Allergenicity of Fermented Foods: Emphasis on Seeds Protein-Based Products

 , , , and
, , , and
Abstract
:1. Introduction
2. Food Allergy
2.1. Allergic Reactions and Prevalence
2.2. Seed Protein Allergens
3. Food Processing and Protein Allergenicity
4. Fermented Foods and Allergenicity
4.1. Fermented Dairy Products and Milk Allergy
4.2. Seeds Protein-Based Products
4.2.1. Soy Fermented Products
4.2.2. Other Cereal-Based Fermented Foods
4.3. Modulation of Immune Responses by Fermented Foods: In Vivo Studies
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Prescott, S.L.; Pawankar, R.; Allen, K.J.; Campbell, D.E.; Sinn, J.K.H.; Fiocchi, A.; Ebisawa, M.; Sampson, H.A.; Beyer, K.; Lee, B.W. A global survey of changing patterns of food allergy burden in children. World Allergy Organ. J. 2013, 6, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du Toit, G.; Roberts, G.; Sayre, P.H.; Plaut, M.; Bahnson, H.T.; Mitchell, H.; Radulovic, S.; Chan, S.; Fox, A.; Turcanu, V.; et al. Identifying infants at high risk of peanut allergy: The learning early about peanut allergy (LEAP) screening study. J. Allergy Clin. Immunol. 2013, 131, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Sicherer, S.H.; Sampson, H.A. Food allergy: Epidemiology, pathogenesis, diagnosis, and treatment. J. Allergy Clin. Immunol. 2014, 133, 291–307. [Google Scholar] [CrossRef] [PubMed]
- Fox, M.; Mugford, M.; Voordouw, J.; Cornelisse-Vermaat, J.; Antonides, G.; Caballer, B.D.L.H.; Cerecedo, I.; Zamora, J.; Rokicka, E.; Jewczak, M.; et al. Health sector costs of self-reported food allergy in Europe: A patient-based cost of illness study. Eur. J. Public Health 2013, 23, 757–762. [Google Scholar] [CrossRef] [Green Version]
- Pelz, B.J.; Bryce, P.J. Pathophysiology of food allergy. Pediatr. Clin. 2015, 62, 1363–1375. [Google Scholar] [CrossRef]
- Sicherer, S.H.; Sampson, H.A. Food allergy: A review and update on epidemiology, pathogenesis, diagnosis, prevention, and management. J. Allergy Clin. Immunol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, C.A.; Cannon, G.; Levy, R.; Moubarac, J.C.; Jaime, P.; Martins, A.P.; Canella, D.; Louzada, M.; Parra, D. NOVA. The star shines bright. World Nutr. 2016, 7, 28–38. [Google Scholar]
- Moubarac, J.-C.; Martins, A.P.B.; Claro, R.M.; Levy, R.B.; Cannon, G.; Monteiro, C.A. Consumption of ultra-processed foods and likely impact on human health. Evidence from Canada. Public Health Nutr. 2013, 16, 2240–2248. [Google Scholar] [CrossRef] [Green Version]
- Canella, D.S.; Levy, R.B.; Martins, A.P.B.; Claro, R.M.; Moubarac, J.-C.; Baraldi, L.G.; Cannon, G.; Monteiro, C.A. Ultra-processed food products and Obesity in Brazilian Households (2008–2009). PLoS ONE 2014, 9, e92752. [Google Scholar] [CrossRef] [Green Version]
- Fiolet, T.; Srour, B.; Sellem, L.; Kesse-Guyot, E.; Allès, B.; Méjean, C.; Deschasaux, M.; Fassier, P.; Latino-Martel, P.; Beslay, M.; et al. Consumption of ultra-processed foods and cancer risk: Results from NutriNet-Santé prospective cohort. BMJ 2018, 360. [Google Scholar] [CrossRef] [Green Version]
- Marrón-Ponce, J.A.; Flores, M.; Cediel, G.; Monteiro, C.A.; Batis, C. Associations between Consumption of Ultra-processed foods and intake of nutrients related to chronic non-communicable diseases in Mexico. J. Acad. Nutr. Diet. 2019, 119, 1852–1865. [Google Scholar] [CrossRef]
- Fardet, A.; Rock, E. Ultra-processed foods: A new holistic paradigm? Trends Food Sci. Technol. 2019, 93, 174–184. [Google Scholar] [CrossRef]
- Villemin, C.; Tranquet, O.; Solé, V.; Smit, J.; Pieters, R.; Denery, S.; Bouchaud, G. Deamidation and enzymatic hydrolysis of gliadins alter their processing by dendritic cells in vitro. J. Agric. Food Chem. 2019, 1447–1456. [Google Scholar] [CrossRef] [PubMed]
- Taheri-Kafrani, A.; Gaudin, J.-C.; Rabesona, H.; Nioi, C.; Agarwal, D.; Drouet, M.; Chobert, J.-M.; Bordbar, A.-K.; Haertle, T. Effects of heating and glycation of β-Lactoglobulin on its recognition by IgE of sera from cow milk allergy patients. J. Agric. Food Chem. 2009, 57, 4974–4982. [Google Scholar] [CrossRef] [PubMed]
- Rahaman, T.; Vasiljevic, T.; Ramchandran, L. Effect of processing on conformational changes of food proteins related to allergenicity. Trends Food Sci. Technol. 2016, 49, 24–34. [Google Scholar] [CrossRef]
