Analysis of the Microbial Diversity and Characteristics of Fermented Blueberry Beverages from Different Regions



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction, 16S rRNA, and Internal Transcribed Spacer (ITS) Sequencing
2.3. Enumeration of Lactic Acid Bacteria and Yeast
2.4. Determination of pH Value, Total Soluble Solids and Titratable Acidity
2.5. Determination of Total Phenolic Content
2.6. Determination of Total Flavonoid Content
2.7. Determination of Total Anthocyanin Content
2.8. Determination of Antioxidant Capacity
2.9. Determination of Superoxide Dismutase
2.10. Data Analysis
3. Results
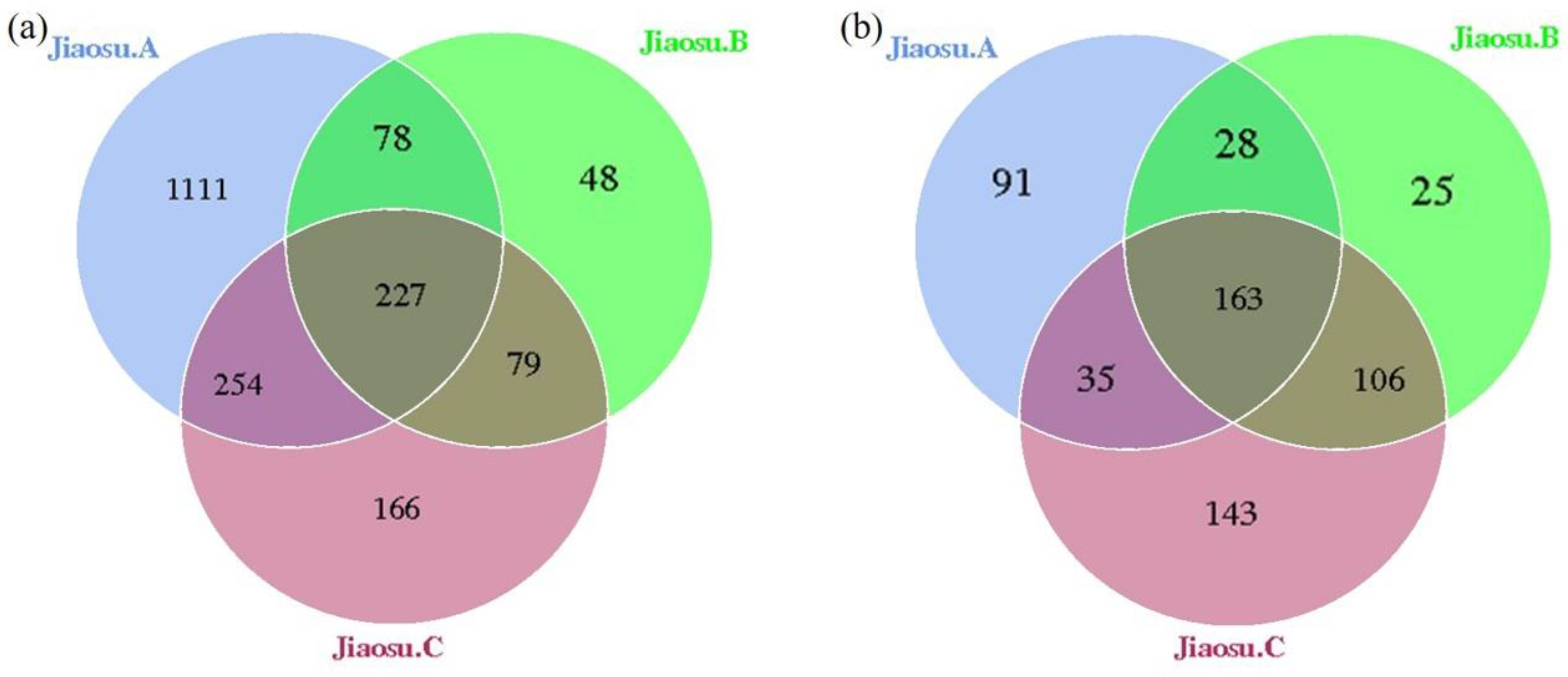
3.1. Abundance and Diversity of Bacteria and Fungi in Traditional Fermented Blueberry Beverages
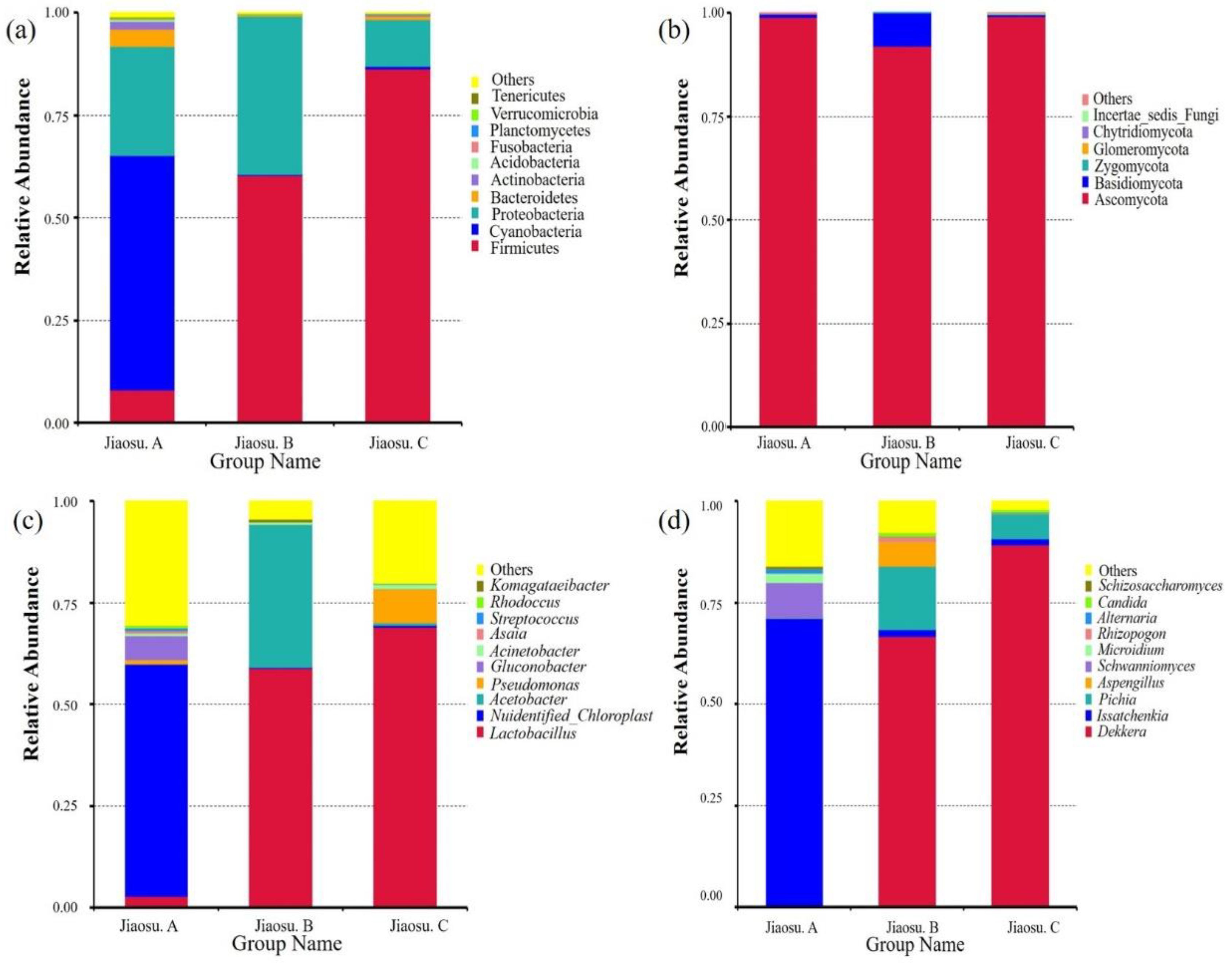
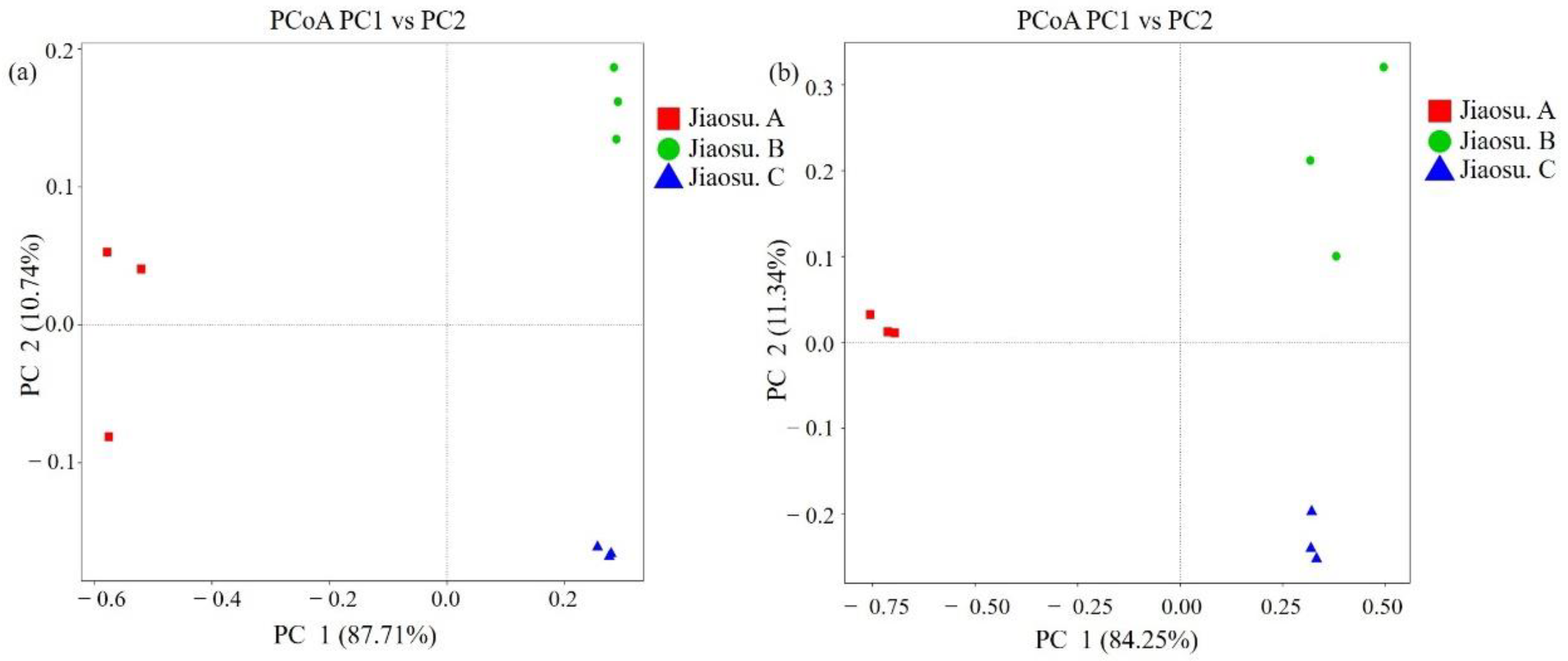
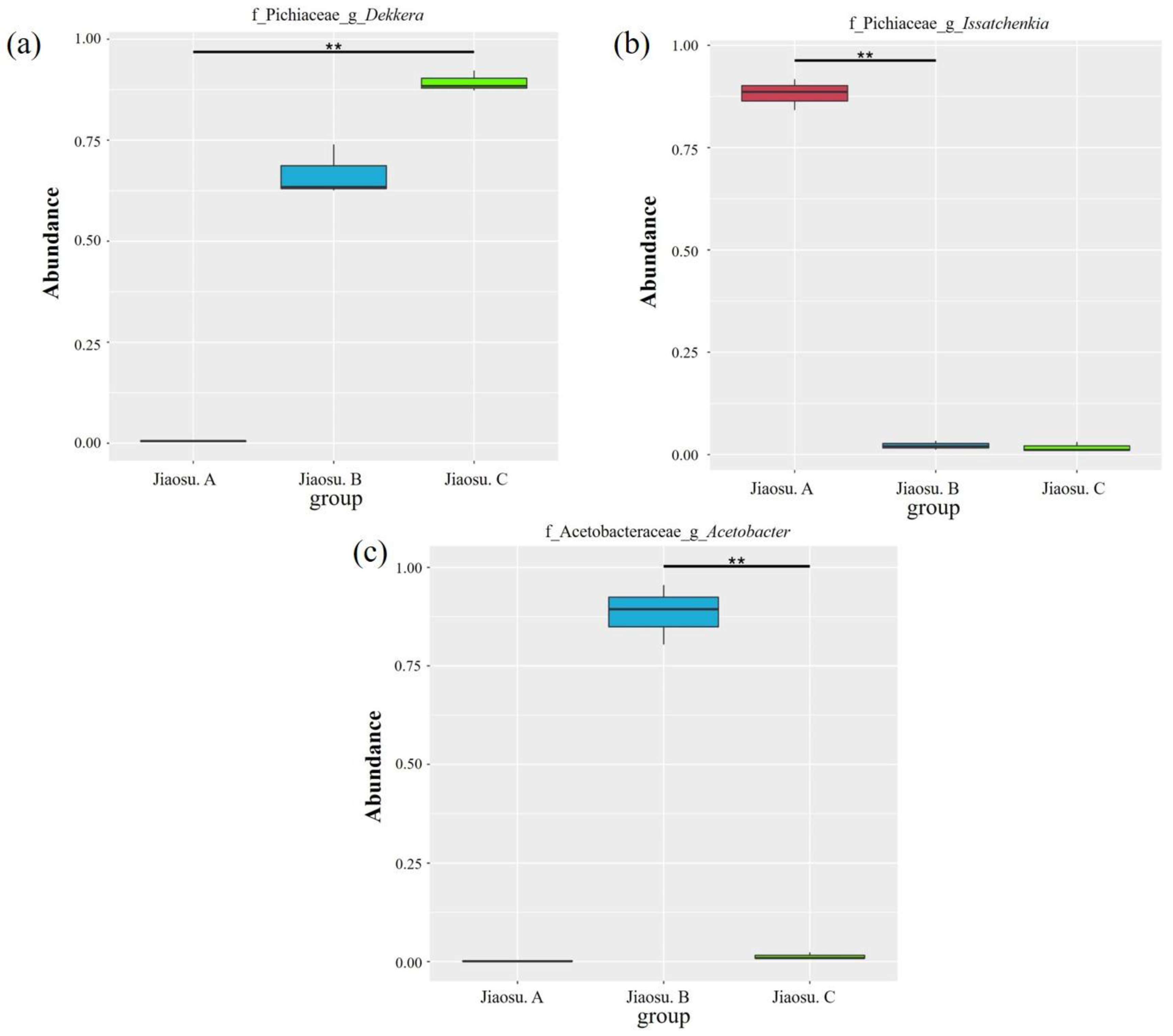
3.2. Bacterial and Fungal Community Distribution in Traditional Fermented Blueberry Beverage Samples
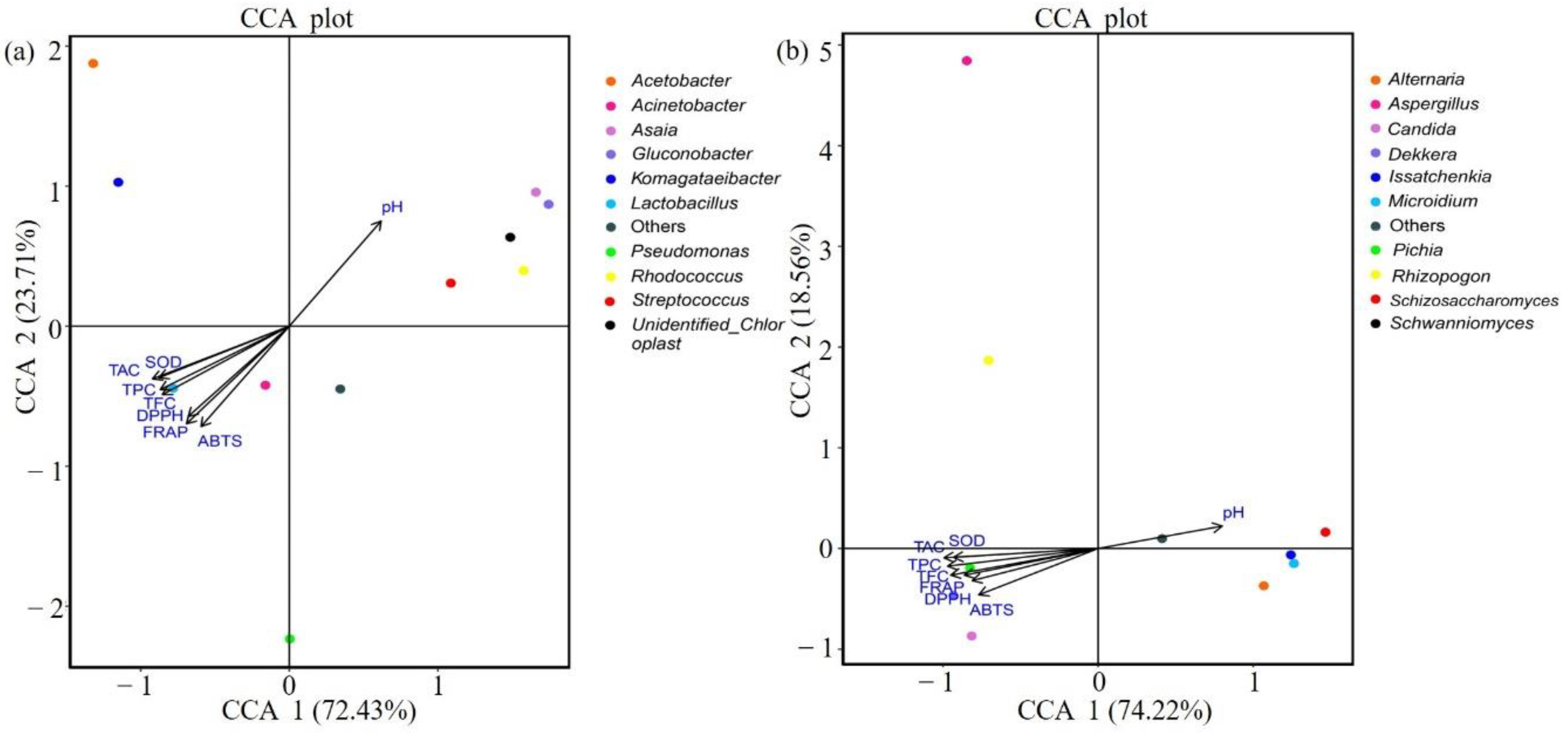
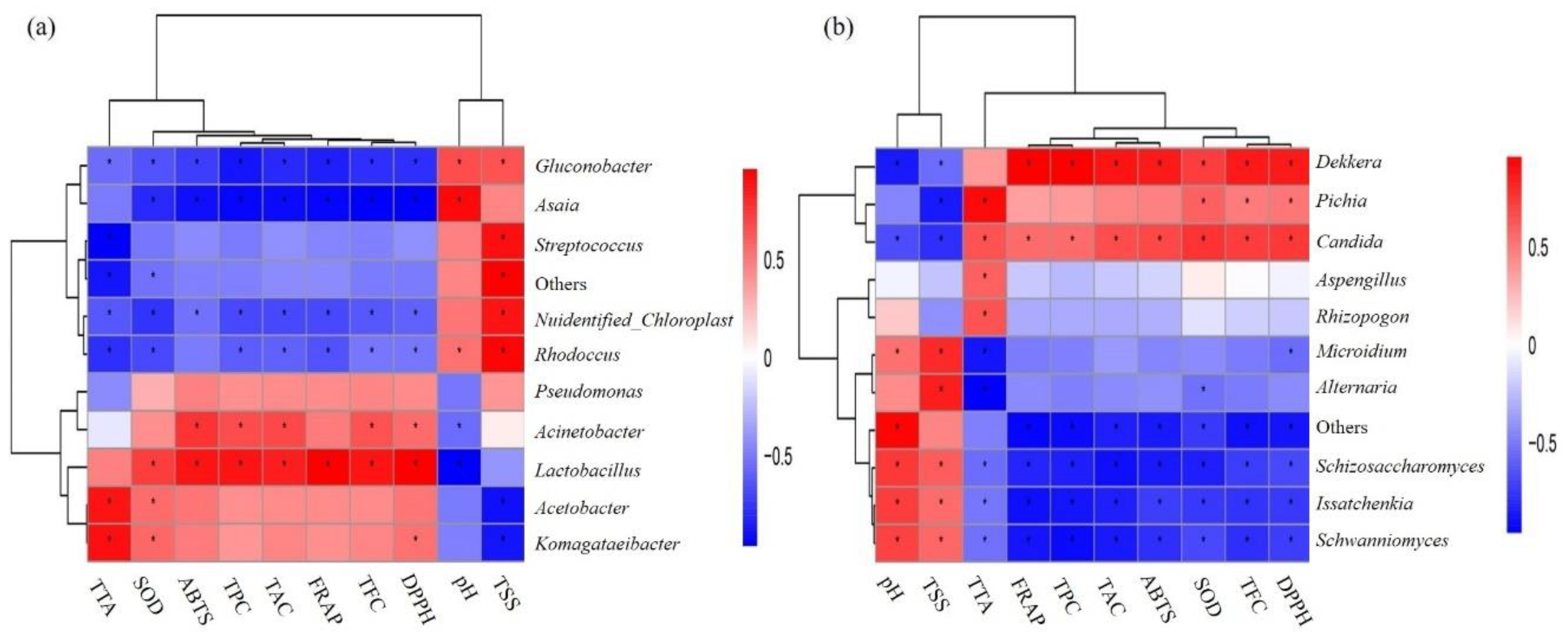
3.3. Comparison of Bacterial and Fungal Communities in Traditional Fermented Blueberry Beverage Samples
3.4. LAB, Yeast Count, and Antioxidant Indicators
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Holzapfel, W.H. Appropriate starter culture technologies for small-scale fermentation in developing countries. Int. J. Food Microbiol. 2002, 75, 197–212. [Google Scholar] [CrossRef]
- Adebo, O.A.; Kayitesi, E.; Tugizimana, F.; Njobeh, P.B. Differential metabolic signatures in naturally and lactic acid bacteria (LAB) fermented ting (a Southern African food) with different tannin content, as revealed by gas chromatography mass spectrometry (GC–MS)-based metabolomics. Food Res. Int. 2019, 121, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Sumby, K.M.; Bartle, L.; Grbin, P.R.; Jiranek, V. Measures to improve wine malolactic fermentation. Appl. Microbiol. Biot. 2019, 103, 2033–2051. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Xiong, T.; Peng, Z.; Liu, C.; Huang, T.; Yu, H.; Xie, M. Correlation between microbiota and flavours in fermentation of Chinese Sichuan Paocai. Food Res. Int. 2018, 114, 123–132. [Google Scholar] [CrossRef]
