Protection by the Total Flavonoids from Rosa laevigata Michx Fruit against Lipopolysaccharide-Induced Liver Injury in Mice via Modulation of FXR Signaling

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Herbal Material and Preparation of TFs
2.3. Animals
2.4. Determination of Biochemical Parameters in Serum and Liver
2.5. Hematoxylin and Eosin Staining
2.6. Oil Red O Staining
2.7. Transmission Electron Microscopy (TEM) Assay
2.8. Immunohistochemical and Immunofluorescent Assays
2.9. Quantitative Real-Time PCR Assay
2.10. Western Blotting Assay
2.11. Statistical Analysis
3. Results
3.1. Effects of TFs on LPS-Induced Liver Injury
3.2. TFs Improve LPS-Induced Cellular Structure Changes in Mice
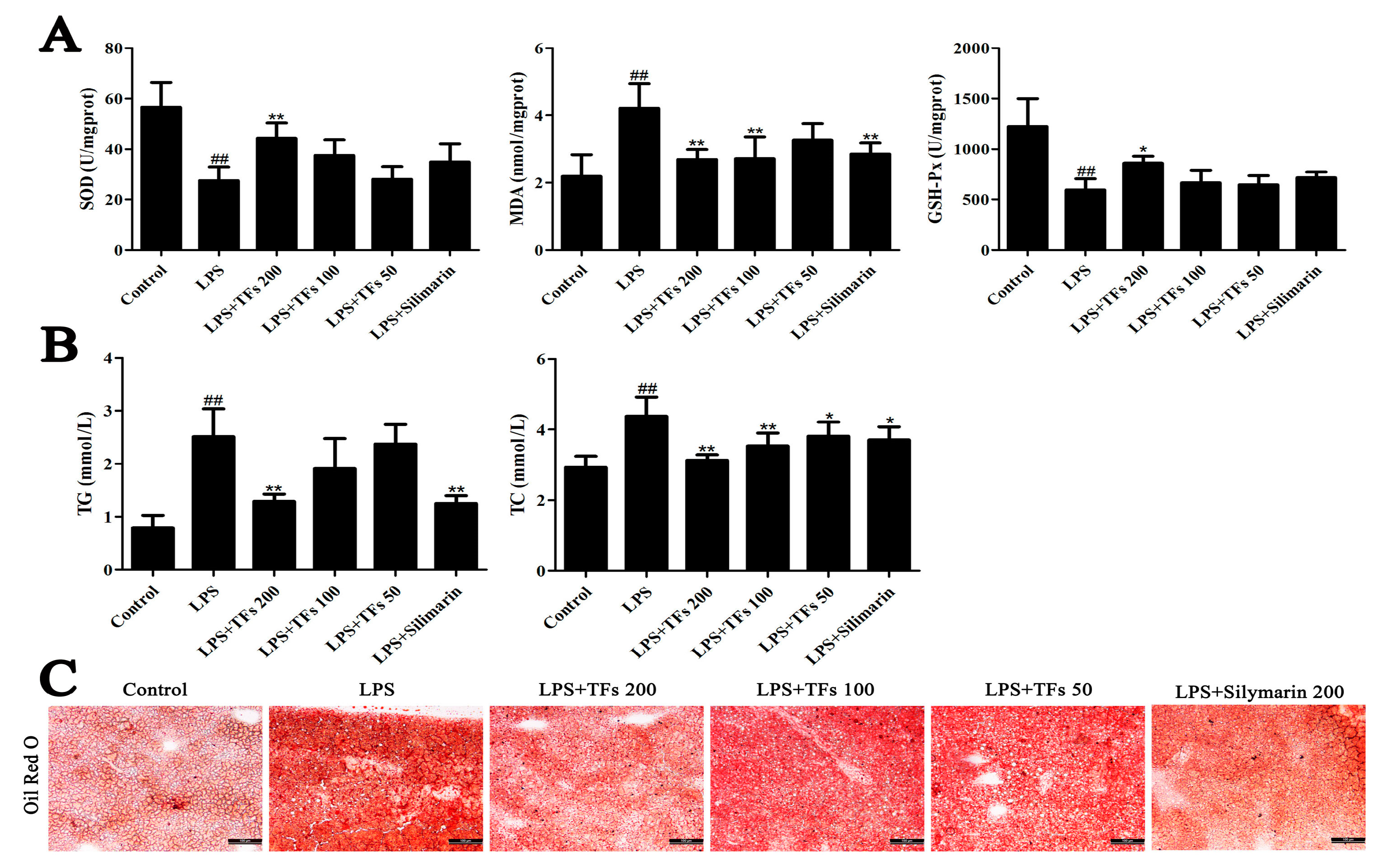
3.3. TFs Suppresse LPS-Induced Oxidative Liver Injury
3.4. Effects of TFs on Lipid Metabolism
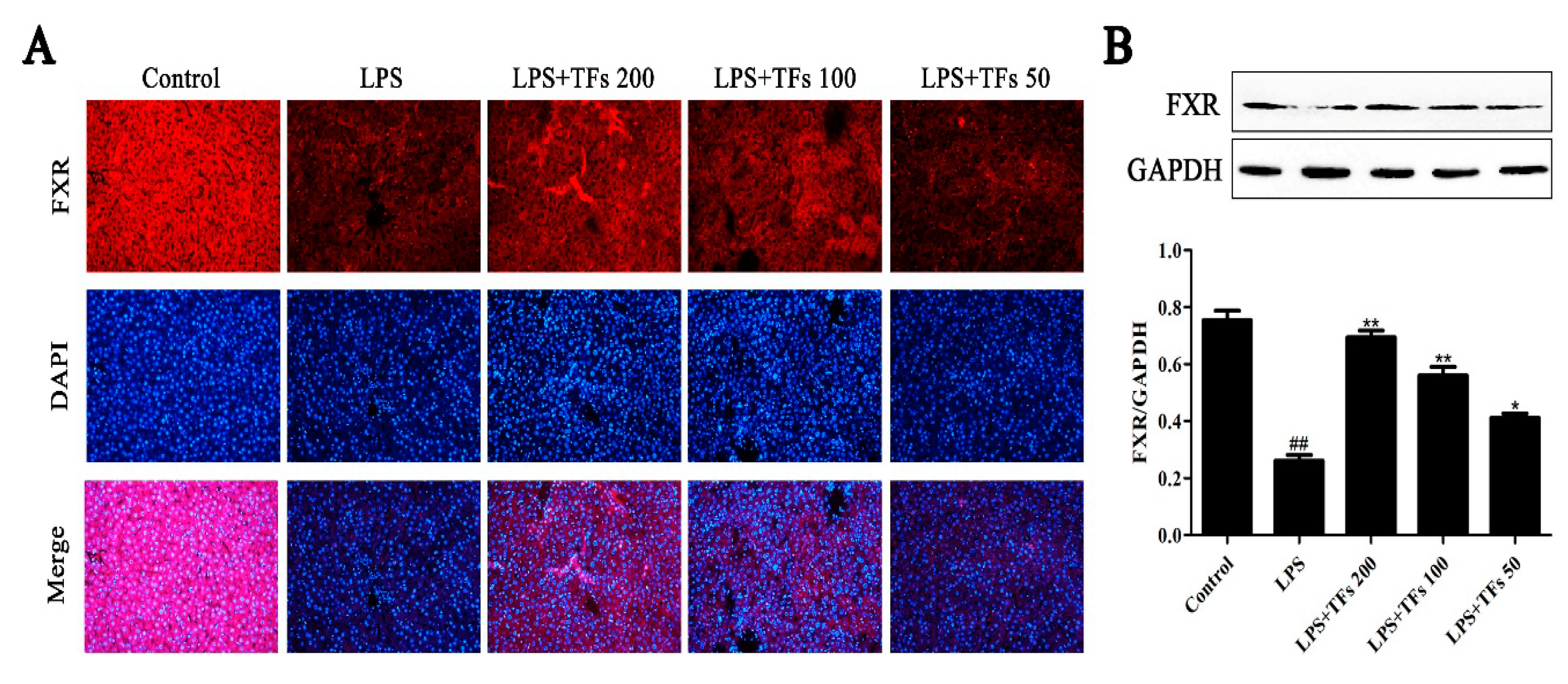
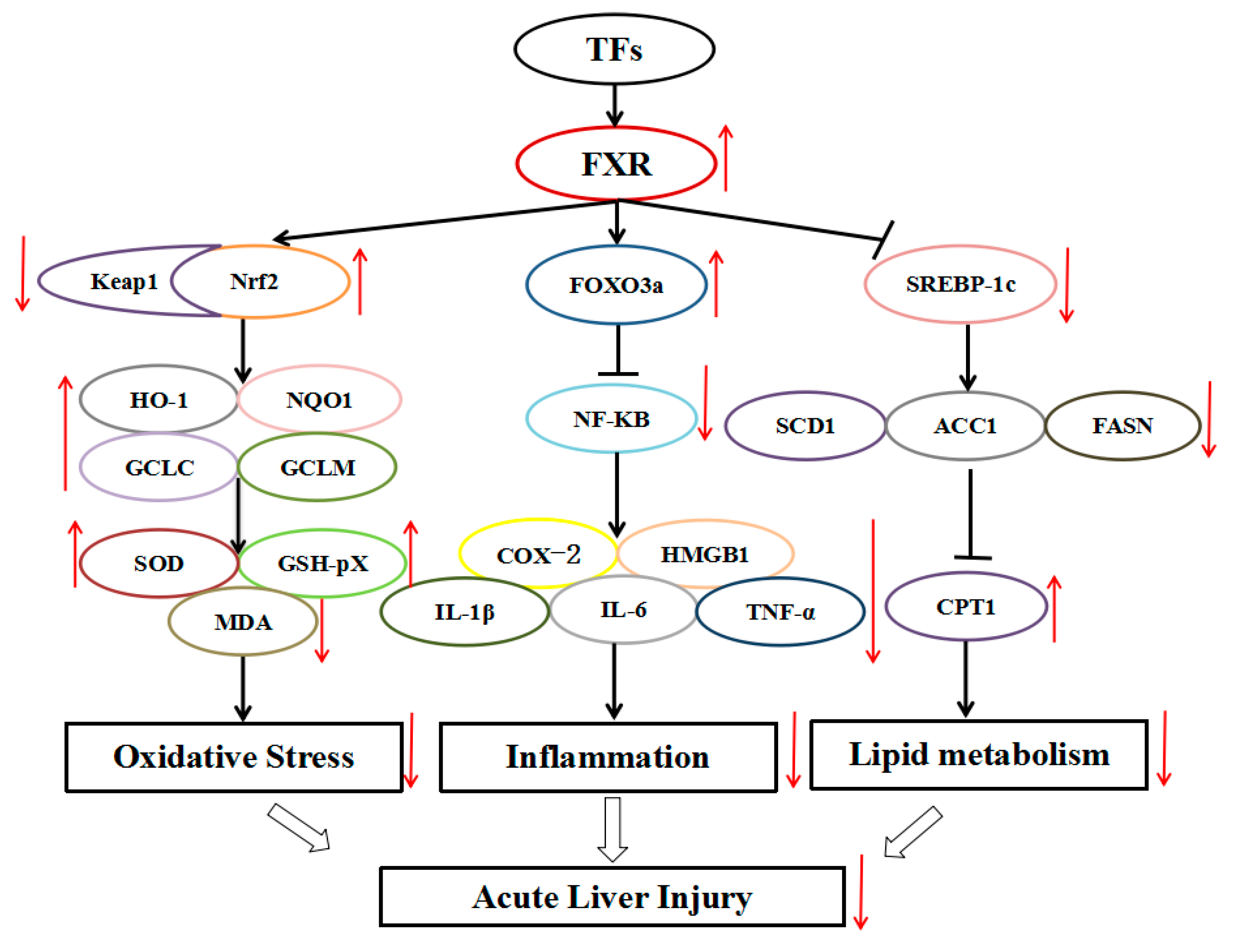
3.5. TFs Activate the FXR Signal Pathway
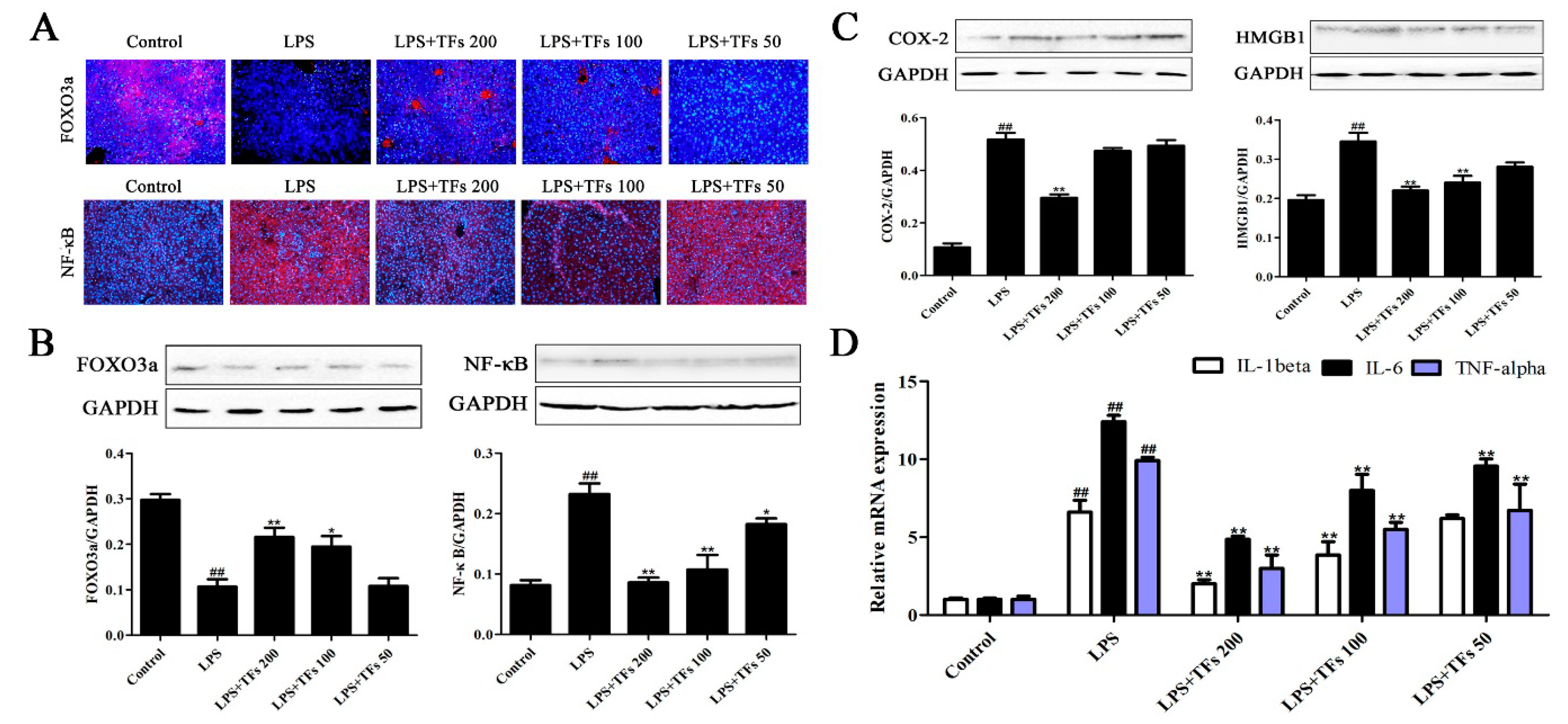
3.6. TFs Regulate FXR-Nrf2-Mediated Oxidative Stress
3.7. TFs Regulate FXR-FOXO3a-Mediated Inflammation
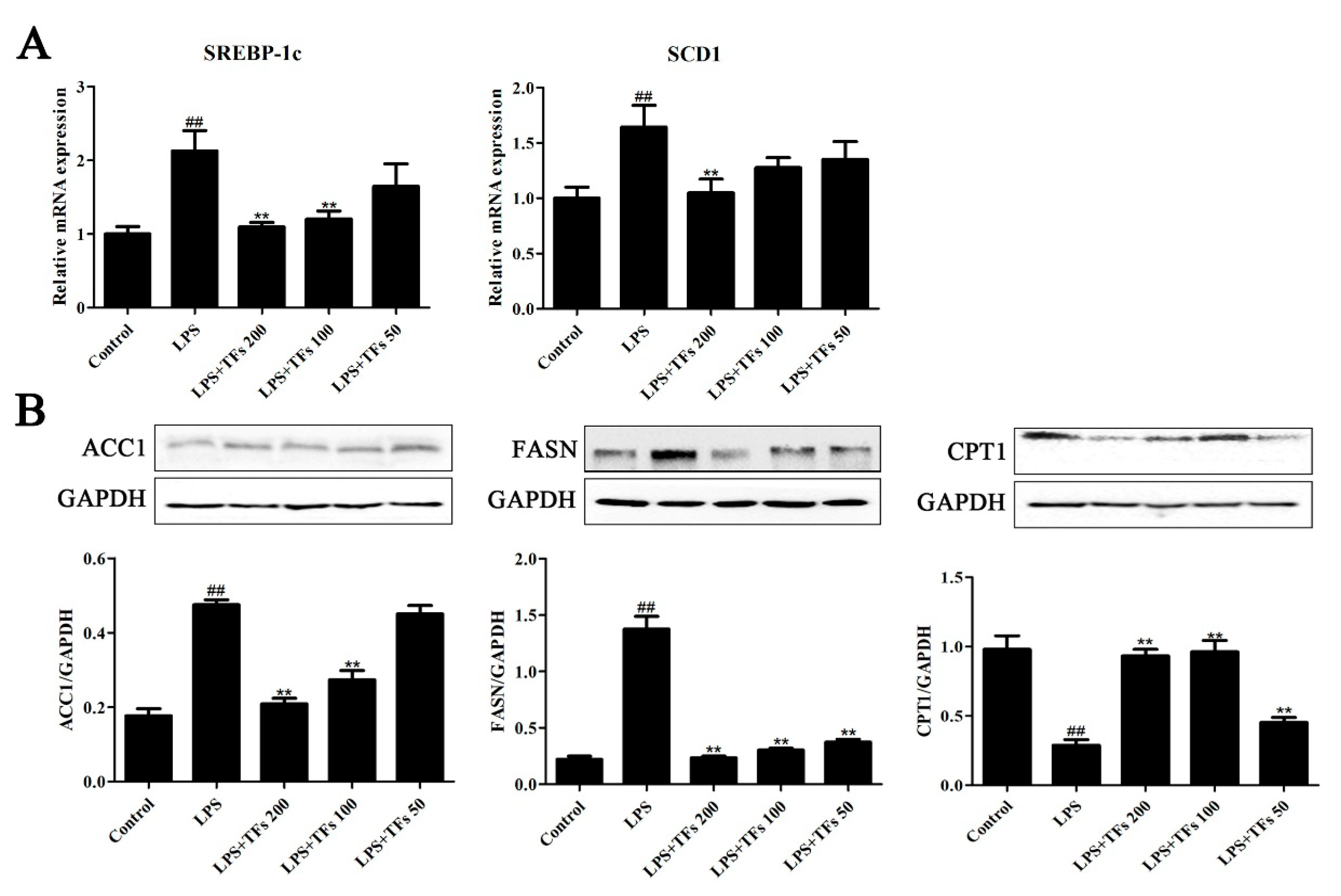
3.8. TFs Regulats FXR-SREBP-1c-Mediated Lipid Metabolism
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACC1 | acetyl-CoA carboxylase-1 |
| ALI | acute liver injury |
| ALT | alanine transferase |
| AST | aspartate transaminase |
| COX2 | cyclooxygenase-2 |
| CPT1 | carnitine palmitoyltransferase 1 |
| FASN | fatty acid synthase |
| FOXO3a | forkhead box O3 |
| FXR | farnesoid X receptor |
| GAPDH | glyceraldehyde-3-phosphatedehydrogenase |
| GCLC | glutamate-cysteine ligasecatalytic subunit |
| GCLM | glutamate-cysteine ligase regulatory subunit |
| GSH-Px | glutathione peroxidase |
| HMGB-1 | high-mobility group box 1 |
| HO-1 | heme oxygenase-1 |
| IL-1β | interleukin-1 β |
| Keap1 | Kelch-likeECH-associated protein 1 |
| MDA | malondialdehyde |
| NF-κB | nuclear factor-kappa B |
| Nrf2 | nuclear erythroid factor2-related factor 2 |
| NQO1 | NAD(P)H dehydrogenase(quinone 1) |
| SCD1 | stearoyl-coenzyme A desaturase 1 |
| SOD | superoxide dismutase |
| SREBP-1c | sterol regulatory element-binding proteins-1c |
| TC | total cholesterol |
| TFs | total flavonoids from Rosa laevigata Michx fruit |
| TG | total triglyceride |
| TNF-α | tumor necrosis factor alpha |
References