- Verhoeckx, K.C.M.; Vissers, Y.M.; Baumert, J.L.; Faludi, R.; Feys, M.; Flanagan, S.; Herouet-Guicheney, C.; Holzhauser, T.; Shimojo, R.; van der Bolt, N.; et al. Food processing and allergenicity. Food Chem. Toxicol. 2015, 80, 223–240. [Google Scholar] [CrossRef] [PubMed]
- Sathe, S.K.; Liu, C.; Zaffran, V.D. Food Allergy. Annu. Rev. Food Sci. Technol. 2016, 7, 191–220. [Google Scholar] [CrossRef]
- Tranquet, O.; Larré, C.; Denery-Papini, S. Allergic reactions to hydrolysed wheat proteins: Clinical aspects and molecular structures of the allergens involved. Crit. Rev. Food Sci. Nutr. 2018, 60, 1–10. [Google Scholar] [CrossRef]
- Chilton, S.; Burton, J.; Reid, G.; Chilton, S.N.; Burton, J.P.; Reid, G. Inclusion of fermented foods in food guides around the world. Nutrients 2015, 7, 390–404. [Google Scholar] [CrossRef] [Green Version]
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligné, B.; Gänzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef]
- Tamang, J.P.; Shin, D.H.; Jung, S.J.; Chae, S.W. Functional properties of microorganisms in fermented foods. Front. Microbiol. 2016, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iweala, O.I.; Nagler, C.R. The Microbiome and Food Allergy. Annu. Rev. Immunol. 2019, 37, 377–403. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Amigo, C.; Popping, B. Food allergens: A regulatory/labelling overview including the VITAL approach. Encycl. Food Chem. 2019, 615–621. [Google Scholar] [CrossRef]
- Nwaru, B.I.; Hickstein, L.; Panesar, S.S.; Roberts, G.; Muraro, A.; Sheikh, A.; EAACI Food Allergy and Anaphylaxis Guidelines Group. Prevalence of common food allergies in Europe: A systematic review and meta-analysis. Allergy 2014, 69, 992–1007. [Google Scholar] [CrossRef]
- Dunlop, J.H.; Keet, C.A. Epidemiology of food allergy. Immunol. Allergy Clin. North Am. 2018, 38, 13–25. [Google Scholar] [CrossRef]
- Leung, A.S.Y.; Wong, G.W.K.; Tang, M.L.K. Food allergy in the developing world. J. Allergy Clin. Immunol. 2018, 141, 76–78.e1. [Google Scholar] [CrossRef] [Green Version]
- Soller, L.; Ben-Shoshan, M.; Harrington, D.W.; Knoll, M.; Fragapane, J.; Joseph, L.; Pierre, Y.S.; La Vieille, S.; Wilson, K.; Elliott, S.J.; et al. Adjusting for nonresponse bias corrects overestimates of food allergy prevalence. J. Allergy Clin. Immunol. Pract. 2015, 3, 291–293.e2. [Google Scholar] [CrossRef] [PubMed]
- Živanović, M.; Atanasković-Marković, M.; Medjo, B.; Gavrović-Jankulović, M.; Smiljanić, K.; Tmušić, V.; Djurić, V. Evaluation of food allergy in children by skin prick tests with commercial extracts and fresh foods, specific IgE and, open oral food challenge-our five years experience in food allergy work-up. Iran. J. Allergy Asthma Immunol. 2017, 16, 127–132. [Google Scholar] [PubMed]
- Acker, W.W.; Plasek, J.M.; Blumenthal, K.G.; Lai, K.H.; Topaz, M.; Seger, D.L.; Goss, F.R.; Slight, S.P.; Bates, D.W.; Zhou, L. Prevalence of food allergies and intolerances documented in electronic health records. J. Allergy Clin. Immunol. 2017, 140, 1587–1591.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.S.; Springston, E.E.; Warrier, M.R.; Smith, B.; Kumar, R.; Pongracic, J.; Holl, J.L. The prevalence, severity, and distribution of childhood food allergy in the United States. Pediatrics 2011, 128, 9–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radauer, C.; Bublin, M.; Wagner, S.; Mari, A.; Breiteneder, H. Allergens are distributed into few protein families and possess a restricted number of biochemical functions. J. Allergy Clin. Immunol. 2008, 121, 847–852. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Bahna, S.L. Hypersensitivities to sesame and other common edible seeds. Allergy Eur. J. Allergy Clin. Immunol. 2016, 71, 1405–1413. [Google Scholar] [CrossRef] [PubMed]
- Osborne, N.J.; Koplin, J.J.; Martin, P.E.; Gurrin, L.C.; Lowe, A.J.; Matheson, M.C.; Ponsonby, A.L.; Wake, M.; Tang, M.L.K.; Dharmage, S.C.; et al. Prevalence of challenge-proven IgE-mediated food allergy using population-based sampling and predetermined challenge criteria in infants. J. Allergy Clin. Immunol. 2011, 127, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Richard, C.; Jacquenet, S.; Sergeant, P.; Moneret-Vautrin, D.A. Cross-reactivity of a new food ingredient, dun pea, with legumes, and risk of anaphylaxis in legume allergic children. Eur. Ann. Allergy Clin. Immunol. 2015, 47, 118–125. [Google Scholar]