- Pico, J.; Antolín, B.; Román, L.; Gómez, M.; Bernal, J. Analysis of volatile compounds in gluten-free bread crusts with an optimised and validated SPME-GC/QTOF methodology. Food Res. Int. 2018, 106, 686–695. [Google Scholar] [CrossRef]
- Hu, N.; Lei, M.; Zhao, X.; Wang, Y.; Zhang, Y.; Wang, S. Analysis of microbiota in Hainan Yucha during fermentation by 16S rRNA gene high-throughput sequencing. J. Food Process. Pres. 2020, 44. [Google Scholar] [CrossRef]
- McGovern, P.E.; Zhang, J.; Tang, J.; Zhang, Z.; Hall, G.R.; Moreau, R.A.; Nuñez, A.; Butrym, E.D.; Richards, M.P.; Wang, C.S. Fermented beverages of pre-and proto-historic China. Proc. Natl. Acad. Sci. USA 2004, 101, 17593–17598. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.; Yan, H.; Han, C.; Wang, W.; Tian, Y.; Chen, X. Polyphenols from blueberries modulate inflammation cytokines in LPS-induced RAW264. 7 macrophages. Int. J. Biol. Macromol. 2014, 69, 382–387. [Google Scholar] [CrossRef]
- Kraujalytė, V.; Venskutonis, P.R.; Pukalskas, A.; Česonienė, L.; Daubaras, R. Antioxidant properties, phenolic composition and potentiometric sensor array evaluation of commercial and new blueberry (Vaccinium corymbosum) and bog blueberry (Vaccinium uliginosum) genotypes. Food Chem. 2015, 188, 583–590. [Google Scholar] [CrossRef]
- Ramos, C.L.; de Almeida, E.G.; de Melo Pereira, G.V.; Cardoso, P.G.; Dias, E.S.; Schwan, R.F. Determination of dynamic characteristics of microbiota in a fermented beverage produced by Brazilian Amerindians using culture-dependent and culture-independent methods. Int. J. Food Microbiol. 2010, 140, 225–231. [Google Scholar] [CrossRef]
- Blandino, A.; Al-Aseeri, M.E.; Pandiella, S.S.; Cantero, D.; Webb, C. Cereal-based fermented foods and beverages. Food Res. Int. 2003, 36, 527–543. [Google Scholar] [CrossRef]
- Azeke, M.A.; Fretzdorff, B.; Buening-Pfaue, H.; Holzapfel, W.; Betsche, T. Nutritional value of African yambean (Sphenostylis stenocarpa L): Improvement by lactic acid fermentation. J. Sci. Food Agric. 2005, 85, 963–970. [Google Scholar] [CrossRef]