- Nolan, J.P. The role of intestinal endotoxin in liver injury: A long and evolving history. Hepatology 2010, 52, 1829–1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emlet, D.R.; Shaw, A.D.; Kellum, J.A. Sepsis-associated AKI: Epithelial cell dysfunction. Semin. Nephrol. 2015, 35, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Davis, B.C.; Tillman, H.; Chung, R.T.; Stravitz, R.T.; Reddy, K.R.; Fontana, R.J.; McGuire, B.; Davern, T.; Lee, W.M. Heat Stroke Leading to Acute Liver Injury & Failure: A Case Series from the Acute Liver Failure Study Group. Liver Int. 2017, 37, 509–513. [Google Scholar] [PubMed]
- Stoyanoff, T.R.; Todaro, J.S.; Aguirre, M.V.; Zimmermann, M.C.; Brandan, N.C. Amelioration of lipopolysaccharide-induced acute kidney injury by erythropoietin: involvement of mitochondria- regulated apoptosis. Toxicology 2014, 318, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.H.; Yang, J.J.; Yang, M.L.; Li, Y.C.; Kuan, Y.H. Rutin decreases lipopolysaccharide-induced acute lung injury via inhibition of oxidative stress and the MAPK-NF-kappaB pathway. Free Radic. Biol. Med. 2014, 69, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Jawan, B.; Kao, Y.H.; Goto, S.; Pan, M.C.; Lin, Y.C.; Hsu, L.W.; Nakano, T.; Lai, C.Y.; Sun, C.K.; Cheng, Y.F.; et al. Propofol pretreatment attenuates LPS-induced granulocyte-macrophage colony-stimulating factor production in cultured hepatocytes by suppressing MAPK/ERK activity and NF-kappaB translocation. Toxicol. Appl. Pharmacol. 2008, 229, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.G.; Yang, J.H.; Kim, K.M.; Jang, C.H.; Jung, J.Y.; Cho, I.J.; Shin, S.M.; Ki, S.H. Regulation of Toll-like Receptor-mediated Sestrin2 Induction by AP-1, Nrf2, and the Ubiquitin-Proteasome System in Macrophages. Toxicol. Sci. 2015, 144, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ding, H.W.; Li, H.D.; Huang, H.M.; Li, X.F.; Yang, Y.; Zhang, Y.L.; Pan, X.Y.; Huang, C.; Meng, X.M. Hesperetin derivative-14 alleviates inflammation by activating PPAR-γ in mice with CCl4-induced acute liver injury and LPS-treated RAW264.7 cells. Toxicol. Lett. 2017, 274, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Ning, Y.X.; Kim, J.K.; Min, H.K.; Ren, S.L. Cholesterol Metabolites Alleviate Injured Liver Function and Decrease Mortality in an LPS-Induced Mouse Model. Metabolism 2016, 71, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Honke, N.; Shaabani, N.; Hardt, C.; Krings, C.; Häussinger, D.; Lang, P.A.; Lang, K.S.; Keitel, V. Farnesoid X Receptor in Mice Prevents Severe Liver Immunopathology During Lymphocytic Choriomeningitis Virus Infection. Cell Physiol. Biochem. 