- Just, J.; Deschildre, A.; Beaudouin, E. Allergies Alimentaires: Nouveaux Concepts, Affections Actuelles, Perspectives Thérapeutiques; Elsevier: Paris, France, 2017; ISBN 9782294750137. [Google Scholar]
- Kim, M.; Lee, J.Y.; Jeon, H.; Yang, H.; Lee, K.-J.; Han, Y.; Kim, Y.H.; Kim, J.; Ahn, K. Prevalence of immediate-type food allergy in Korean schoolchildren in 2015: A Nationwide, Population-based Study. Allergy. Asthma Immunol. Res. 2017, 9, 410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ukleja-Sokołowska, N.; Gawrońska-Ukleja, E.; Zbikowska-Gotz, M.; Bartuzi, Z.; Sokołowski, Ł. Sunflower seed allergy. Int. J. Immunopathol. Pharmacol. 2016, 29, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Jensen-Jarolim, E.; Gerstmayer, G.; Kraft, D.; Scheiner, O.; Ebner, H.; Ebner, C. Serological characterization of allergens in poppy seeds. Clin. Exp. Allergy 1999, 29, 1075–1079. [Google Scholar] [CrossRef]
- Lucassen, R.; Fooke, M.; Kleine-Tebbe, J.; Mahler, M. Development and evaluation of a rapid assay for the diagnosis of immunoglobulin E-mediated type I allergies. J. Investig. Allergol. Clin. Immunol. 2008, 18, 223–224. [Google Scholar] [CrossRef]
- Valverde-Monge, M.; Bartolome, B.; Custa-Herranz, J.; Heras, M.D.L. Allergy to pumpkin seeds. J. Allergy Clin. Immunol. 2017, 139, AB138. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.; Park, S.-Y.; Noh, S.; Kim, J.; Seo, B.; Kwon, Y.; Kwon, H.-S.; Cho, S.; Moon, H.-B.; Kim, T.-B. Case report: A first case of flaxseed-induced anaphylaxis in Korea. Medicine 2017, 96, e8220. [Google Scholar] [CrossRef]
- Masilamani, M.; Commins, S.; Shreffler, W. Determinants of Food Allergy. Immunol. Allergy Clin. N. Am. 2012, 32, 2–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Sathe, S.K. Food Allergen Epitope Mapping. J. Agric. Food Chem. 2018, 66, 7238–7248. [Google Scholar] [CrossRef] [PubMed]
- Frank, R. The SPOT-synthesis technique: Synthetic peptide arrays on membrane supports—Principles and applications. J. Immunol. Methods 2002, 267, 13–26. [Google Scholar] [CrossRef]
- Kühne, Y.; Reese, G.; Ballmer-Weber, B.K.; Niggemann, B.; Hanschmann, K.M.; Vieths, S.; Holzhauser, T. A novel multipeptide microarray for the specific and sensitive mapping of linear IgE-binding epitopes of food allergens. Int. Arch. Allergy Immunol. 2015, 166, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Volpicella, M.; Leoni, C.; Dileo, M.C.G.; Ceci, L.R. Progress in the analysis of food allergens through molecular biology approaches. Cells 2019, 8, 1073. [Google Scholar] [CrossRef] [Green Version]
- Saeed, H.; Gagnon, C.; Cober, E.; Gleddie, S. Using patient serum to epitope map soybean glycinins reveals common epitopes shared with many legumes and tree nuts. Mol. Immunol. 2016, 70, 125–133. [Google Scholar] [CrossRef]
- Verhoeckx, K.; Bøgh, K.L.; Dupont, D.; Egger, L.; Gadermaier, G.; Larré, C.; Mackie, A.; Menard, O.; Adel-Patient, K.; Picariello, G.; et al. The relevance of a digestibility evaluation in the allergenicity risk assessment of novel proteins. Opinion of a joint initiative of COST action ImpARAS and COST action INFOGEST. Food Chem. Toxicol. 2019, 129, 405–423. [Google Scholar] [CrossRef] [PubMed]
- Beyer, K.; Morrowa, E.; Li, X.-M.; Bardina, L.; Bannon, G.A.; Burks, A.W.; Sampson, H.A. Effects of cooking methods on peanut allergenicity. J. Allergy Clin. Immunol. 2001, 107, 1077–1081. [Google Scholar] [CrossRef]
- Palladino, C.; Breiteneder, H. Peanut allergens. Mol. Immunol. 2018, 100, 58–70. [Google Scholar] [CrossRef]
- Tian, Y.; Rao, H.; Zhang, K.; Tao, S.; Xue, W.-T. Effects of different thermal processing methods on the structure and allergenicity of peanut allergen Ara h 1. Food Sci. Nutr. 2018, 6, 1706–1714. [Google Scholar] [CrossRef]
- Turner, P.J.; Mehr, S.; Sayers, R.; Wong, M.; Shamji, M.H.; Campbell, D.E.; Mills, E.N.C. Loss of allergenic proteins during boiling explains tolerance to boiled peanut in peanut allergy. J. Allergy Clin. Immunol. 2014, 134, 751–753. [Google Scholar] [CrossRef]
- Mondoulet, L.; Paty, E.; Drumare, M.F.; Ah-Leung, S.; Scheinmann, P.; Willemot, R.M.; Wal, J.M.; Bernard, H. Influence of thermal processing on the allergenicity of peanut proteins. J. Agric. Food Chem. 2005, 53, 4547–4553. [Google Scholar] [CrossRef]
