The Antioxidant and Anti-Inflammatory Properties of Rice Bran Phenolic Extracts

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Rice Bran Extract Preparation
2.3. Cell Culture Conditions
2.4. Cytotoxicity Assessment
2.5. Antioxidant Effect of RB Phenolic Extract
2.5.1. Malondialdehyde Determination
2.5.2. Intracellular Reactive Oxygen Species Generation
2.6. Anti-Inflammatory Effect of RB Phenolic Extract
2.6.1. Nitric Oxide Determination
2.6.2. Inflammatory Cytokine Determination
2.7. Statistical Analysis
3. Results
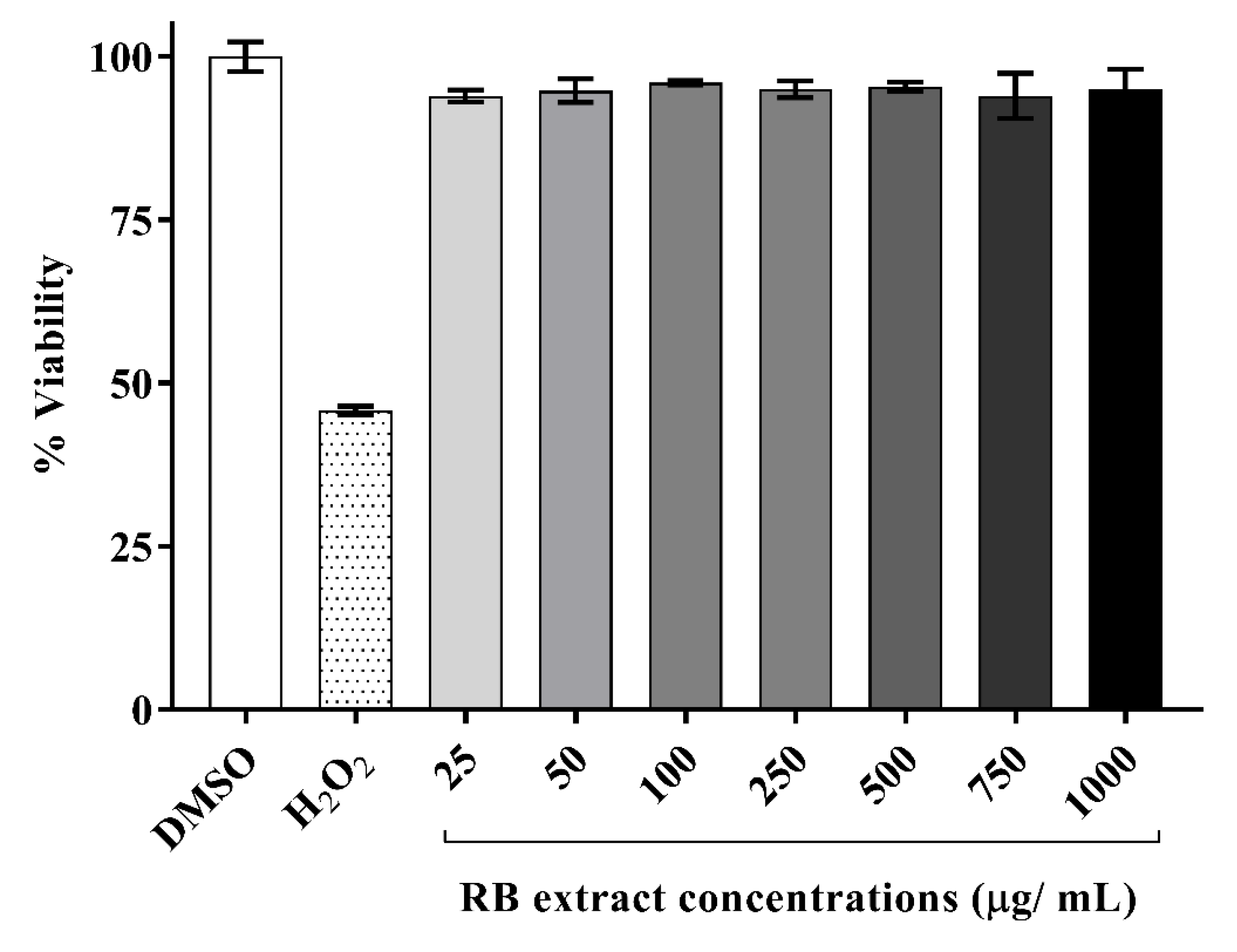
3.1. Cytotoxicity of RB Phenolic Extracts on RAW264.7 Cells
3.2. Antioxidant Properties of RB Phenolic Extract
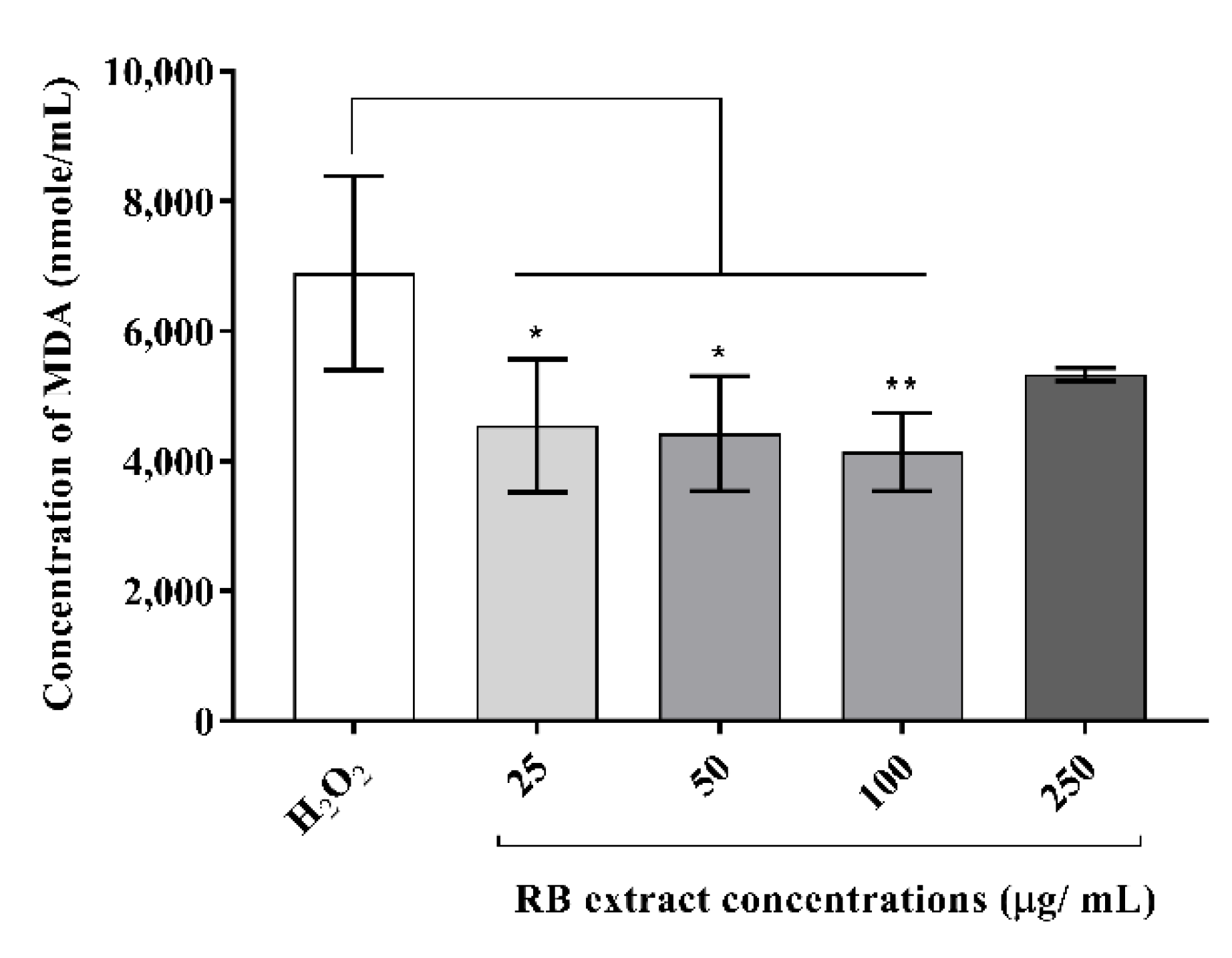
