Bacillus subtilis Fermentation of Malva verticillata Leaves Enhances Antioxidant Activity and Osteoblast Differentiation



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extraction of the M. verticillata Leaves
2.3. Isolation and Identification of the Bacteria from M. verticillata Leaves
2.4. Preparation of M. verticillata Leaf Fermented Aqueous Extracts
2.4.1. Biotransformation of M. verticillata Leaves by Bacillus spp.
2.4.2. Biotransformation of M. verticillata Leaves by Aspergillus oryzae
2.4.3. Biotransformation of Aqueous Extracts of M. verticillata Leaves by B. subtilis MV1
2.5. Determination of Phenolic and Flavonoid Contents
2.6. Antioxidant Activity Assay
2.7. Cell Culture and Cell Viability Assay
2.8. Inhibition Activity of Osteoclast Differentiation
2.9. Alkaline Phosphatase (ALP) Activity Assay and ALP Staining
2.10. Mineralized Nodule Measurement
2.11. Western Blotting
2.12. Total RNA Isolation and RT-PCR
2.13. Liquid Chromatography and Mass Spectrometry Analysis
2.14. Statistical Analysis
3. Results
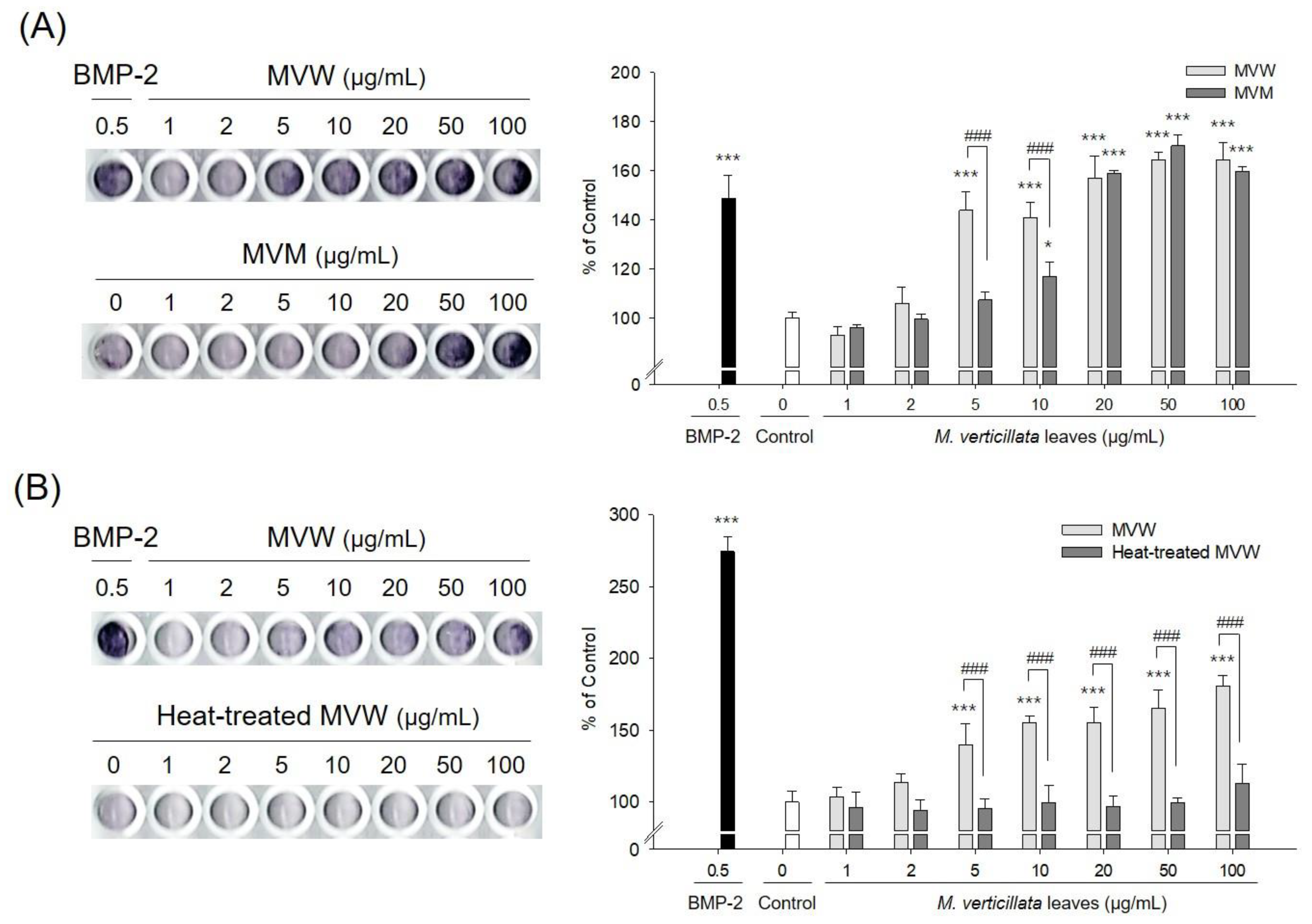
3.1. Methanol and Aqueous Extracts of M. verticillata Leaves Exhibit Osteoblast Differentiation
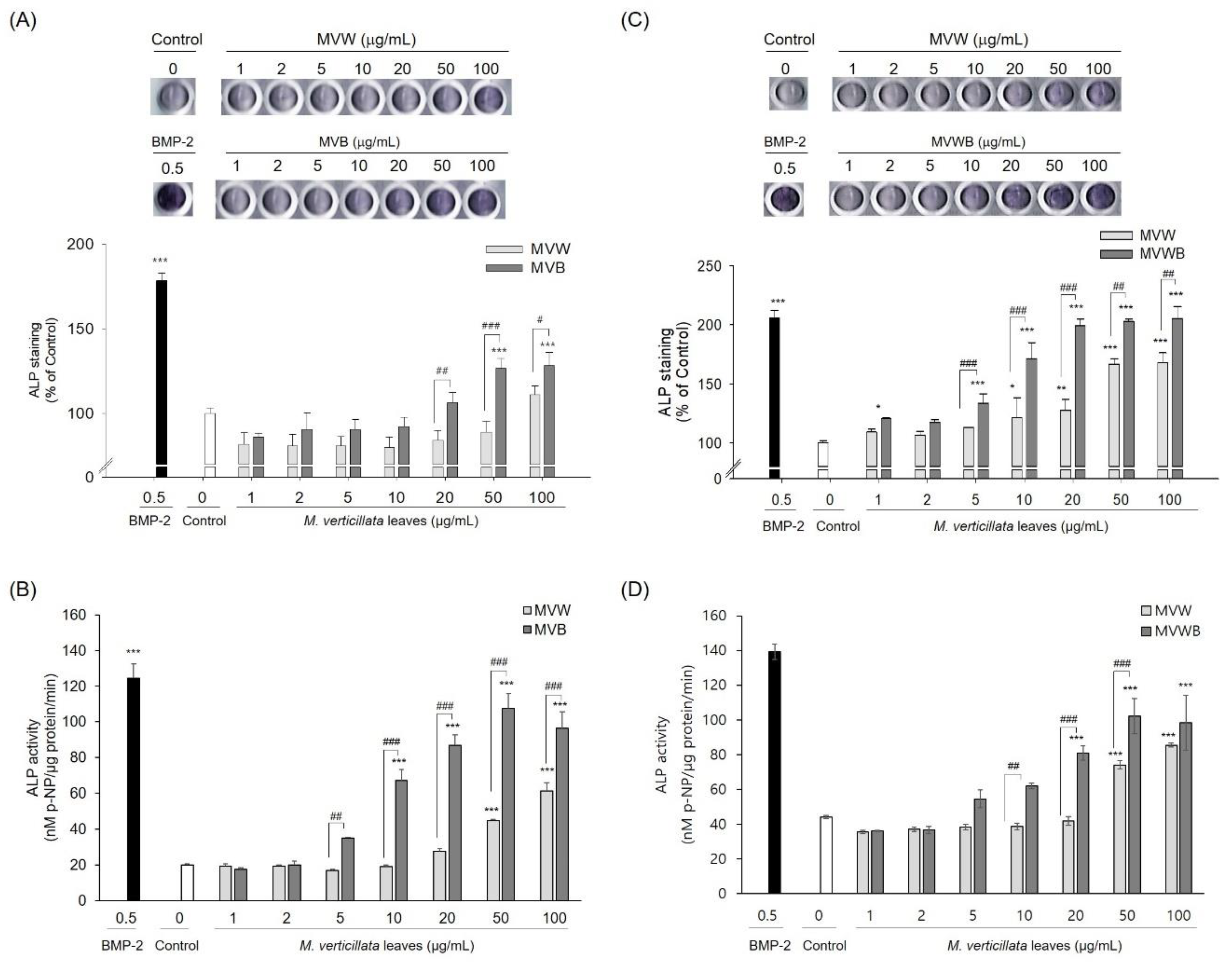
3.2. Biotransformation of the M. verticillata Leaves by B. subtilis MV1 Enhances the Phenolic and Flavonoid Compounds and the Antioxidant Activities
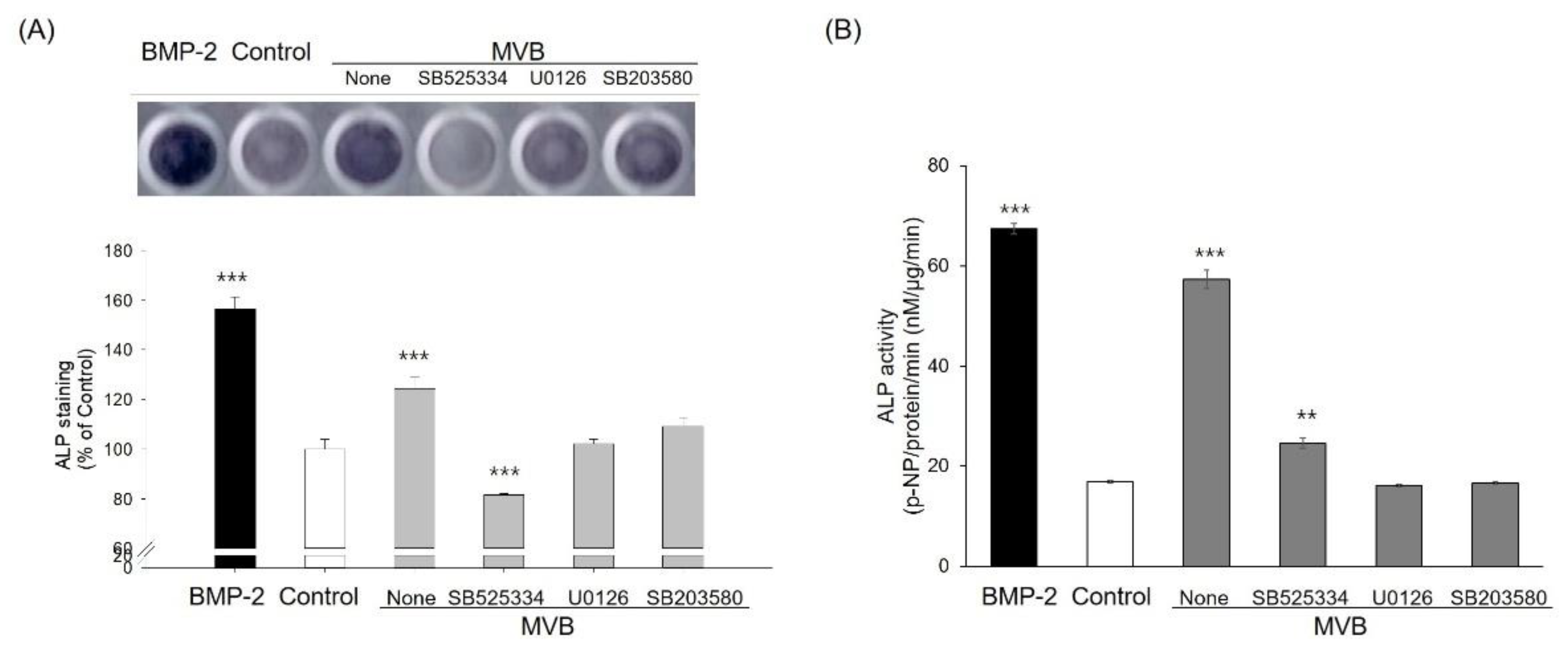
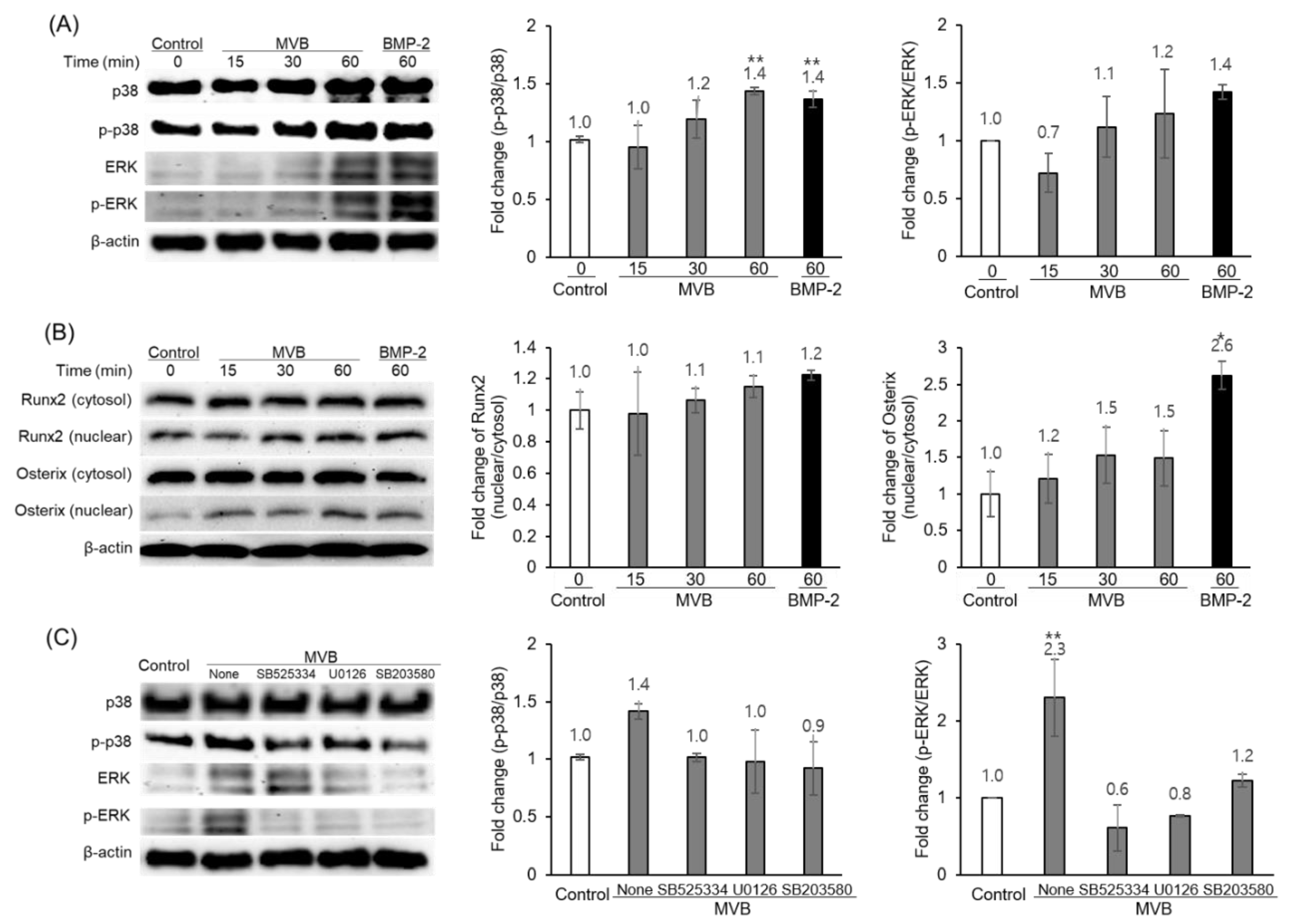
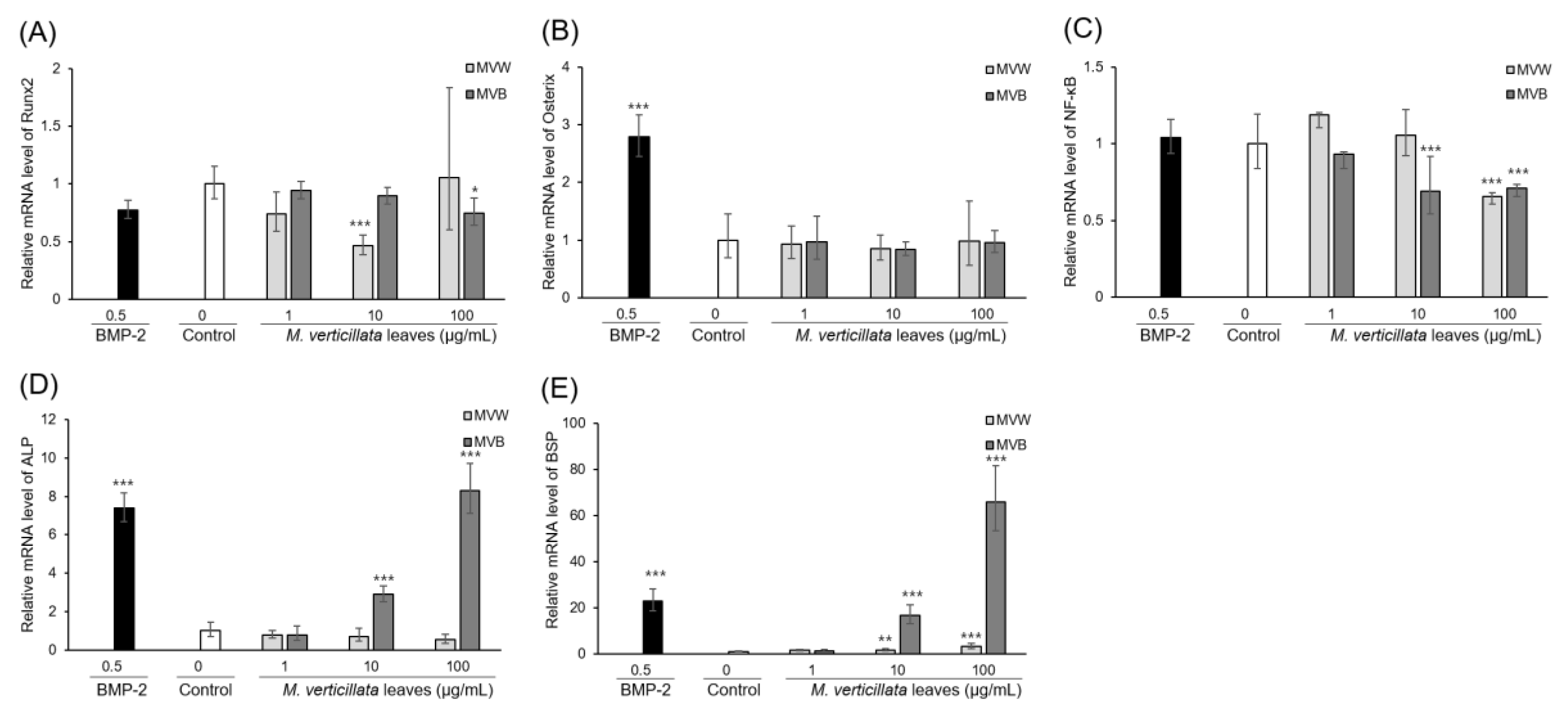