- Cabanillas, B.; Maleki, S.J.; Rodríguez, J.; Burbano, C.; Muzquiz, M.; Jiménez, M.A.; Pedrosa, M.M.; Cuadrado, C.; Crespo, J.F. Heat and pressure treatments effects on peanut allergenicity. Food Chem. 2012, 132, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Ahmedna, M.; Goktepe, I.; Cheng, H.; Maleki, S. Enzymatic treatment of peanut kernels to reduce allergen levels. Food Chem. 2011, 127, 1014–1022. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado, C.; Cabanillas, B.; Pedrosa, M.M.; Muzquiz, M.; Haddad, J.; Allaf, K.; Rodriguez, J.; Crespo, J.F.; Burbano, C. Effect of instant controlled pressure drop on IgE antibody reactivity to peanut, lentil, chickpea and soybean proteins. Int. Arch. Allergy Immunol. 2011, 156, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, A.; Boye, J.I. Impact of thermal processing time and cookie size on the detection of casein, egg, gluten and soy allergens in food. Food Res. Int. 2013, 52, 483–489. [Google Scholar] [CrossRef]
- Frias, J.; Song, Y.S.; Martínez-Villaluenga, C.; De Mejia, E.G.; Vidal-Valverde, C. Fermented soyabean products as hypoallergenic food. Proc. Nutr. Soc. 2008, 67, 2008. [Google Scholar] [CrossRef] [Green Version]
- Takács, K.; Guillamon, E.; Pedrosa, M.M.; Cuadrado, C.; Burbano, C.; Muzquiz, M.; Haddad, J.; Allaf, K.; Maczó, A.; Polgár, M.; et al. Study of the effect of instant controlled pressure drop (DIC) treatment on IgE-reactive legume-protein patterns by electrophoresis and immunoblot. Food Agric. Immunol. 2014, 25, 173–185. [Google Scholar] [CrossRef]
- Pastorello, E.A.; Farioli, L.; Conti, A.; Pravettoni, V.; Bonomi, S.; Iametti, S.; Fortunato, D.; Scibilia, J.; Bindslev-Jensen, C.; Ballmer-Weber, B.; et al. Wheat IgE-mediated food allergy in european patients: α-amylase inhibitors, lipid transfer proteins and low-molecular-weight glutenins. Int. Arch. Allergy Immunol. 2007, 144, 10–22. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Rizzello, C.G.; Scala, E.; De Simone, C.; Farris, G.A.; Turrini, F.; Gobbetti, M. Probiotic preparation has the capacity to hydrolyze proteins responsible for wheat allergy. J. Food Prot. 2007, 70, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Álvarez, J.; Guillamón, E.; Crespo, J.F.; Cuadrado, C.; Burbano, C.; Rodríguez, J.; Fernández, C.; Muzquiz, M. Effects of extrusion, boiling, autoclaving, and microwave heating on lupine allergenicity. J. Agric. Food Chem. 2005, 53, 1294–1298. [Google Scholar] [CrossRef] [PubMed]
- Guillamón, E.; Burbano, C.; Cuadrado, C.; Muzquiz, M.; Pedrosa, M.M.; Sánchez, M.; Cabanillas, B.; Crespo, J.F.; Rodriguez, J.; Haddad, J.; et al. Effect of an instantaneous controlled pressure drop on in vitro allergenicity to Lupins (Lupinus albus var. Multolupa). Int. Arch. Allergy Immunol. 2008, 145, 9–14. [Google Scholar] [CrossRef]
- Cuadrado, C.; Cabanillas, B.; Pedrosa, M.M.; Varela, A.; Guillamón, E.; Muzquiz, M.; Crespo, J.F.; Rodriguez, J.; Burbano, C. Influence of thermal processing on IgE reactivity to lentil and chickpea proteins. Mol. Nutr. Food Res. 2009, 53, 1462–1468. [Google Scholar] [CrossRef] [PubMed]
- Comstock, S.S.; Maleki, S.J.; Teuber, S.S. Boiling and frying peanuts decreases soluble peanut (Arachis Hypogaea) allergens Ara h 1 and Ara h 2 but does not generate hypoallergenic peanuts. PLoS ONE 2016, 11, e0157849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillon, B.; Bernard, H.; Drumare, M.-F.; Hazebrouck, S.; Adel-Patient, K. Heat processing of peanut seed enhances the sensitization potential of the major peanut allergen Ara h 6. Mol. Nutr. Food Res. 2016, 60, 2722–2735. [Google Scholar] [CrossRef] [Green Version]
- Ekezie, F.G.C.; Cheng, J.H.; Sun, D.W. Effects of nonthermal food processing technologies on food allergens: A review of recent research advances. Trends Food Sci. Technol. 2018, 74, 12–25. [Google Scholar] [CrossRef]
- El Mecherfi, K.E.; Curet, S.; Lupi, R.; Larré, C.; Rouaud, O.; Choiset, Y.; Rabesona, H.; Haertlé, T. Combined microwave processing and enzymatic proteolysis of bovine whey proteins: The impact on bovine β-lactoglobulin allergenicity. J. Food Sci. Technol. 2019, 56, 177–186. [Google Scholar] [CrossRef]
- Bu, G.; Luo, Y.; Chen, F.; Liu, K.; Zhu, T. Milk processing as a tool to reduce cow’s milk allergenicity: A mini-review. Dairy Sci. Technol. 2013, 93, 211–223. [Google Scholar] [CrossRef] [Green Version]
- Meulenbroek, L.A.P.M.; Oliveira, S.; Jager, C.F.D.H.; Klemans, R.J.B.; Lebens, A.F.M.; van Baalen, T.; Knulst, A.C.; Bruijnzeel-Koomen, C.A.F.M.; Garssen, J.; Knippels, L.M.J.; et al. The degree of whey hydrolysis does not uniformly affect in vitro basophil and T cell responses of cow’s milk-allergic patients. Clin. Exp. Allergy 2014, 44, 529–539. [Google Scholar] [CrossRef]
