Korean Red Ginseng Suppresses the Expression of Oxidative Stress Response and NLRP3 Inflammasome Genes in Aged C57BL/6 Mouse Ovaries

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
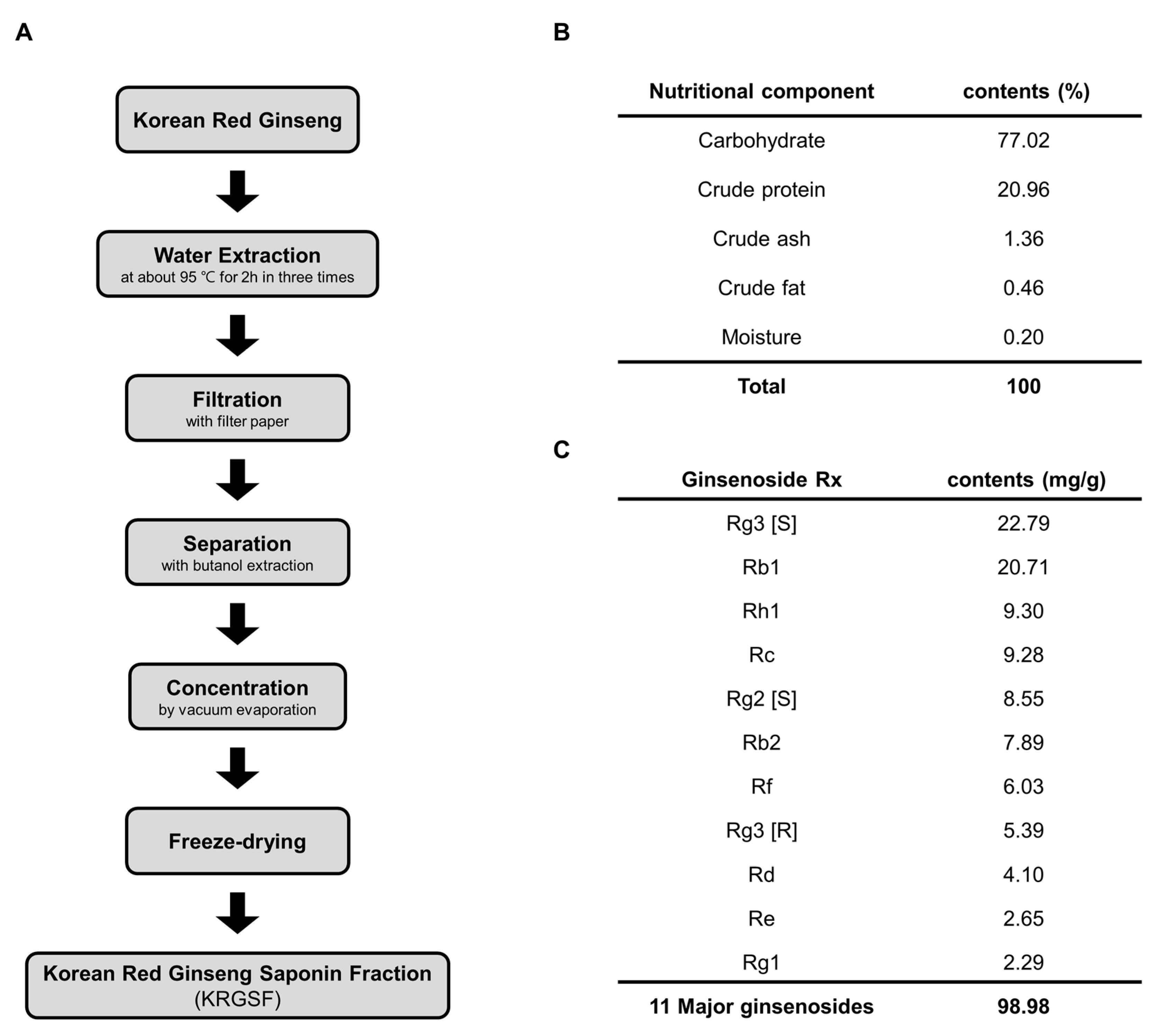
2.1. Preparation of Ginseng Saponin Fraction
2.2. Chemicals and Reagents
2.3. Animal Experiments
2.4. Culture and KRGSF Treatment of Mouse Ovaries
2.5. Total RNA Isolation and Microarray Analysis
2.6. qRT-PCR
2.7. Statistical Analysis
3. Results
3.1. Preparation and Composition of KRGSF
3.2. Age and KRGSF Treatment Affect the Ovarian mRNA Expression Profile
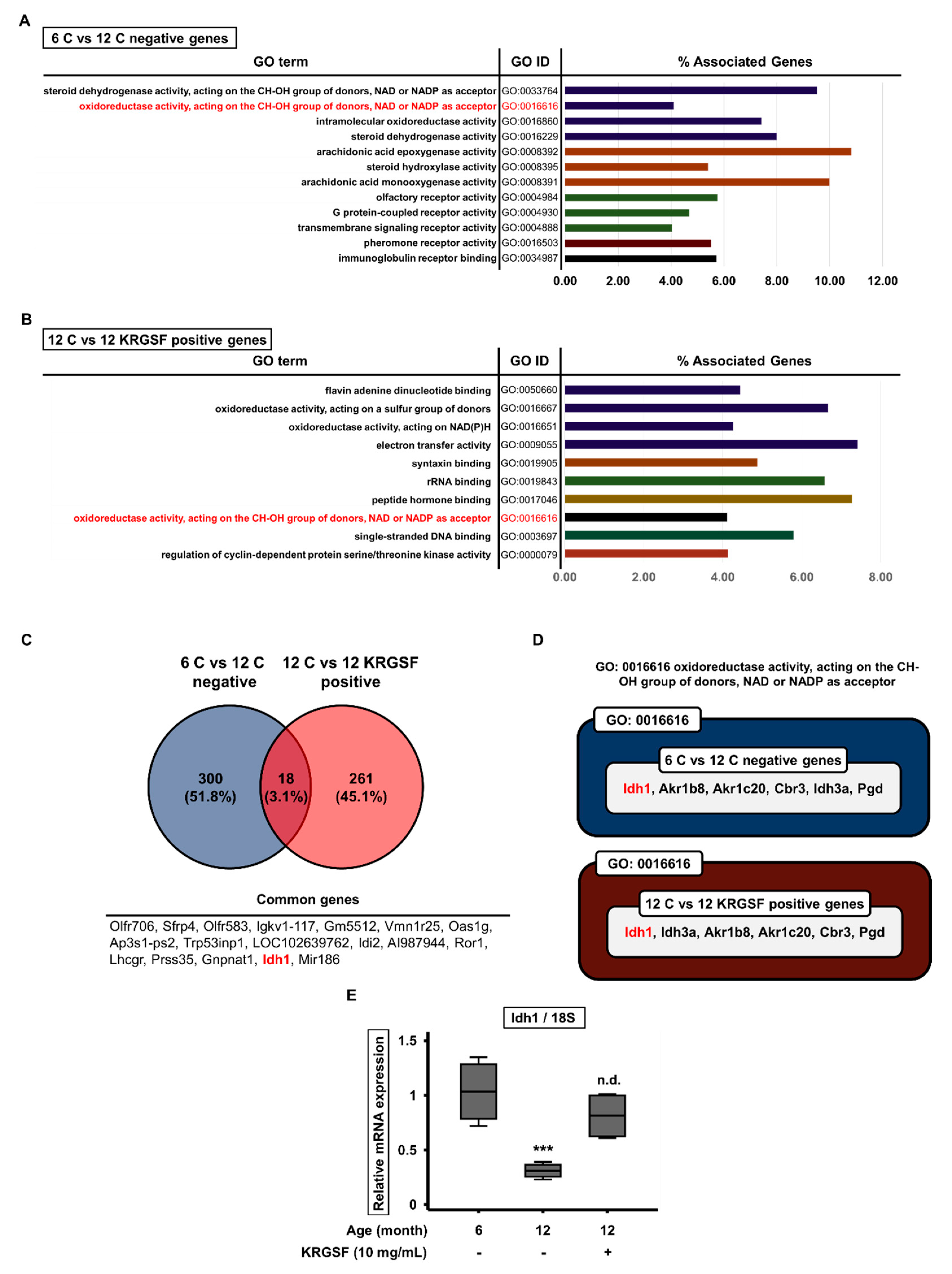
3.3. Gene Ontology Enrichment Analysis to Identify the Functions of Genes Affected by KRGSF Treatment
3.4. KRGSF Inhibits Age-Related Intracellular Oxidative Stress Response
3.5. KRGSF Ameliorates the Age-Related Upregulation of NLRP3 Inflammasome Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cervellati, M.; Sunde, U. Life expectancy and economic growth: The role of the demographic transition. J. Econ. Growth 2011, 16, 99–133. [Google Scholar] [CrossRef]
- Mishra, S. Does modern medicine increase life-expectancy: Quest for the Moon Rabbit? Indian Hear. J. 2016, 68, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Oeppen, J.; Vaupel, J.W. DEMOGRAPHY: Enhanced: Broken Limits to Life Expectancy. Science 2002, 296, 1029–1031. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.W. Delayed Marriage and Very Low Fertility in Pacific Asia. Popul. Dev. Rev. 2007, 33, 453–478. [Google Scholar] [CrossRef]
- Ames, B.N.; Shigenaga, M.K.; Hagen, T.M. Oxidants, antioxidants, and the degenerative diseases of aging. Proc. Natl. Acad. Sci. USA 1993, 90, 7915–7922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csiszar, A.; Ungvari, Z.; Koller, A.; Edwards, J.; Kaley, G. Aging-induced proinflammatory shift in cytokine expression profile in rat coronary arteries. FASEB J. 2003, 17, 1183–1185. [Google Scholar] [CrossRef] [PubMed]
- Carnevale, E.; Ginther, O. Defective Oocytes as a Cause of Subfertility in Old Mares1. Boil. Reprod. 1995, 52, 209–214. [Google Scholar] [CrossRef]