3.3. Biotransformation of M. verticillata Leaves by the Genus Bacillus Enhances the Osteoblast Differentiation of C3H10T1/2 Cells and the Osteoblast Differentiation Occurs via the TGF-β Signaling Pathway
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yedavally-Yellayi, S.; Ho, A.M.; Patalinghug, E.M. Update on osteoporosis. Prim. Care 2019, 46, 175–190. [Google Scholar] [CrossRef]
- Alves, R.R. Osteoblast Differentiation and Bone: Relevant Proteins, Regulatory Processes and the Vascular Connection. Ph.D. Thesis, Erasmus University, Rotterdam, The Netherlands, 5 September 2012. [Google Scholar]
- de Oliveira, L.P.; Bovini, M.G.; da Costa Bortoluzzi, R.L.; Boff, M.I.C.; Boff, P. Species of Malva L.(Malvaceae) cultivated in the western of santa Catarina state and conformity with species marketed as medicinal plants in southern Brazil. J. Agric. Sci. 2019, 11, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Azab, A. Malva: Food, medicine and chemistry. Eur. Chem. Bull. 2017, 6, 295–320. [Google Scholar] [CrossRef] [Green Version]
- Shim, K.S.; Lee, C.J.; Yim, N.H.; Ha, H.; Ma, J.Y. A water extract of Malva verticillata seeds suppresses osteoclastogenesis and bone resorption stimulated by RANK ligand. BMC Complement. Altern. Med. 2016, 16, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.Y.; Choi, E.J.; Kim, J.A.; Hwang, Y.L.; Kim, C.D.; Lee, M.H.; Roh, S.S.; Kim, Y.H.; Han, I.; Kang, S. Malva verticillata seed extracts upregulate the Wnt pathway in human dermal papilla cells. Int. J. Cosmet. Sci. 2016, 38, 148–154. [Google Scholar] [CrossRef]