2017, 41, 323–338. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.M.; Xu, Y.W.; Qi, Y.; Xu, L.N.; Song, S.S.; Yin, L.H.; Tao, X.F.; Zhen, Y.H.; Han, X.; Ma, X.D.; et al. Protective effects of dioscin against doxorubicin-induced nephrotoxicity via adjusting FXR-mediated oxidative stress and inflammation. Toxicology 2017, 378, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Manley, S.; Ni, H.M.; Williams, J.A.; Kong, B.; DiTacchio, L.; Guo, G.; Ding, W.X. Farnesoid X receptor regulates forkhead Box O3a activation in ethanol-induced autophagy and hepatotoxicity. Redox Biol. 2014, 2, 991–1002. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Duan, X.P.; Wang, C.P.; Liu, Z.H.; Sun, P.Y.; Huo, X.K.; Sun, H.J.; Peng, J.Y.; Liu, K.X. Alisol B 23-acetate protects against non-alcoholic steatohepatitis in mice via farnesoid X receptor activation. Acta Pharmacol. Sin. 2017, 38, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.X.; Liu, X.X.; Yuan, Z.H.; Li, X.J.Y.; Yang, H.; Yuan, Z.Q.; Sun, X.L.; Zhang, L.Y.; Jiang, Z.Z. SRT1720 Alleviates ANIT-Induced Cholestasis in a Mouse Model. Front. Pharmacol. 2017, 8, 256. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Hron, J.D.; Peng, S.L. Regulation of NF-κB, Th Activation, and Autoinflammation by the Forkhead Transcription Factor Foxo3a. Immunity 2004, 21, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Mencarelli, A.; Renga, B.; D’Amore, C.; Santorelli, C.; Graziosi, L.; Bruno, A.; Monti, M.C.; Distrutti, E.; Cipriani, S.; Donini, A.; et al. Dissociation of Intestinal and Hepatic Activities of FXR and LXRα Supports Metabolic Effects of Terminal Ileum Interposition in Rodents. Diabetes 2013, 62, 3384–3393. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.T.; Lu, B.N.; Peng, J.Y. Hepatoprotective activity of the total flavonoids from Rosa laevigata Michx fruit in mice treated by paracetamol. Food Chem. 2011, 125, 719–725. [Google Scholar] [CrossRef]