- Lee, B.J.; Kim, J.S.; Kang, Y.M.; Lim, J.H.; Kim, Y.M.; Lee, M.S.; Jeong, M.H.; Ahn, C.B.; Je, J.Y. Antioxidant activity and γ-aminobutyric acid (GABA) content in sea tangle fermented by Lactobacillus brevis BJ20 isolated from traditional fermented foods. Food Chem. 2010, 122, 271–276. [Google Scholar] [CrossRef]
- Chae, G.Y.; Ha, B.J. The Comparative Evaluation of Fermented and Non-fermented Soybean Extract on Antioxidation and Whitening. Toxicol. Res. 2011, 27, 205–209. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhao, Y.; Zhang, L.; Zhang, X.; Huang, L.; Li, D.; Niu, C.; Yang, Z.; Wang, Q. Antioxidant activity of Lactobacillus plantarum strains isolated from traditional Chinese fermented foods. Food Chem. 2012, 135, 1914–1919. [Google Scholar] [CrossRef]
- Li, J.Y.; Jin, M.M.; Meng, J.; Gao, S.M.; Lu, R.R. Exopolysaccharide from Lactobacillus planterum LP6: Antioxidation and the effect on oxidative stress. Carbohydr. Polym. 2013, 98, 1147–1152. [Google Scholar] [CrossRef]
- Tsai, C.C.; Chan, C.F.; Huang, W.Y.; Lin, J.S.; Chan, P.; Liu, H.Y.; Lin, Y.S. Applications of Lactobacillus rhamnosus spent culture supernatant in cosmetic antioxidation, whitening and moisture retention applications. Molecules 2013, 18, 14161–14171. [Google Scholar] [CrossRef]
- Landete, J.M.; Curiel, J.A.; Rodríguez, H.; De Las Rivas, B.; Muñoz, R. Aryl glycosidases from Lactobacillus plantarum increase antioxidant activity of phenolic compounds. J. Funct. Foods 2014, 7, 322–329. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.Y.; Li, J.S.; Chen, C. Effects of blending wheatgrass juice on enhancing phenolic compounds and antioxidant activities of traditional kombucha beverage. J. Food. Drug Anal. 2015, 23, 709–718. [Google Scholar] [CrossRef] [Green Version]
- Xiong, T.; Peng, F.; Liu, Y.; Deng, Y.; Wang, X.; Xie, M. Fermentation of Chinese sauerkraut in pure culture and binary co-culture with Leuconostoc mesenteroides and Lactobacillus plantarum. LWT 2014, 59, 713–717. [Google Scholar] [CrossRef]