- Shi, X.; Guo, R.; White, B.L.; Yancey, A.; Sanders, T.H.; Davis, J.P.; Burks, A.W.; Kulis, M. Allergenic properties of enzymatically hydrolyzed peanut flour extracts. Int. Arch. Allergy Immunol. 2013, 162, 123–130. [Google Scholar] [CrossRef]
- Kulis, M.; Macqueen, I.; Li, Y.; Guo, R.; Zhong, X.-P.; Burks, A.W. Pepsinized cashew proteins are hypoallergenic and immunogenic and provide effective immunotherapy in mice with cashew allergy. J. Allergy Clin. Immunol. 2012, 130, 716–723. [Google Scholar] [CrossRef] [Green Version]
- McGovern, P.E.; Zhang, J.; Tang, J.; Zhang, Z.; Hall, G.R.; Moreau, R.A.; Nuñez, A.; Butrym, E.D.; Richards, M.P.; Wang, C.-S.; et al. Fermented beverages of pre- and proto-historic China. Proc. Natl. Acad. Sci. USA 2004, 101, 17593–17598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalieri, D.; McGovern, P.E.; Hartl, D.L.; Mortimer, R.; Polsinelli, M. Evidence for S. cerevisiae Fermentation in Ancient Wine. J. Mol. Evol. 2003, 57, S226–S232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legras, J.-L.; Merdinoglu, D.; Cornuet, J.-M.; Karst, F. Bread, beer and wine: Saccharomyces cerevisiae diversity reflects human history. Mol. Ecol. 2007, 16, 2091–2102. [Google Scholar] [CrossRef]
- Penna, A.L.B.; Nero, L.A.; Todorov, S.D. Fermented Foods of Latin America: From Traditional Knowledge to Innovative Applications, 1st ed.; CRC Press: Boca Raton, FL, USA, 2017; ISBN 9781315369433. [Google Scholar]
- Tamang, J.P.; Watanabe, K.; Holzapfel, W.H. Review: Diversity of microorganisms in global fermented foods and beverages. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzello, C.G.; Nionelli, L.; Coda, R.; Gobbetti, M. Synthesis of the cancer preventive peptide lunasin by lactic acid bacteria during sourdough fermentation. Nutr. Cancer 2012, 64, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ledesma, B.; Hsieh, C.C.; de Lumen, B.O. Chemopreventive properties of peptide lunasin: A review. Protein Pept. Lett. 2013, 20, 424–432. [Google Scholar] [PubMed] [Green Version]
- Malaguti, M.; Dinelli, G.; Leoncini, E.; Bregola, V.; Bosi, S.; Cicero, A.; Hrelia, S.; Malaguti, M.; Dinelli, G.; Leoncini, E.; et al. Bioactive peptides in cereals and legumes: Agronomical, biochemical and clinical aspects. Int. J. Mol. Sci. 2014, 15, 21120–21135. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Martinez, M.; Winkler, R.; García-Lara, S.; López-Camarillo, C.; Pando-Robles, V.; Barkla, B.J. Preventive and therapeutic potential of peptides from cereals against cancer. J. Proteomics 2014, 111, 165–183. [Google Scholar] [CrossRef]
- Hole, A.S.; Rud, I.; Grimmer, S.; Sigl, S.; Narvhus, J.; Sahlstrøm, S. Improved bioavailability of dietary phenolic acids in whole grain barley and oat groat following fermentation with probiotic Lactobacillus acidophilus, Lactobacillus johnsonii, and Lactobacillus reuteri. J. Agric. Food Chem. 2012, 60, 6369–6375. [Google Scholar] [CrossRef]
- Hur, S.J.; Lee, S.Y.; Kim, Y.-C.; Choi, I.; Kim, G.-B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, H.; Starrenburg, M.J.C.; Molenaar, D.; Kleerebezem, M.; Vlieg, J.E.V.H. Microbial domestication signatures of Lactococcus lactis can be reproduced by experimental evolution. Genome Res. 2012, 22, 115–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addeo, F.; Chianese, L.; Sacchi, R.; Cappuccio, U.; Ferranti, P.; Malorni, A. Characterization of the 12% trichloroacetic acid-insoluble oligopeptides of Parmigiano-Reggiano cheese. J. Dairy Res. 1992, 59, 401–411. [Google Scholar] [CrossRef]
- Addeo, F.; Chianese, L.; Sacchi, R.; Musso, S.S.; Ferranti, P.; Malorni, A. Characterization of the oligopeptides of Parmigiano-Reggiano cheese soluble in 120 g trichloroacetic acid/1. J. Dairy Res. 1994, 61, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Ferranti, P.; Itolli, E.; Barone, F.; Malorni, A.; Garro, G.; Laezza, P.; Chianese, L.; Migliaccio, F.; Stingo, V.; Addeo, F. Combined high resolution chromatographic techniques (FPLC and HPLC) and mass spectrometry-based identification of peptides and proteins in Grana Padano cheese. Lait 1997, 77, 683–697. [Google Scholar] [CrossRef]
- Singh, T.K.; Fox, P.F.; Healy, Á. Isolation and identification of further peptides in the diafiltration retentate of the water-soluble fraction of Cheddar cheese. J. Dairy Res. 1997, 64, 433–443. [Google Scholar] [CrossRef]