- Dong, D.S.; Qi, Y.; Xu, L.N.; Yin, L.H.; Xu, Y.W.; Han, X.; Zhao, Y.Y.; Peng, J.Y. Total saponins from Rosa laevigata Michx fruit attenuates hepatic steatosis induced by high-fat diet in rats. Food Funct. 2014, 5, 3065–3075. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.S.; Xu, L.N.; Tao, X.F.; Han, X.; Yin, L.H.; Qi, Y.; Peng, J.Y. Protective Effect of the Total Flavonoids from Rosa laevigata Michx Fruit on Renal Ischemia-Reperfusion Injury through Suppression of Oxidative Stress and Inflammation. Molecules 2016, 21, 952. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Lu, B.N.; Han, X.; Xu, L.N.; Qi, Y.; Yin, L.H.; Xu, Y.W.; Zhao, Y.Y.; Liu, K.X.; Peng, J.Y. Protection of the flavonoid fraction from Rosa laevigata Michx fruit againstcarbon tetrachloride-induced acute liver injury in mice. Food Chem. Toxicol. 2013, 55, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zheng, L.L.; Dong, D.S.; Xu, L.N.; Yin, L.H.; Qi, Y.; Han, X.; Lin, Y.; Liu, K.X.; Peng, J.Y. Effects of flavonoids from Rosa laevigata Michx fruit against high-fatdiet-induced non-alcoholic fatty liver disease in rats. Food Chem. 2013, 141, 2108–2116. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.F.; Sun, X.C.; Xu, L.N.; Yin, L.H.; Han, X.; Qi, Y.; Xu, Y.W.; Zhao, Y.Y.; Wang, C.Y.; Peng, J.Y. Total Flavonoids from Rosa laevigata Michx Fruit Ameliorates Hepatic Ischemia/Reperfusion Injury through Inhibition of Oxidative Stress and Inflammation in Rats. Nutrients 2016, 8, 418. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zheng, L.L.; Xu, L.N.; Sun, H.J.; Li, H.; Yao, J.H.; Liu, K.X.; Peng, J.Y. Subchronic toxicity study of the total flavonoids from Rosa laevigata Michx fruitin rats. Regul. Toxicol. Pharmacol. 2012, 62, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.T.; Lu, B.N.; Xu, L.N.; Yin, L.H.; Wang, X.N.; Peng, J.Y.; Liu, K.X. The antioxidant activity and hypolipidemic activity of the total flavonoids from the fruit of Rosa laevigata Michx. Nat Sci. 2010, 2, 175–183. [Google Scholar] [CrossRef]
- Yao, H.; Hu, C.S.; Yin, L.H.; Tao, X.F.; Xu, L.N.; Qi, Y.; Han, X.; Xu, Y.W.; Zhao, Y.Y.; Wang, C.Y.; et al. Dioscin reduces lipopolysaccharide-induced inflammatory liver injury via regulating TLR4/MyD88 signal pathway. Int. Immunopharmacol. 2016, 36, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.M.; Li, H.D.; Chen, X.; Yang, Y.; Li, X.F.; Li, W.X.; Huang, C.; Meng, X.M.; Zhang, L.; Li, J. HMGA2, a driver of inflammation, is associated with hypermethylation in acute liver injury. Toxicol. Appl. Pharmacol. 2017, 328, 34–35. [Google Scholar] [CrossRef] [PubMed]