- Wu, R.; Yu, M.; Liu, X.; Meng, L.; Wang, Q.; Xue, Y.; Wu, J.; Yue, X. Changes in flavour and microbial diversity during natural fermentation of suan-cai, a traditional food made in Northeast China. Int. J. Food Microbiol. 2015, 211, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Van Hijum, S.A.F.T.; Vaughan, E.E.; Vogel, R.F. Application of state-of-art sequencing technologies to indigenous food fermentations. Curr. Opin. Biotech. 2013, 24, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, L.M.; Neef, A.; Vignolo, G.; Belloch, C. Yeast diversity during the fermentation of Andean chicha: A comparison of high-throughput sequencing and culture-dependent approaches. Food Microbiol. 2017, 67, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Maguire, T.L.; Collins, G.G.; Sedgley, M. A modified CTAB DNA extraction procedure for plants belonging to the family Proteaceae. Plant. Mol. Biol. Rep. 1994, 12, 106–109. [Google Scholar] [CrossRef]
- Kocabaş, M.; Kutluyer, F.; Ertekin, Ö.; Aksu, Ö.; Başçınar, N. Improvement of sperm motility of Oncorhynchus mykiss and Salvelinus fontinalis by L-tryptophan. Syst. Biol. Reprod. Med. 2019, 2, 1–7. [Google Scholar] [CrossRef]
- Berenguer, M.; Vegara, S.; Barrajón, E.; Saura, D.; Valero, M.; Martí, N. Physicochemical characterization of pomegranate wines fermented with three different Saccharomyces cerevisiae yeast strains. Food Chem. 2016, 190, 848–855. [Google Scholar] [CrossRef]
- Silva, V.L.M.; Silva, A.C.O.; Costa-Lima, B.R.C.; Carneiro, C.S.; Conte-Junior, C.A. Stability of polyphenols from blueberry (Vaccinium corymbosum L.) in fermented dairy beverage. J. Food Process. Preserv. 2017, 41, e13305. [Google Scholar] [CrossRef]
- Sun, Y.; Li, M.; Mitra, S.; Muhammad, R.H.; Debnath, B.; Lu, X.; Jian, H.; Qiu, D. Comparative phytochemical profiles and antioxidant enzyme activity analyses of the southern highbush blueberry (Vaccinium corymbosum) at different developmental stages. Molecules 2018, 23, 2209. [Google Scholar] [CrossRef] [Green Version]
- Hwang, H.; Kim, Y.J.; Shin, Y. Assessment of physicochemical quality, antioxidant content and activity, and inhibition of cholinesterase between unripe and ripe blueberry fruit. Foods 2020, 9, 690. [Google Scholar] [CrossRef]