- Bao, L.; Bao, X.; Li, P.; Wang, X.; Ao, W. Chemical profiling of Malva verticillata L. by UPLC-Q-TOF-MSE and their antioxidant activity in vitro. J. Pharm. Biomed. Anal. 2018, 150, 420–426. [Google Scholar] [CrossRef]
- Madeira Junior, J.V.; Teixeira, C.B.; Macedo, G.A. Biotransformation and bioconversion of phenolic compounds obtainment: An overview. Crit. Rev. Biotechnol. 2013, 35, 75–81. [Google Scholar] [CrossRef]
- Hegazy, M.E.; Mohamed, T.A.; ElShamy, A.I.; Mohamed, A.E.; Mahalel, U.A.; Reda, E.H.; Shaheen, A.M.; Tawfik, W.A.; Shahat, A.A.; Shams, K.A.; et al. Microbial biotransformation as a tool for drug development based on natural products from mevalonic acid pathway: A review. J. Adv. Res. 2015, 6, 17–33. [Google Scholar] [CrossRef] [Green Version]
- Bianchini, L.F.; Arruda, M.F.; Vieira, S.R.; Campelo, P.M.; Gregio, A.M.; Rosa, E.A. Microbial biotransformation to obtain new antifungals. Front. Microbiol. 2015, 6, 1433. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Yan, L.; Wu, Z.; Li, S.; Bai, Z.; Yan, X.; Wang, N.; Liang, N.; Li, H. A microbial transformation using Bacillus subtilis B7-S to produce natural vanillin from ferulic acid. Sci. Rep. 2016, 6, 20400. [Google Scholar] [CrossRef]