- Ranaa, M.; Reddya, S.S.; Mauryaa, P.; Singha, V.; Chaturvedia, S.; Kaura, K.; Agarwala, H.; Ahmadb, H.; Naqvib, A.; Dwivedib, A.K.; et al. Turmerone enriched standardized Curcuma longa extract alleviates LPS induced inflammation and cytokine production by regulating TLR4-IRAK1-ROS-MAPK-NFκB axis. J. Funct. Foods 2015, 16, 152–163. [Google Scholar] [CrossRef]
- Yoo, S.; Kim, M.Y.; Cho, J.Y. Beauvericin, a cyclic peptide, inhibits inflammatory responses in macrophages by inhibiting the NF-κB pathway. Korean J. Physiol. Pharmacol. 2017, 21, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.; Ubaid, N.; Qureshi, T.M.; Munir, M.; Mehmood, A. Effect of ultrasound and chemical treatment on total phenol, flavonoids and antioxidant properties on carrot-grape juice blend during storage. Ultrason. Sonochem. 2018, 45, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Pereira, G.A.; Arruda, H.S.; de Morais, D.R.; Eberlin, M.N.; Pastore, G. Carbohydrates, volatile and phenolic compounds composition, and antioxidant activity of calabura (Muntingia calabura L.) fruit. Food Res. Int. 2018, 108, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.H.K.; Baqueta, M.R.; Coqueiro, A.; Dias, M.I.; Barros, L.; Barreiro, M.F.; Ferreira, I.C.F.R.; Gonçalves, O.H.; Bona, E.; da Silva, M.V.; Leimann, F.V. Systematic study on the extraction of antioxidants from pinhão (Araucaria angustifolia (bertol.) Kuntze) coat. Food Chem. 2018, 261, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.L.; Wu, Y.F.; Wang, Y.S.; Wang, X.Z.; Piao, C.H.; Liu, J.M.; Liu, Y.L.; Wang, Y.H. The protective effects of probiotic-fermented soymilk on high-fat diet-induced hyperlipidemia and liver injury. J. Funct. Foods 2017, 30, 220–227. [Google Scholar] [CrossRef]
- Yashaswini, P.S.; Sadashivaiah, B.; Ramaprasad, T.R.; Singh, S.A. In vivo modulation of LPS induced leukotrienes generation and oxidative stress by sesame lignans. J. Nutr. Biochem. 2017, 41, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Anders, L.C.; Lang, A.L.; Anwar-Mohamed, A.; Douglas, A.N.; Bushau, A.M.; Falkner, K.C.; Hill, B.G.; Warner, N.L.; Arteel, G.E.; Cave, M.; et al. Vinyl Chloride Metabolites Potentiate Inflammatory Liver Injury caused by LPS in Mice. Toxicol. Sci. 2016, 151, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.M.; Sonsalla, P.K.; Richardson, J.R. Coordinated role of voltage-gated sodium channels and the Na+/H+ exchanger in sustaining microglial activation during inflammation. Toxicol. Appl. Pharmcol. 