- De Souza, A.C.; Fernandes, A.C.F.; Silva, M.S.; Schwan, R.F.; Dias, D.R. Antioxidant activities of tropical fruit wines. J. Inst. Brew. 2018, 124, 492–497. [Google Scholar] [CrossRef] [Green Version]
- Oszmiański, J.; Wojdylo, A. Aronia melanocarpa phenolics and their antioxidant activity. Eur. Food Res. Technol. 2005, 221, 809–813. [Google Scholar] [CrossRef]
- Yang, J.; Liu, X.; Zhang, X.; Jin, Q.; Li, J. Phenolic profiles, antioxidant activities, and neuroprotective properties of mulberry (Morus atropurpurea Roxb.) fruit extracts from different ripening stages. J. Food Sci. 2016, 81, C2439–C2446. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Stiles, M.E.; Holzapfel, W.H. Lactic acid bacteria of foods and their current taxonomy. Int. J. Food Microbiol. 1997, 36, 1–29. [Google Scholar] [CrossRef]
- Tamang, J.P.; Watanabe, K.; Holzapfel, W.H. Review: Diversity of microorganisms in global fermented foods and beverages. Front. Microbiol. 2016, 7, 377. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Tong, C. Bacterial community diversity of traditional fermented vegetables in China. LWT 2017, 86, 40–48. [Google Scholar] [CrossRef]
- Kwon, Y.J.; Wang, F.; Liu, C.Z. Deep-bed solid state fermentation of sweet sorghum stalk to ethanol by thermotolerant Issatchenkia orientalis IPE 100. Bioresour. Technol. 2011, 102, 11262–11265. [Google Scholar] [CrossRef]
- Negi, B.; Dey, G. Effects of co-fermentation by Saccharomyces cerevisiae and Issatchenkia orientalis on sea buckthorn juice. Int. J. Food Sci. Nutr. 2013, 64, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Hong, Y.A.; Park, H.D. Co-fermentation of grape must by Issatchenkia orientalis and Saccharomyces cerevisiae reduces the malic acid content in wine. Biotechnol. Lett. 2008, 30, 1633–1638. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.W.; Wen, P.F.; Wu, Z.F. Fermentation efficiency and flavor characteristic of mixed fermentation of bayberry wine by Issatchenkio orientalis and Saccharomyces cerevisiae. Food Ence. 2019, 40, 144–151. [Google Scholar] [CrossRef]
