–154. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leads | AP Delta | AP Theta | AP Alpha | AP Beta |
|---|---|---|---|---|
| Fp1-AVR | −1.566 | −1.906 * | 0.332 | 0.352 |
| Fp2-AVR | −2.073 * | −1.409 | −0.122 | 0.042 |
| F3-AVR | −2.795 * | −3.035 * | −0.349 | 0.128 |
| F4-AVR | −2.905 * | −1.323 | −0.521 | 1.681 |
| C3-AVR | −2.994 * | −2.108 * | 0.295 | 2.027 * |
| C4-AVR | −1.378 | −2.196 * | −0.791 | 2.110 * |
| P3-AVR | −2.172 * | −1.431 | 1.407 | 2.494 * |
| P4-AVR | −2.776 * | −1.204 | 0.058 | 2.191 * |
| O1-AVR | −2.680 * | −0.395 | 2.557 * | 3.356 * |
| O2-AVR | −1.796 * | 0.318 | 2.061 * | 3.800 * |
| F7-AVR | −1.499 | −1.809 * | −0.168 | −1.000 |
| F8-AVR | −1.559 | −1.273 | −0.177 | 0.66 |
| T3-AVR | −2.817 * | −1.967 * | 0.443 | −0.915 |
| T4-AVR | −1.509 | −0.397 | 0.076 | 1.741 * |
| T5-AVR | −3.213 * | −1.541 | 1.768 * | 1.381 |
| T6-AVR | −0.878 | −1.222 | 2.134 * | 2.088 * |
| FZ-AVR | −1.577 | −0.878 | 0.000 | 1.663 |
| CZ-AVR | −2.354 * | −2.154 * | −0.595 | 2.528 * |
| PZ-AVR | −2.607 * | −1.851 * | 0.312 | 2.493 * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santiago-Rodríguez, E.; Estrada-Zaldívar, B.; Zaldívar-Uribe, E. Effects of Dark Chocolate Intake on Brain Electrical Oscillations in Healthy People. Foods 2018, 7, 187. https://doi.org/10.3390/foods7110187
Santiago-Rodríguez E, Estrada-Zaldívar B, Zaldívar-Uribe E. Effects of Dark Chocolate Intake on Brain Electrical Oscillations in Healthy People. Foods. 2018; 7(11):187. https://doi.org/10.3390/foods7110187
Chicago/Turabian StyleSantiago-Rodríguez, Efraín, Brenda Estrada-Zaldívar, and Elba Zaldívar-Uribe. 2018. "Effects of Dark Chocolate Intake on Brain Electrical Oscillations in Healthy People" Foods 7, no. 11: 187. https://doi.org/10.3390/foods7110187
APA StyleSantiago-Rodríguez, E., Estrada-Zaldívar, B., & Zaldívar-Uribe, E. (2018). Effects of Dark Chocolate Intake on Brain Electrical Oscillations in Healthy People. Foods, 7(11), 187. https://doi.org/10.3390/foods7110187