2013, 273, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.F.; Zhang, Y.L.; Bai, R.X.; Wang, M.; Du, S.Y. Baicalin Attenuates Alcoholic Liver Injury through Modulation of Hepatic Oxidative Stress, Inflammation and Sonic Hedgehog Pathway in Rats. Cell Physiol. Biochem. 2016, 39, 1129–1140. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.H.; Lei, J.L.; Zhuang, Y.; Zhang., K.; Lu, D.G. Overexpression of HMGB1 A-box reducedIL-1β-induced MMP expression and the production of inflammatory mediators in human chondrocytes. Exp. Cell Res. 2016, 349, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Yamamoto, H.; Nakahashi, O.; Kagawa, T.; Ishiguro, M.; Masuda, M.; Kozai, M.; Ikeda, S.; Taketani, Y.; Takeda, E. Dietary phosphate restriction induces hepatic lipid accumulation through dysregulation of cholesterol metabolism in mice. Nutr. Res. 2013, 33, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.L.; Pathak, P.; Boehme, S.; Chiang, J.Y.L. Cholesterol 7α-hydroxylase protects the liver from inflammation and fibrosis by maintaining cholesterol homeostasis. J. Lipid Res. 2016, 57, 1831–1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.Z.; Huang, G.; Gong, W.; Zhou, P.; Zhao, Y.Y.; Zhang, Y.; Zeng, Y.J.; Gao, M.; Pan, Z.S.; He, F.T. FXR ligands protect against hepatocellular inflammation via SOCS3 induction. Cell Signal. 2012, 24, 1658–1664. [Google Scholar] [CrossRef] [PubMed]
- Pathak, P.; Liu, H.; Boehme, S.; Xie, C.; Krausz, K.W.; Gonzalez, F.; Chiang, J.Y.L. Farnesoid X receptor induces Takeda G-protein receptor 5 cross-talk to regulate bile acid synthesis and hepatic metabolism. J. Biol. Chem. 2017, 292, 11055–11069. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Q.; Zhou, L.L.; Yuen, J.; Birkner, N.; Leppert, V.; O’Day, P.A. Delayed Nrf2-regulated antioxidant gene induction in response to silica nanoparticles. Free Radic. Biol. Med. 2017, 108, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.R.; Donepudi, A.C.; Xu, J.; Wei, W.; Cheng, Q.C.; Driscoll, M.V.; Johnson, D.A.; Johnson, J.A.; Li, X.; Slitt, A.L. Fasting induces nuclear factor E2-related factor 2 and ATP-binding cassette transporters via protein kinase A and Sirtuin-1 in mouse and human. Antioxid. Redox Signal. 