- Tamang, J.P. Diversity of fermented foods. In Fermented Foods and Beverages of the World; Tamang, J.P., Kailasapathy, K., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 41–84. [Google Scholar] [CrossRef]
- Schifferdecker, A.J.; Dashko, S.; Ishchuk, O.P.; Piškur, J. The wine and beer yeast Dekkera bruxellensis. Yeast 2014, 31, 323–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, P.; Capece, A.; Jespersen, L. Taxonomic and ecological diversity of food and beverage yeasts. In Yeasts in Food and Beverages; Querol, A., Fleet, G.H., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 13–53. [Google Scholar] [CrossRef]
- Tamang, J.P.; Fleet, G.H. Yeasts diversity in fermented foods and beverages. In Yeast Biotechnology: Diversity and Applications; Kunze Satyanarayana, T., Kunze, G., Eds.; Springer: New York, NY, USA, 2009; pp. 169–198. [Google Scholar] [CrossRef]
- Lv, X.C.; Huang, X.L.; Zhang, W.; Rao, P.F.; Ni, L. Yeast diversity of traditional alcohol fermentation starters for Hong Qu glutinous rice wine brewing, revealed by culture-dependent and culture-independent methods. Food Control 2013, 34, 183–190. [Google Scholar] [CrossRef]
- Burattini, E.; Cavagna, M.; Dell’Anna, R.; Campeggi, F.M.; Monti, F.; Rossi, F.; Torriani, S. A FTIR microspectroscopy study of autolysis in cells of the wine yeast Saccharomyces cerevisiae. Vib. Spectrosc. 2008, 47, 139–147. [Google Scholar] [CrossRef]
- Vong, W.C.; Liu, S.Q. Changes in volatile profile of soybean residue (okara) upon solid-state fermentation by yeasts. J. Sci. Food Agric. 2017, 97, 135–143. [Google Scholar] [CrossRef]
- Dalié, D.K.D.; Deschamps, A.M.; Richard-Forget, F. Lactic acid bacteria–Potential for control of mould growth and mycotoxins: A review. Food Control 2010, 21, 370–380. [Google Scholar] [CrossRef]