- Atanasova, J.; Moncheva, P.; Ivanova, I. Proteolytic and antimicrobial activity of lactic acid bacteria grown in goat milk. Biotechnol. Biotechnol. Equip. 2014, 28, 1073–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, M.; Wilm, M. Error-Tolerant Identification of peptides in sequence databases by peptide sequence tags. Anal. Chem. 1994, 66, 4390–4399. [Google Scholar] [CrossRef]
- Fox, P.F.; O’Connor, T.P.; Mcsweeney, P.L.H.; Guinee, T.P.; O’Brien, N.M. Cheese: Physical, biochemical, and nutritional aspects. Adv. Food Nutr. Res. 1996, 39, 163–328. [Google Scholar] [CrossRef] [PubMed]
- Sieber, R. Käse—Ein wertvolles Lebensmittel in der menschlichen Ernährung. Mitt. Lebensm. Hyg. 2005, 96, 141–170. [Google Scholar]
- Silva, S.V.; Malcata, F.X. Caseins as source of bioactive peptides. Int. Dairy J. 2005, 15, 1–15. [Google Scholar] [CrossRef]
- Schlimme, E.; Meisel, H. Bioactive peptides derived from milk proteins. Structural, physiological and analytical aspects. Food/Nahrung 1995, 39, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Migliore-Samour, D.; Jolles, P.; Floćh, F. Biologically active casein peptides implicated in immunomodulation. J. Dairy Res. 1989, 56, 357–362. [Google Scholar] [CrossRef] [PubMed]
- McGowan, E.C.; Keet, C.A. Prevalence of self-reported food allergy in the National Health and Nutrition Examination Survey (NHANES) 2007–2010. J. Allergy Clin. Immunol. 2013, 132, 1216–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nwaru, B.I.; Hickstein, L.; Panesar, S.S.; Muraro, A.; Werfel, T.; Cardona, V.; Dubois, A.E.J.; Halken, S.; Hoffmann-Sommergruber, K.; Poulsen, L.K.; et al. The epidemiology of food allergy in Europe: A systematic review and meta-analysis. Allergy Eur. J. Allergy Clin. Immunol. 2014, 69, 62–75. [Google Scholar] [CrossRef] [Green Version]
- Zeng, G.Q.; Luo, J.Y.; Huang, H.M.; Zheng, P.Y.; Luo, W.T.; Wei, N.L.; Sun, B.Q. Food allergy and related risk factors in 2540 preschool children: An epidemiological survey in Guangdong Province, Southern China. World J. Pediatr. 2015, 11, 219–225. [Google Scholar] [CrossRef]
- Schoemaker, A.A.; Sprikkelman, A.B.; Grimshaw, K.E.; Roberts, G.; Grabenhenrich, L.; Rosenfeld, L.; Siegert, S.; Dubakiene, R.; Rudzeviciene, O.; Reche, M.; et al. Incidence and natural history of challenge-proven cow’s milk allergy in European children—EuroPrevall birth cohort. Allergy 2015, 70, 963–972. [Google Scholar] [CrossRef]
- Pescuma, M.; Hébert, E.M.; Rabesona, H.; Drouet, M.; Choiset, Y.; Haertlé, T.; Mozzi, F.; de Valdez, G.F.; Chobert, J.-M. Proteolytic action of Lactobacillus delbrueckii subsp. bulgaricus CRL 656 reduces antigenic response to bovine β-lactoglobulin. Food Chem. 2011, 127, 487–492. [Google Scholar] [CrossRef]
- El-Ghaish, S.; Rabesona, H.; Choiset, Y.; Sitohy, M.; Haertlé, T.; Chobert, J.-M. Proteolysis by Lactobacillus fermentum IFO3956 isolated from Egyptian milk products decreases immuno-reactivity of αS1-casein. J. Dairy Res. 2011, 78, 203–210. [Google Scholar] [CrossRef]
- Kordesedehi, R.; Taheri-Kafrani, A.; Khorasgani, M.R.; Kazemi, R.; Mutangadura, D.; Haertle, T. Modification of IgE binding to αS1-casein by proteolytic activity of Enterococcus faecium isolated from Iranian camel milk samples. J. Biotechnol. 2018, 276–277, 10–14. [Google Scholar] [CrossRef]
- Wróblewska, B.; Markiewicz, L.H.; Szyc, A.M.; Dietrich, M.A.; Szymkiewicz, A.; Fotschki, J. Lactobacillus casei LcY decreases milk protein immunoreactivity of fermented buttermilk but also contains IgE-reactive proteins. Food Res. Int. 2016, 83, 95–101. [Google Scholar] [CrossRef]
- Uncuoglu, A.; Yologlu, N.; Simsek, I.E.; Uyan, Z.S.; Aydogan, M. Tolerance to baked and fermented cow’s milk in children with IgE-mediated and non-IgE-mediated cow’s milk allergy in patients under two years of age. Allergol. Immunopathol. (Madr.). 2017, 45, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Monaco, S.; Russo, G.; Romano, A.; Liotti, L.; Verga, M.C.; Miceli Sopo, S. Yogurt is tolerated by the majority of children with IgE-mediated cow’s milk allergy. Allergol. Immunopathol. (Madr.) 2019, 47, 322–327. [Google Scholar] [CrossRef]