- Seong, J.S.; Xuan, S.H.; Park, S.H.; Lee, K.S.; Park, Y.M.; Park, S.N. Antioxidative and antiaging activities and component analysis of Lespedeza cuneata G. Don extracts fermented with Lactobacillus pentosus. J. Microbiol. Biotechnol. 2017, 27, 1961–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razak, D.L.A.; Rashid, N.Y.A.; Jamaluddin, A.; Sharifudin, S.A.; Kahar, A.A.; Long, K. Cosmeceutical potentials and bioactive compounds of rice bran fermented with single and mix culture of Aspergillus oryzae and Rhizopus oryzae. J. Saudi Soc. Agric. Sci. 2017, 16, 127–134. [Google Scholar]
- De Carvalho, C.C.; da Fonseca, M.M. Biotransformation of terpenes. Biotechnol. Adv. 2006, 24, 134–142. [Google Scholar] [CrossRef]
- Velićanski, A.S.; Cvetković, D.D.; Markov, S.L.; Šaponjac, V.T.T.; Vulić, J.J. Antioxidant and antibacterial activity of the beverage obtained by fermentation of sweetened lemon balm (Melissa officinalis L.) tea with symbiotic consortium of bacteria and yeasts. Food Technol. Biotechnol. 2014, 52, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Gong, J.; Chisti, Y.; Sirisansaneeyakul, S. Bioconversion of tea polyphenols to bioactive theabrownins by Aspergillus fumigatus. Biotechnol. Lett. 2014, 36, 2515–2522. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Xu, X.; Wu, Y.; Niu, T.; Liu, Y.; Li, J.; Du, G.; Liu, L. Advances and prospects of Bacillus subtilis cellular factories: From rational design to industrial applications. Metab. Eng. 2018, 50, 109–121. [Google Scholar] [CrossRef]
- Dajanta, K.; Janpum, P.; Leksing, W. Antioxidant capacities, total phenolics and flavonoids in black and yellow soybeans fermented by Bacillus subtilis: A comparative study of Thai fermented soybeans (thua nao). Int. Food Res. J. 2013, 20, 3125. [Google Scholar]