- Huang, Y.; Hu, C.; Ye, H.; Luo, R.; Fu, X.; Li, X.; Huang, J.; Chen, W.; Zheng, Y.-H. Inflamm-Aging: A New Mechanism Affecting Premature Ovarian Insufficiency. J. Immunol. Res. 2019, 2019, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Solon-Biet, S.M.; Walters, K.; Simanainen, U.K.; McMahon, A.C.; Ruohonen, K.; Ballard, J.W.O.; Raubenheimer, D.; Handelsman, D.; Le Couteur, D.G.; Simpson, S.J. Macronutrient balance, reproductive function, and lifespan in aging mice. Proc. Natl. Acad. Sci. USA 2015, 112, 3481–3486. [Google Scholar] [CrossRef] [Green Version]
- Broekmans, F.J.; Soules, M.R.; Fauser, B.C. Ovarian Aging: Mechanisms and Clinical Consequences. Endocr. Rev. 2009, 30, 465–493. [Google Scholar] [CrossRef] [Green Version]
- Itsumi, M.; Inoue, S.; Elia, A.J.; Murakami, K.; Sasaki, M.; Lind, E.F.; Brenner, D.; Harris, I.; Chio, I.I.C.; Afzal, S.; et al. Idh1 protects murine hepatocytes from endotoxin-induced oxidative stress by regulating the intracellular NADP+/NADPH ratio. Cell Death Differ. 2015, 22, 1837–1845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chei, S.; Oh, H.-J.; Song, J.-H.; Seo, Y.-J.; Lee, K.; Kim, K.-J.; Lee, B.-Y. Spirulina maxima extract prevents activation of the NLRP3 inflammasome by inhibiting ERK signaling. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Chei, S.; Song, J.-H.; Oh, H.-J.; Lee, K.; Jin, H.; Choi, S.-H.; Nah, S.-Y.; Lee, B.-Y. Gintonin-Enriched Fraction Suppresses Heat Stress-Induced Inflammation through LPA Receptor. Molecules 2020, 25, 1019. [Google Scholar] [CrossRef] [Green Version]
- Song, J.-H.; Kim, K.-J.; Choi, S.-Y.; Koh, E.-J.; Park, J.; Lee, B.-Y. Korean ginseng extract ameliorates abnormal immune response through the regulation of inflammatory constituents in Sprague Dawley rat subjected to environmental heat stress. J. Ginseng Res. 2019, 43, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Seo, Y.-J.; Song, J.-H.; Chei, S.; Lee, B.-Y. Ginsenoside Rg1 promotes browning by inducing UCP1 expression and mitochondrial activity in 3T3-L1 and subcutaneous white adipocytes. J. Ginseng Res. 2018, 43, 589–599. [Google Scholar] [CrossRef]
- Lee, C.H.; Kim, J.-H. A review on the medicinal potentials of ginseng and ginsenosides on cardiovascular diseases. J. Ginseng Res. 2014, 38, 161–166. [Google Scholar] [CrossRef] [Green Version]
- Lim, W.; Mudge, K.W.; Vermeylen, F. Effects of Population, Age, and Cultivation Methods on Ginsenoside Content of Wild American Ginseng (Panax quinquefolium). J. Agric. Food Chem. 2005, 53, 8498–8505. [Google Scholar] [CrossRef]