- Kazemi, R.; Taheri-Kafrani, A.; Motahari, A.; Kordesedehi, R. Allergenicity reduction of bovine milk β-lactoglobulin by proteolytic activity of lactococcus lactis BMC12C and BMC19H isolated from Iranian dairy products. Int. J. Biol. Macromol. 2018, 112, 876–881. [Google Scholar] [CrossRef] [PubMed]
- Ehn, B.-M.; Allmere, T.; Esbjö, E.; Telemo, E.; Bengtsson, U.; Ekstrand, B.O. Modification of IgE binding to β-Lactoglobulin by fermentation and proteolysis of cow’s milk. J. Agric. Food Chem. 2005, 53, 3743–3748. [Google Scholar] [CrossRef]
- Wang, T.; Qin, G.-X.; Sun, Z.-W.; Zhao, Y. Advances of research on glycinin and β-conglycinin: A review of two major soybean allergenic proteins. Crit. Rev. Food Sci. Nutr. 2014, 54, 850–862. [Google Scholar] [CrossRef]
- Gagnon, C.; Poysa, V.; Cober, E.R.; Gleddie, S. Soybean allergens affecting North American patients identified by 2D Gels and mass spectrometry. Food Anal. Methods 2010, 3, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Meinlschmidt, P.; Schweiggert-Weisz, U.; Eisner, P. Soy protein hydrolysates fermentation: Effect of debittering and degradation of major soy allergens. LWT Food Sci. Technol. 2016, 71, 202–212. [Google Scholar] [CrossRef]
- Biscola, V.; de Olmos, A.R.; Choiset, Y.; Rabesona, H.; Garro, M.S.; Mozzi, F.; Chobert, J.-M.; Drouet, M.; Haertlé, T.; Franco, B.D.G.M. Soymilk fermentation by Enterococcus faecalis VB43 leads to reduction in the immunoreactivity of allergenic proteins β-conglycinin (7S) and glycinin (11S). Benef. Microbes 2017, 8, 635–643. [Google Scholar] [CrossRef]
- Yang, A.; Zuo, L.; Cheng, Y.; Wu, Z.; Li, X.; Tong, P.; Chen, H. Degradation of major allergens and allergenicity reduction of soybean meal through solid-state fermentation with microorganisms. Food Funct. 2018, 9, 1899–1909. [Google Scholar] [CrossRef]
- Magishi, N.; Yuikawa, N.; Kobayashi, M.; Taniuchi, S. Degradation and removal of soybean allergen in Japanese soy sauce. Mol. Med. Rep. 2017, 16, 2264–2268. [Google Scholar] [CrossRef]
- Scherf, K.A. Immunoreactive cereal proteins in wheat allergy, non-celiac gluten/wheat sensitivity (NCGS) and celiac disease. Curr. Opin. Food Sci. 2019, 25, 35–41. [Google Scholar] [CrossRef]
- Stefańska, I.; Piasecka-Jóźwiak, K.; Kotyrba, D.; Kolenda, M.; Stecka, K.M. Selection of lactic acid bacteria strains for the hydrolysis of allergenic proteins of wheat flour. J. Sci. Food Agric. 2016, 96, 3897–3905. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; De Angelis, M.; Coda, R.; Gobbetti, M. Use of selected sourdough lactic acid bacteria to hydrolyze wheat and rye proteins responsible for cereal allergy. Eur. Food Res. Technol. 2006, 223, 405–411. [Google Scholar] [CrossRef]
- Kobayashi, M.; Hashimoto, Y.; Taniuchi, S.; Tanabe, S. Degradation of wheat allergen in Japanese soy sauce. Int. J. Mol. Med. 2004. [Google Scholar] [CrossRef]
- Wróblewska, B.; Kaliszewska-Suchodoła, A.; Markiewicz, L.H.; Szyc, A.; Wasilewska, E. Whey prefermented with beneficial microbes modulates immune response and lowers responsiveness to milk allergens in mouse model. J. Funct. Foods 2019, 54, 41–52. [Google Scholar] [CrossRef]
- Park, S.; Bae, J.-H. Fermented food intake is associated with a reduced likelihood of atopic dermatitis in an adult population (Korean National Health and Nutrition Examination Survey 2012–2013). Nutr. Res. 2016, 36, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.K.; Kwon, M.-S.; Lee, J.; Oh, Y.J.; Jang, J.-Y.; Lee, J.-H.; Park, H.W.; Nam, Y.-D.; Seo, M.-J.; Roh, W.; et al. Weissella cibaria WIKIM28 ameliorates atopic dermatitis-like skin lesions by inducing tolerogenic dendritic cells and regulatory T cells in BALB/c mice. Sci. Rep. 2016, 7, 40040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, W.S.; Chen, Y.P.; Chen, M.J. The antiallergic effect of kefir lactobacilli. J. Food Sci. 2010, 75, 244–253. [Google Scholar] [CrossRef]



{kind=link}
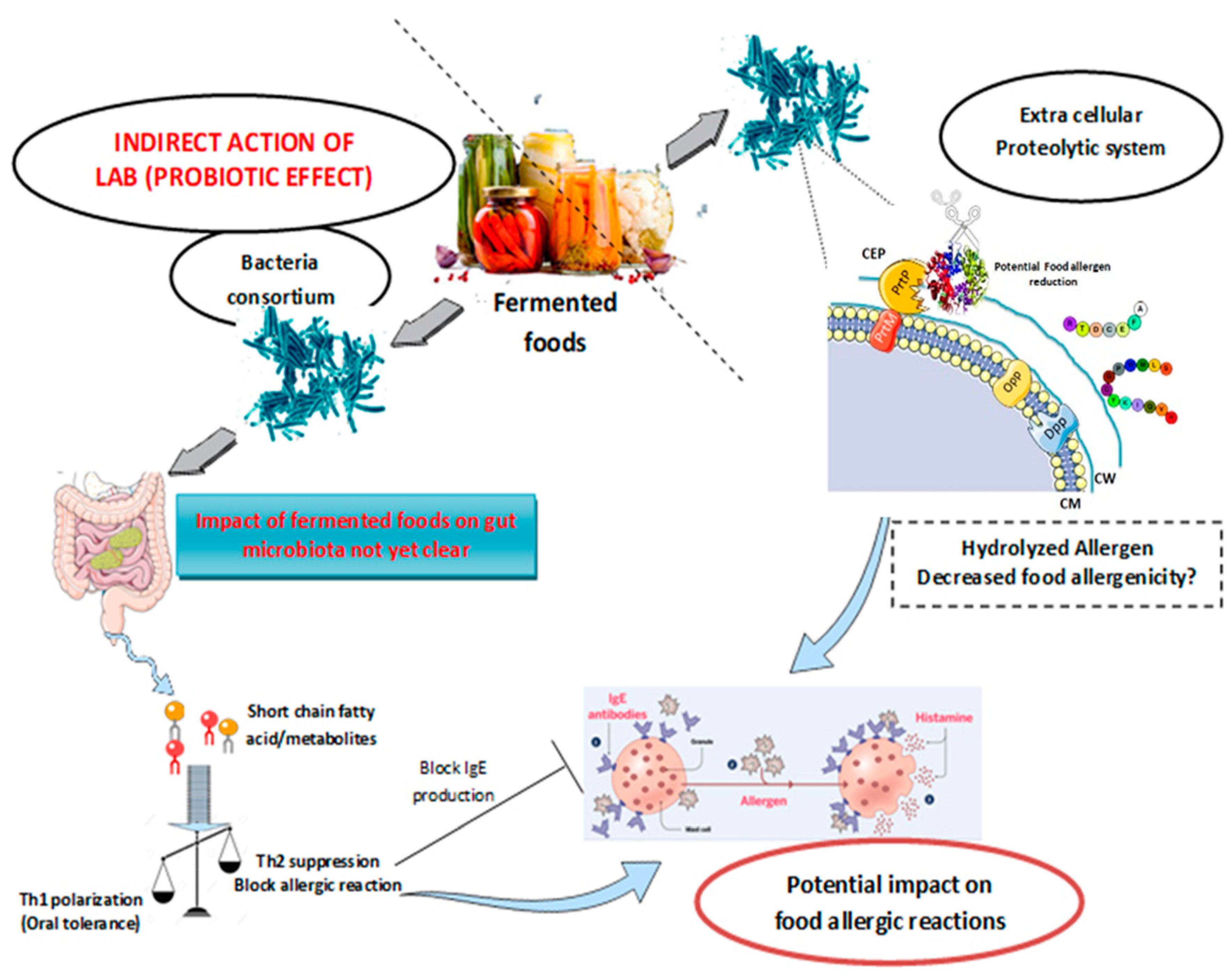{kind=link}
| Cupin | Prolamin | Profilin | PR | |||||
|---|---|---|---|---|---|---|---|---|
| 11S Globulin | 7S, Vicilin, Vicilin-Like | 2S Albumin | Prolamin | LTP, ns LTP | Amylase/Trypsin Inhibitors | |||
| Wheat (Triticum aestivum; T. turgidum ssp.; T. urartu) | Tri a 19, Tri a 20, Tri a 21, Tri a 26, Tri a 36, Tri tp 26, Tri ur 26 | Tri a 14 Tri tu 14 | Tri a 15, Tri a 28, Tri a 29, Tri a 30, Tri a 40, Tri tu 28, Tri ur 28, Tri tu 30 | Tri a 12 | ||||
| Soybean (Glycin max, G. soya) | Gly m 6 Gly s 6 | Gly m 5 Gly s 5 | Gly m 8 | Gly m 3 | Gly m 4 | |||