- Moon, K.; Cha, J. Enhancement of antioxidant and antibacterial activities of Salvia miltiorrhiza roots fermented with Aspergillus oryzae. Foods 2020, 9, 34. [Google Scholar] [CrossRef] [Green Version]
- Song, S.H.; Zhai, Y.K.; Li, C.Q.; Yu, Q.; Lu, Y.; Zhang, Y.; Hua, W.P.; Wang, Z.Z.; Shang, P. Effects of total flavonoids from Drynariae Rhizoma prevent bone loss in vivo and in vitro. Bone Rep. 2016, 5, 262–273. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, W.; Ren, F.; Guo, H. Activation of TGF-β canonical and noncanonical signaling in bovine lactoferrin-induced osteogenic activity of C3H10T1/2 mesenchymal stem cells. Int. J. Mol. Sci. 2019, 20, 2880. [Google Scholar] [CrossRef] [Green Version]
- Sakisaka, Y.; Kanaya, S.; Nakamura, T.; Tamura, M.; Shimauchi, H.; Nemoto, E. p38 MAP kinase is required for Wnt3a-mediated osterix expression independently of Wnt-LRP5/6-GSK3β signaling axis in dental follicle cells. Biochem. Biophys. Res. Commun. 2016, 478, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.R.; Yun, H.S.; Lee, T.K.; Lee, S.; Kim, S.H.; Moon, E.; Park, K.M.; Kim, K.H. Chemical characterization of novel natural products from the roots of asian rice (Oryza sativa) that control adipocyte and osteoblast differentiation. J. Agric. Food Chem. 2018, 66, 2677–2684. [Google Scholar] [CrossRef] [PubMed]