2014, 20, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Weerachayaphorn, J.; Cai, S.Y.; Soroka, C.J.; Boyer, J.L. Nuclear factor erythroid 2-related factor 2 is a positive regulator of human bile salt export pump expression. Hepatology 2009, 50, 1588–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, H.J.; Cai, J.; Zhang, L.F.; Peng, Y.X.; Wu, P.; Xie, T.; Pan, Q.J. A novel acute lethal liver injury mouse model with visualization of NF-κB activity for treatment of severe acute liver injury. Am. J. Transl. Res. 2017, 9, 962–970. [Google Scholar] [PubMed]
- Xu, W.X.; Lu, C.F.; Yao, L.; Zhang, F.; Shao, J.J.; Zheng, S.Z. Dihydroartemisinin protects against alcoholic liver injury through alleviating hepatocyte steatosis in a farnesoid Xreceptor-dependent manner. Toxicol. Appl. Pharmacol. 2017, 315, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Nagaya, T.; Tanaka, N.; Suzuki, T.; Sano, K.; Horiuch, A.; Komatsu, M.; Nakajima, T.; Nishizawa, T.; Joshita, S.; Umemura, T.; et al. Down-regulation of SREBP-1c is associated with the development of burned-out NASH. J. Hepatol. 2010, 53, 724. [Google Scholar] [CrossRef] [PubMed]
- Schindler, M.; Pendzialek, M.; Grybel, K.J.; Seeling, T.; Gürke, J.; Fischer, B.; Santos, A.N. Adiponectin stimulates lipid metabolism via AMPK in rabbit blastocysts. Hum. Reprod. 2017, 32, 1382–1392. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | GenBank | Full Name | Primer (5′-3′) |
|---|---|---|---|
| GAPDH | NM_008084.2 | Glyceraldehyde-3-phosphatedehydrogenase | TGTGTCCGTCGTGGATCTGATTGCTGTTGAAGTCGCAGGAG |
| TNF-α | NM_013693.2 | Tumour necrosis factor alpha | TATGGCCCAGACCCTCACAGGAGTAGACAAGGTACAACCCATC |
| IL-1β | NM_008361.3 | Interleukin-1 beta | TCCAGGATGAGGACATGAGCACGAACGTCACACACCAGCAGGTTA |
| IL-6 | NM_031168.1 | Interleukin-6 | CCACTTCACAAGTCGGAGGCTTACCAGTTTGGTAGCATCCATCATTTC |
| SREBP-1c | NM_011480.3 | Sterol regulatory element-binding proteins-1c | CCGAGATGTGCGAACTGGAGAAGTCACTGTCTTGGTTGTTGATG |
| SCD1 | NM_009127.4 | Stearoyl-Coenzyme desaturase-1 | ATGTCTGACCTGAAAGCCGAGAAGAGCACCAGAGTGTATCGCAAGAA |
| Antibody | Full Name | Source | Dilutions | Company |
|---|---|---|---|---|
| FXR | Farnesoid X Recepter | Rabbit | 1:500 | Bioss, Beijing, China |
| Nrf2 | Nuclear erythroid factor 2-related factor 2 | Rabbit | 1:1000 | Proteintech Group, Chicago, USA |
| Keap1 | Kelch-like ECH-associated protein 1 | Rabbit | 1:1000 | Proteintech Group, Chicago, USA |