| Peanut (Arachis hypogea, A. ipaensis) | Ara h 3 | Ara h 1 | Ara h 2 Ara h 6 Ara h 7 | Ara h 9 Ara h 16 Ara h 17 Ara h 9 | Ara h 5 | Ara h 8 | ||
| Sesame (Sesamum indicum) | Ses i 6 Ses i 7 | Ses i 3 | Ses i 1 Ses i 2 | Ses i 8 | ||||
| Brazil Nut (Bertholletia excelsa) | Ber e 2 | Ber e 1 | ||||||
| Hazelnut (Corylus avellana) | Cor a 9 | Cor a 11 | Cor a 14 | Cor a 8 | Cor a 2 | |||
| Lupine (Lupinus angistifolius, L. albus; L. luteus) | Lup an alpha | Lup a 1 | Lup an delta (1) | Lup a 5 | Lup a 4 Lup l 4 | |||
| Buckwheat Fagopyrum esculentum | Fag e 3 Fag e 5 | Fag e 2 | ||||||
| Mustard (Sinapis alba, Brassica juncea, B. campetris, B. rapa) | Sin a 2 | Sin a 1 Bra j 1 Bra r 1 | Sin a 3 Bra r 3 | Sin a 4 Bra j 8 | ||||
| Rapeseed (Brassica napus) | Bra n 1 | Bra n 8 | ||||||
| Pea (Pisum Sativum) | Pis s 1 Pis s 2 | Pis s 3 | Pis s 5 | Pis s 6 | ||||
| Sunflower (Helianthus annuus) | Hel a 2S | Hel a 3 | Hel a 2 | |||||
| Pumpkin (Cucurbita maxima) | Cuc ma 4 | Cuc ma 5 | Cuc ma 2 | |||||
| Poppy seed (Papaver somniferum) | Pap s 2 | Pap s 1 | ||||||
| Lentil (Lens Culinaris) | Len c 1 | Len c 3 | ||||||
| Process | Heat Treatment | Hydrolysis | Other Treatment | |||||
|---|---|---|---|---|---|---|---|---|
| Allergen | Boiling | Roasting | Microwave | Autoclave | Enzymatic | Fermentation | Instant Controlled Pressure Drop | |
| Peanut (Arachis hypogaea) | ↓IgE reactivity [52] | ↑IgE reactivity [53] | ↓IgE reactivity [54] | ↓IgE reactivity after roasting/boiling No results on raw kernel [55] | ↓IgE reactivity at extreme conditions [56] | |||
| Soybean (Glycine max) | ↓IgE reactivity [57] | ↓IgE reactivity: (β Conglycinin) ≈ No effect: (glycinin) | ↓IgE reactivity [58] | ↓IgE reactivity [59] | ||||
| Wheat (Triticum sp.) | ≈ No effect [60] | ↓IgE reactivity [61] | ||||||
| Lupine (Lupinus angistifolius) | ≈ No effect [62] | ↓IgE reactivity [62] | ↓IgE reactivity at extreme conditions [63] | |||||
| (Lupinus albus) | ≈ No effect [62] | |||||||
| Lentil↑ (Lens culinaris) | ≈ No effect [64] | ↓IgE↓ reactivity [64] | Moderate reduction of IgE immunoreactivity [59] | |||||
| Food/Allergen | Strain | Conditions | In Vitro Analysis | In Vivo Tests | Reference |
|---|---|---|---|---|---|
| Na-caseinate and αS1-casein | Enterococcus faecium | Non proliferative system | Decrease of IgE binding | [102] | |
| Sweet buttermilk | L. casei LcY | Fermented system | Decrease of IgE binding towards all milk allergens. Reactivity to new proteins isolated from cell wall membrane. | [103] | |
| β-lactoglobulin | Lactococcus lactis | Non proliferative system | Decrease of IgE binding | [106] | |
| β-lactoglobulin | Lactobacillus delbrueckii subsp.bulgaricus CRL 656 | Non proliferative system | Decrease of IgE binding | [100] | |
| Milk | Lactobacillus helveticus | Fermented system | Unmodified IgE binding toward β—lactoglobulin | [107] | |
| Fermented milk | N.S. | Decrease PT reactivity, decrease IgE β—lactoglobulin and lactalbumin. | Yogurt tolerated by subjects tolerating heated milk | [104] | |
| Fermented milk (Yogurt) | N.S. | Succeed of oral food challenge with fermented food. | [105] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Mecherfi, K.-E.; Todorov, S.D.; Cavalcanti de Albuquerque, M.A.; Denery-Papini, S.; Lupi, R.; Haertlé, T.; Dora Gombossy de Melo Franco, B.; Larré, C. Allergenicity of Fermented Foods: Emphasis on Seeds Protein-Based Products. Foods 2020, 9, 792. https://doi.org/10.3390/foods9060792
El Mecherfi K-E, Todorov SD, Cavalcanti de Albuquerque MA, Denery-Papini S, Lupi R, Haertlé T, Dora Gombossy de Melo Franco B, Larré C. Allergenicity of Fermented Foods: Emphasis on Seeds Protein-Based Products. Foods. 2020; 9(6):792. https://doi.org/10.3390/foods9060792
Chicago/Turabian StyleEl Mecherfi, Kamel-Eddine, Svetoslav Dimitrov Todorov, Marcela Albuquerque Cavalcanti de Albuquerque, Sandra Denery-Papini, Roberta Lupi, Thomas Haertlé, Bernadette Dora Gombossy de Melo Franco, and Colette Larré. 2020. "Allergenicity of Fermented Foods: Emphasis on Seeds Protein-Based Products" Foods 9, no. 6: 792. https://doi.org/10.3390/foods9060792
APA StyleEl Mecherfi, K. -E., Todorov, S. D., Cavalcanti de Albuquerque, M. A., Denery-Papini, S., Lupi, R., Haertlé, T., Dora Gombossy de Melo Franco, B., & Larré, C. (2020). Allergenicity of Fermented Foods: Emphasis on Seeds Protein-Based Products. Foods, 9(6), 792. https://doi.org/10.3390/foods9060792