- Farhan, H.; Rammal, H.; Hijazi, A.; Hamad, H.; Daher, A.; Reda, M.; Badran, B. In vitro antioxidant activity of ethanolic and aqueous extracts from crude Malva parviflora L. grown in Lebanon. Asian J. Pharm. Clin. Res. 2012, 5, 234–238. [Google Scholar]
- Rao, V. Phytochemicals: A Global Perspective of Their Role in Nutrition and Health; IntechOpen: Rijeka, Croatia, 2012; pp. 467–486. [Google Scholar]
- Date, T.; Doiguchi, Y.; Nobuta, M.; Shindo, H. Bone morphogenetic protein-2 induces differentiation of multipotent C3H10T1/2 cells into osteoblasts, chondrocytes, and adipocytes in vivo and in vitro. J. Orthop. Sci. 2004, 9, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Tarapore, R.S.; Lim, J.; Tian, C.; Pacios, S.; Xiao, W.; Reid, D.; Guan, H.; Mattos, M.; Yu, B.; Wang, C.Y.; et al. NF-kappaB has a direct role in inhibiting Bmp- and Wnt-induced matrix protein expression. J. Bone Miner. Res. 2016, 31, 52–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.; Basu, S.; Yang, G.; Deb, A.; Hu, M. Oral bioavailability challenges of natural products used in cancer chemoprevention. Prog. Chem. 2013, 25, 1553–1574. [Google Scholar]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Yang, S.; Shao, J.; Li, Y.P. Signaling and transcriptional regulation in osteoblast commitment and differentiation. Front. Biosci. 2013, 12, 3068–3092. [Google Scholar] [CrossRef] [Green Version]