- Lee, G.S.; Nam, K.Y.; Choi, J.E. Ginsenoside Composition and Quality Characteristics of Red Ginseng Extracts prepared with Different Extracting Methods. Korean J. Med. Crop. Sci. 2013, 21, 276–281. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.-Y.; Choi, E.-H.; Wee, J.-J. The difference of ginsenoside compositions according to the conditions of extraction and fractionation of crude ginseng saponins. Korean J. Food Sci. Technol. 2001, 33, 282–287. [Google Scholar]
- Xie, J.-T.; Mehendale, S.; Yuan, C.-S. Ginseng and Diabetes. Am. J. Chin. Med. 2005, 33, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Salvati, G.; Genovesi, G.; Marcellini, L.; Paolini, P.; De Nuccio, I.; Pepe, M.; Re, M. Effects of Panax Ginseng C.A. Meyer saponins on male fertility. Panminerva Medica 1996, 38, 249–254. [Google Scholar] [PubMed]
- Hwang, S.Y.; Kim, W.J.; Wee, J.J.; Choi, J.S.; Kim, S.K. Panax ginseng improves survival and sperm quality in guinea pigs exposed to 2, 3, 7, 8-tetrachlorodibenzo-p-dioxin. BJU Int. 2004, 94, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Morgante, G.; Scolaro, V.; Tosti, C.; Di Sabatino, A.; Piomboni, P.; De Leo, V. [Treatment with carnitine, acetyl carnitine, L-arginine and ginseng improves sperm motility and sexual health in men with asthenopermia]. Minerva Urol. Nefrol. 2010, 62, 213–218. [Google Scholar] [PubMed]
- Yayeh, T.; Jung, K.-H.; Jeong, H.-Y.; Park, J.-H.; Song, Y.-B.; Kwak, Y.-S.; Kang, H.-S.; Cho, J.-Y.; Oh, J.-W.; Kim, S.-K.; et al. Korean Red Ginseng Saponin Fraction Downregulates Proinflammatory Mediators in LPS Stimulated RAW264.7 Cells and Protects Mice against Endotoxic Shock. J. Ginseng Res. 2012, 36, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.-H.; Park, K.-Y.; Jeon, J.-H.; Kwak, Y.-S.; Song, Y.-B.; Wee, J.-J.; Rhee, M.-H.; Kim, T.W. Red Ginseng Saponin Fraction A Isolated from Korean Red Ginseng by Ultrafiltration on the Porcine Coronary Artery. J. Ginseng Res. 2011, 35, 325–330. [Google Scholar] [CrossRef] [Green Version]
- Jang, H.; Na, Y.; Hong, K.; Lee, S.; Moon, S.; Cho, M.; Park, M.; Lee, O.-H.; Chang, E.M.; Lee, N.R.; et al. Synergistic effect of melatonin and ghrelin in preventing cisplatin-induced ovarian damage via regulation of FOXO3a phosphorylation and binding to thep27Kip1promoter in primordial follicles. J. Pineal Res. 2017, 63, e12432. [Google Scholar] [CrossRef]