| HO-1 | Heme oxygenase-1 | Rabbit | 1:1000 | Proteintech Group, Chicago, USA |
| NQO1 | NAD(P)Hdehydrogenase(quinone 1) | Rabbit | 1:1000 | Proteintech Group, Chicago, USA |
| GCLC | Glutamate-cysteine ligase catalytic subunit | Rabbit | 1:1000 | Proteintech Group, Chicago, USA |
| GCLM | Glutamate-cysteineligase regulatory subunit | Rabbit | 1:1000 | Proteintech Group, Chicago, USA |
| FOXO3a | Forkhead box O3 | Rabbit | 1:1000 | Proteintech Group, Chicago, USA |
| NF-κB | Nuclear factor kappa B | Rabbit | 1:1000 | Proteintech Group, Chicago, USA |
| COX-2 | Cyclooxygenase-2 | Rabbit | 1:1000 | Proteintech Group, Chicago, USA |
| HMGB1 | High-Mobility Group Box 1 | Rabbit | 1:1000 | Proteintech Group, Chicago, USA |
| ACC1 | Acetyl-Coa carboxylase-1 | Rabbit | 1:1000 | Proteintech Group, Chicago, USA |
| FASN | Fatty acid synthase | Rabbit | 1:1000 | Proteintech Group, Chicago, USA |
| CPT1 | Carnitine palmitoyltransferase 1 | Rabbit | 1:1000 | Proteintech Group, Chicago, USA |
| GAPDH | Glyceraldehyde-3-phosphatedehydrogenase | Rabbit | 1:1000 | Proteintech Group, Chicago, USA |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, L.; Han, X.; Tao, X.; Xu, L.; Xu, Y.; Fang, L.; Yin, L.; Qi, Y.; Li, H.; Peng, J. Protection by the Total Flavonoids from Rosa laevigata Michx Fruit against Lipopolysaccharide-Induced Liver Injury in Mice via Modulation of FXR Signaling. Foods 2018, 7, 88. https://doi.org/10.3390/foods7060088
Dong L, Han X, Tao X, Xu L, Xu Y, Fang L, Yin L, Qi Y, Li H, Peng J. Protection by the Total Flavonoids from Rosa laevigata Michx Fruit against Lipopolysaccharide-Induced Liver Injury in Mice via Modulation of FXR Signaling. Foods. 2018; 7(6):88. https://doi.org/10.3390/foods7060088
Chicago/Turabian StyleDong, Lile, Xu Han, Xufeng Tao, Lina Xu, Youwei Xu, Linlin Fang, Lianhong Yin, Yan Qi, Hua Li, and Jinyong Peng. 2018. "Protection by the Total Flavonoids from Rosa laevigata Michx Fruit against Lipopolysaccharide-Induced Liver Injury in Mice via Modulation of FXR Signaling" Foods 7, no. 6: 88. https://doi.org/10.3390/foods7060088
APA StyleDong, L., Han, X., Tao, X., Xu, L., Xu, Y., Fang, L., Yin, L., Qi, Y., Li, H., & Peng, J. (2018). Protection by the Total Flavonoids from Rosa laevigata Michx Fruit against Lipopolysaccharide-Induced Liver Injury in Mice via Modulation of FXR Signaling. Foods, 7(6), 88. https://doi.org/10.3390/foods7060088