- Byun, M.R.; Sung, M.K.; Kim, A.R.; Lee, C.H.; Jang, E.J.; Jeong, M.G.; Noh, M.; Hwang, E.S.; Hong, J.H. (−)-Epicatechin gallate (ECG) stimulates osteoblast differentiation via Runt-related transcription factor 2 (RUNX2) and transcriptional coactivator with PDZ-binding motif (TAZ)-mediated transcriptional activation. J. Biol. Chem. 2014, 289, 9926–9935. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.L.; Chang, J.K.; Tsai, C.H.; Chien, T.T.C.; Kuo, P.L. Myricetin induces human osteoblast differentiation through bone morphogenetic protein-2/p38 mitogen-activated protein kinase pathway. Biochem. Pharmacol. 2007, 73, 504–514. [Google Scholar] [CrossRef]
- Prouillet, C.; Maziere, J.C.; Maziere, C.; Wattel, A.; Brazier, M.; Kamel, S. Stimulatory effect of naturally occurring flavonols quercetin and kaempferol on alkaline phosphatase activity in MG-63 human osteoblasts through ERK and estrogen receptor pathway. Biochem. Pharmacol. 2004, 67, 1307–1313. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.R.; Nam, J.S. Kaempferol stimulates WNT/beta-catenin signaling pathway to induce differentiation of osteoblasts. J. Nutr. Biochem. 2019, 74, 108228. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Sequence (5’→3’) | Product | ||
|---|---|---|---|---|
| Orientation | Size (Base Pairs) | Reference | ||
| GAPDH | ACCACAGTCCATGCCATCAC | Sense | 452 | [22] |
| TCCACCACCCTGTTGCTGTA | Antisense | |||
| Runx2 | AAGCTGCGGCAAGACAAG | Sense | 61 | [23] |
| TCAAATCTGCAGCTTCAAGG | Antisense | |||
| Osterix | GCTAGAGATCTGAGCCGGGTA | Sense | 56 | [23] |
| AAGAGACCTGGCAAGAGG | Antisense | |||
| ALP | AAACCCAGAACACAAGCATTCC | Sense | 218 | [23] |
| TCCACCAGCAAGAAGAAGCC | Antisense | |||