- Minutoli, L.; Puzzolo, D.; Rinaldi, M.; Irrera, N.; Marini, H.R.; Arcoraci, V.; Bitto, A.; Crea, G.; Pisani, A.; Squadrito, F.; et al. ROS-Mediated NLRP3 Inflammasome Activation in Brain, Heart, Kidney, and Testis Ischemia/Reperfusion Injury. Oxidative Med. Cell. Longev. 2016, 2016, 1–10. [Google Scholar] [CrossRef]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef] [Green Version]
- Swanson, K.V.; Deng, M.; Ting, J.P.-Y. The NLRP3 inflammasome: Molecular activation and regulation to therapeutics. Nat. Rev. Immunol. 2019, 19, 477–489. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, F.; Zhang, Y. Expression and Contribution of NLRP3 Inflammasome During the Follicular Development Induced by PMSG. Front. Cell Dev. Boil. 2019, 7, 256. [Google Scholar] [CrossRef]
- Zhu, L.; Li, J.; Xing, N.; Han, D.; Kuang, H.; Ge, P. American Ginseng Regulates Gene Expression to Protect against Premature Ovarian Failure in Rats. BioMed Res. Int. 2015, 2015, 767124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.-S.; Kim, M.-R.; Park, Y.; Park, H.J.; Chang, U.J.; Kim, S.Y.; Suh, H.J. Fermenting Red Ginseng Enhances Its Safety and Efficacy as a Novel Skin Care Anti-Aging Ingredient: In Vitro and Animal Study. J. Med. Food 2012, 15, 1015–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Sun, B.; Shi, W.; Zuo, H.; Cui, D.; Ni, L.; Chen, J. Decreasing GSH and increasing ROS in chemosensitivity gliomas with IDH1 mutation. Tumor Boil. 2014, 36, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Jyrkkänen, H.-K.; Kansanen, E.; Inkala, M.; Kivelä, A.; Hurttila, H.; Heinonen, S.E.; Goldsteins, G.; Jauhiainen, S.; Tiainen, S.; Makkonen, H.; et al. Nrf2 Regulates Antioxidant Gene Expression Evoked by Oxidized Phospholipids in Endothelial Cells and Murine Arteries In Vivo. Circ. Res. 2008, 103, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Hishida, A.; Okada, R.; Naito, M.; Morita, E.; Wakai, K.; Hamajima, N.; Hosono, S.; Nanri, H.; Turin, T.C.; Suzuki, S.; et al. Polymorphisms in genes encoding antioxidant enzymes (SOD2, CAT, GPx, TXNRD, SEPP1, SEP15 and SELS) and risk of chronic kidney disease in Japanese—Cross-sectional data from the J-MICC study. J. Clin. Biochem. Nutr. 2013, 53, 15–20. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward(5′-3′) | Reverse(5′-3′) |