- Li, D.; Li, B.; Ma, Y.; Sun, X.; Lin, Y.; Meng, X. Polyphenols, anthocyanins, and flavonoids contents and the antioxidant capacity of various cultivars of highbush and half-high blueberries. J. Food Compos. Anal. 2017, 62, 84–93. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Bio. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Ogawa, K.; Sakakibara, H.; Iwata, R.; Ishii, T.; Sato, T.; Goda, T.; Shimoi, K.; Kumazawa, S. Anthocyanin composition and antioxidant activity of the crowberry (Empetrum nigrum) and other berries. J. Agric. Food Chem. 2008, 56, 4457–4462. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Tran, G.B.; Nguyen, C.T. Anti-oxidative effects of superoxide dismutase 3 on inflammatory diseases. J. Mol. Med. 2020, 98, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Oh, B.T.; Jeong, S.Y.; Velmurugan, P.; Park, J.H.; Jeong, D.Y. Probiotic-mediated blueberry (Vaccinium corymbosum L.) fruit fermentation to yield functionalized products for augmented antibacterial and antioxidant activity. J. Biosci. Bioeng. 2017, 124, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, T.; He, G. Antioxidant Activity of Peptides from Fermented Milk with Mix Culture of Lactic Acid Bacteria and Yeast. Adv. J. Food Sci. Technol. 2015, 7, 422–427. [Google Scholar] [CrossRef]
- Wang, L.; Luo, Y.; Wu, Y.; Liu, Y.; Wu, Z. Fermentation and complex enzyme hydrolysis for improving the total soluble phenolic contents, flavonoid aglycones contents and bio-activities of guava leaves tea. Food Chem. 2018, 264, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Hur, S.J.; Lee, S.Y.; Kim, Y.C.; Choi, I.; Kim, G.B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, C.; Li, D.; Zhao, Y.; Zhang, X.; Zeng, X.; Yang, Z.; Li, S. Antioxidant activity of an exopolysaccharide isolated from Lactobacillus plantarum C88. Int. J. Biol. Macromol. 2013, 54, 270–275. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Raw Reads | Clean Tags | Number of OTUs | Observed Species | Chao1 Index | Simpson Index | Shannon Index | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bacteria | Fungi | Bacteria | Fungi | Bacteria | Fungi | Bacteria | Fungi | Bacteria | Fungi | Bacteria | Fungi | Bacteria | Fungi | |
| Jiaosu A | 238,455 | 221,394 | 213,279 | 183,055 | 1670 | 317 | 888 | 209 | 1005.125 | 232.498 | 0.868 | 0.928 | 4.907 | 4.737 |
| Jiaosu B | 251,629 | 199,545 | 235,353 | 168,265 | 432 | 322 | 287 | 248 | 329.996 | 277.954 | 0.775 | 0.914 | 3.477 | 5.054 |