| BSP | TTGAGTTAGCGGCACTCCAA | Sense | 78 | This study |
| CGTCGCTTTCCTTCACTTTT | Antisense | |||
| NF-kB | ACACGAGGCTACAACTCTGC | Sense | 164 | This study |
| GGTACCCCCAGAGACCTCAT | Antisense | |||
| Sample | TPC (mg GAE/g) | TFC (mg QE/g) | DPPH (mg QE/g) | FRAP (mg FeSO4/g) |
|---|---|---|---|---|
| MVW | 8.6 ± 0.4 | 2.4 ± 0.1 | 1.9 ± 0.2 | 12.3 ± 0.1 |
| MVB | 23.6 ± 0.2 *** | 13.2 ± 0.3 *** | 7.5 ± 0.4 *** | 74.5 ± 1.1 *** |
| MVWB | 10.6 ± 0.4 *** | 3.0 ± 0.2 | 4.1 ± 0.3 *** | 39.7 ± 0.1 *** |
| MVA | 8 ± 0.8 | 1.9 ± 0.1 | 1.4 ± 0.2 | 6.2 ± 0.1 *** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moon, K.; Lee, S.; Cha, J. Bacillus subtilis Fermentation of Malva verticillata Leaves Enhances Antioxidant Activity and Osteoblast Differentiation. Foods 2020, 9, 671. https://doi.org/10.3390/foods9050671
Moon K, Lee S, Cha J. Bacillus subtilis Fermentation of Malva verticillata Leaves Enhances Antioxidant Activity and Osteoblast Differentiation. Foods. 2020; 9(5):671. https://doi.org/10.3390/foods9050671
Chicago/Turabian StyleMoon, Keumok, Seola Lee, and Jaeho Cha. 2020. "Bacillus subtilis Fermentation of Malva verticillata Leaves Enhances Antioxidant Activity and Osteoblast Differentiation" Foods 9, no. 5: 671. https://doi.org/10.3390/foods9050671
APA StyleMoon, K., Lee, S., & Cha, J. (2020). Bacillus subtilis Fermentation of Malva verticillata Leaves Enhances Antioxidant Activity and Osteoblast Differentiation. Foods, 9(5), 671. https://doi.org/10.3390/foods9050671