|---|---|---|
| Idh1 | CCAGTTTGAAGCTCAGAAGA | TACCGTCTTACCATCTGGAC |
| Nrf2 | GCCCACATTCCCAAACAAGAT | CCAGAGAGCTATTGAGGGACTG |
| Gclm | GCATCACGATTCTCTATGCCTAA | TCCAGCTGTGCAACTCCAAGG |
| Nqo1 | AGGATGGGAGGTACTCGAATC | AGGCGTCCTTCCTTATATGCTA |
| Nox2 | GACCCAGATGCAGGAAAGGAA | TCATGGTGCACAGCAAAGTGAT |
| Gpx1 | CTCTCCGCGGCGGCACAGT | CCGCCACCAGGTCGGACGTAC |
| Gr | CACGACCATGATTCCAGATG | CAGCATAGACGCCTTTGACA |
| Sod2 | CTGAGGAGAGCAGCGGTCGT | CTTGGCCAGAGCCTCGTGGT |
| Catalase | TCCGAGATCTTTTCAATGCCATCG | TCGAGCGCGGTAGGGACAGTTCAC |
| Nlrp3 | ACCAGCCAGAGTGGAATGAC | ATAGAGAGGGCGTAGAGGTA |
| Asc | TCACAGAAGTGGACGGAGTG | TCATCTTGTCTTGGCTGGTG |
| pro-IL-1β | TTGACGGACCCCAAAAGATG | AGGACAGCCCAGGTCAAAG |
| IL-1β | CAGGATGAGGACATGAGCAC | CTCTGCACACTCAAACTCCA |
| pro-Caspase-1 | TGGTCTTGTGACTTGGAGGA | ACGACTCTTAGCACGGTTAT |
| Caspase-1 | ATGCCGTGGAGAGAAACAAG | GGTGTTGAAGAGCAGAAAGCA |
| iNOS | AACGGAGAACGTTGGATTTG | CAGCACAAGGGGTTTTCTTC |
| Cox2 | CACTACATCCAGACCCACTT | ATGCTCCTGCTTGAGTATGT |
| 18S | CGGCGACGACCCATTCGAAC | GAATCGAACCCTGATTCCCCGTC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chei, S.; Oh, H.-J.; Jang, H.; Lee, K.; Jin, H.; Choi, Y.; Lee, B.-Y. Korean Red Ginseng Suppresses the Expression of Oxidative Stress Response and NLRP3 Inflammasome Genes in Aged C57BL/6 Mouse Ovaries. Foods 2020, 9, 526. https://doi.org/10.3390/foods9040526
Chei S, Oh H-J, Jang H, Lee K, Jin H, Choi Y, Lee B-Y. Korean Red Ginseng Suppresses the Expression of Oxidative Stress Response and NLRP3 Inflammasome Genes in Aged C57BL/6 Mouse Ovaries. Foods. 2020; 9(4):526. https://doi.org/10.3390/foods9040526
Chicago/Turabian StyleChei, Sungwoo, Hyun-Ji Oh, Hoon Jang, Kippeum Lee, Heegu Jin, Youngsok Choi, and Boo-Yong Lee. 2020. "Korean Red Ginseng Suppresses the Expression of Oxidative Stress Response and NLRP3 Inflammasome Genes in Aged C57BL/6 Mouse Ovaries" Foods 9, no. 4: 526. https://doi.org/10.3390/foods9040526
APA StyleChei, S., Oh, H. -J., Jang, H., Lee, K., Jin, H., Choi, Y., & Lee, B. -Y. (2020). Korean Red Ginseng Suppresses the Expression of Oxidative Stress Response and NLRP3 Inflammasome Genes in Aged C57BL/6 Mouse Ovaries. Foods, 9(4), 526. https://doi.org/10.3390/foods9040526