| Jiaosu C | 240,571 | 204,011 | 220,616 | 175,779 | 726 | 447 | 408 | 286 | 438.773 | 323.8 | 0.883 | 0.668 | 4.024 | 3.554 |
| Samples | Jiaosu A | Jiaosu B | Jiaosu C |
|---|---|---|---|
| LAB enumeration (log CFU/mL) | 4.31 ± 0.23 a | 9.86 ± 0.10 b | 10.90 ± 0.05 c |
| Yeast enumeration (log CFU/mL) | 7.35 ± 0.22 c | 6.71 ± 0.13 a | 6.95 ± 0.08 b |
| TTA (ACA, mg/mL) | 6.61 ± 0.10 a | 7.36 ± 0.10 c | 7.00 ± 0.11 b |
| TPC (GAE, mg/mL) | 3.23 ± 0.15 a | 5.05 ± 0.25 b | 5.85 ± 0.21 c |
| TFC (RE, mg/mL) | 0.44 ± 0.01 a | 0.60 ± 0.03 b | 0.68 ± 0.02 c |
| TAC (C3GE, mg/mL) | 0.32 ± 0.003 a | 0.55 ± 0.002 b | 0.60 ± 0.004 c |
| ABTS (IC50, mg/mL) | 2.40 ± 0.02 a | 2.45 ± 0.02 b | 2.54 ± 0.02 c |
| DPPH (IC50, mg/mL) | 1.73 ± 0.08 a | 2.50 ± 0.39 b | 2.53 ± 0.17 b |
| FRAP (U/mL) | 13.14 ± 0.28 a | 15.85 ± 0.55 b | 19.55 ± 0.16 c |
| SOD (U/mL) | 162.72 ± 22.90 a | 263.55 ± 19.61 b | 285.27 ± 4.91 b |
| TSS (°Bx) | 3.5 ± 0.23 b | 2.4 ± 0.06 a | 2.6 ± 0.06 a |
| pH | 3.92 ± 0.03 c | 3.55 ± 0.04 a | 3.77 ± 0.05 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, N.; Lei, M.; Zhao, X.; Zhang, Z.; Gu, Y.; Zhang, Y.; Wang, S. Analysis of the Microbial Diversity and Characteristics of Fermented Blueberry Beverages from Different Regions. Foods 2020, 9, 1656. https://doi.org/10.3390/foods9111656
Hu N, Lei M, Zhao X, Zhang Z, Gu Y, Zhang Y, Wang S. Analysis of the Microbial Diversity and Characteristics of Fermented Blueberry Beverages from Different Regions. Foods. 2020; 9(11):1656. https://doi.org/10.3390/foods9111656
Chicago/Turabian StyleHu, Nan, Ming Lei, Xiuli Zhao, Zhen Zhang, Ying Gu, Yan Zhang, and Shuo Wang. 2020. "Analysis of the Microbial Diversity and Characteristics of Fermented Blueberry Beverages from Different Regions" Foods 9, no. 11: 1656. https://doi.org/10.3390/foods9111656
APA StyleHu, N., Lei, M., Zhao, X., Zhang, Z., Gu, Y., Zhang, Y., & Wang, S. (2020). Analysis of the Microbial Diversity and Characteristics of Fermented Blueberry Beverages from Different Regions. Foods, 9(11), 1656. https://doi.org/10.3390/foods9111656