3.2.1. Malondialdehyde Concentration
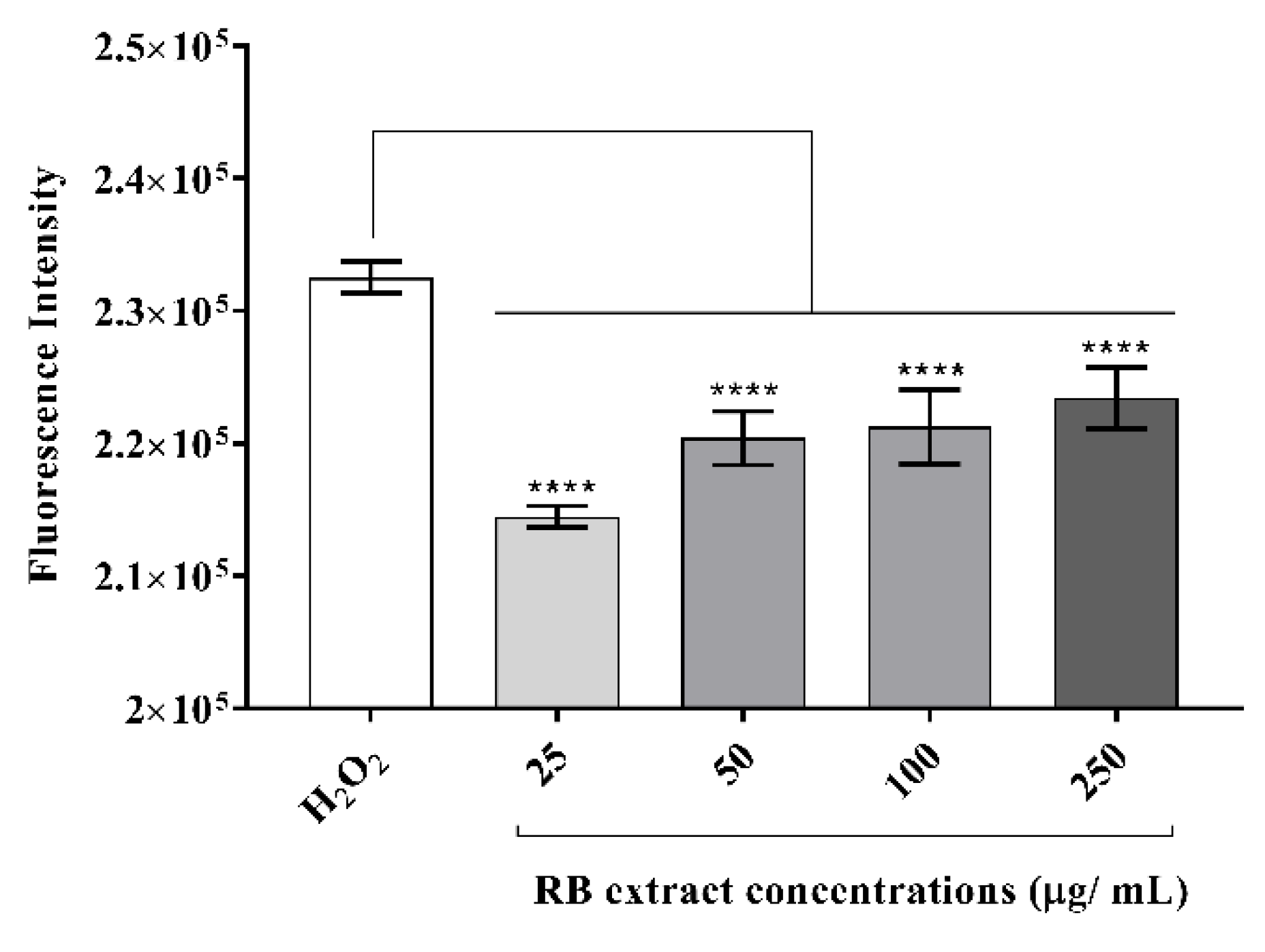
3.2.2. Intracellular Reactive Oxygen Species Generation
3.3. Anti-Inflammatory Effect of RB Phenolic Extract
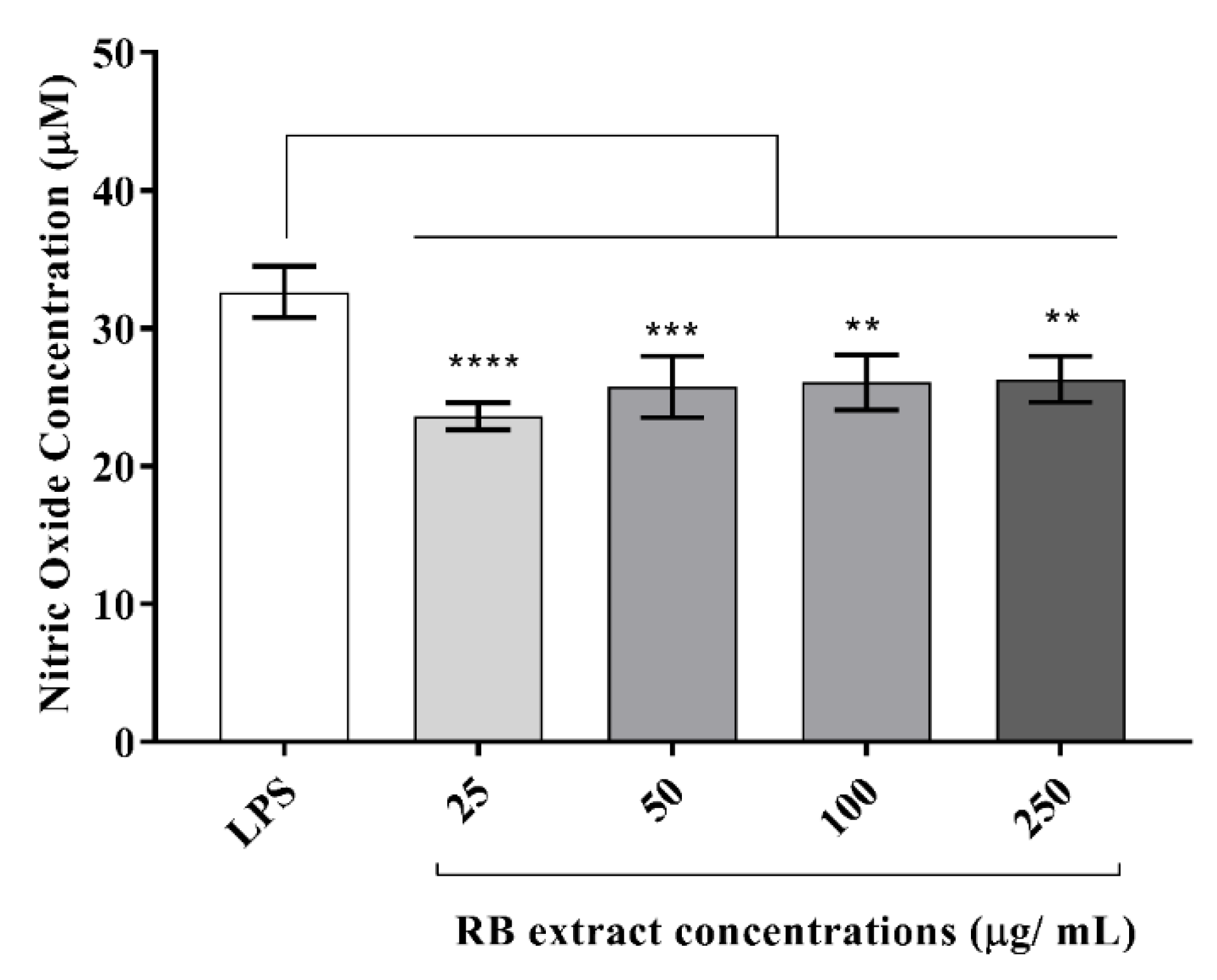
3.3.1. Nitric Oxide Determination
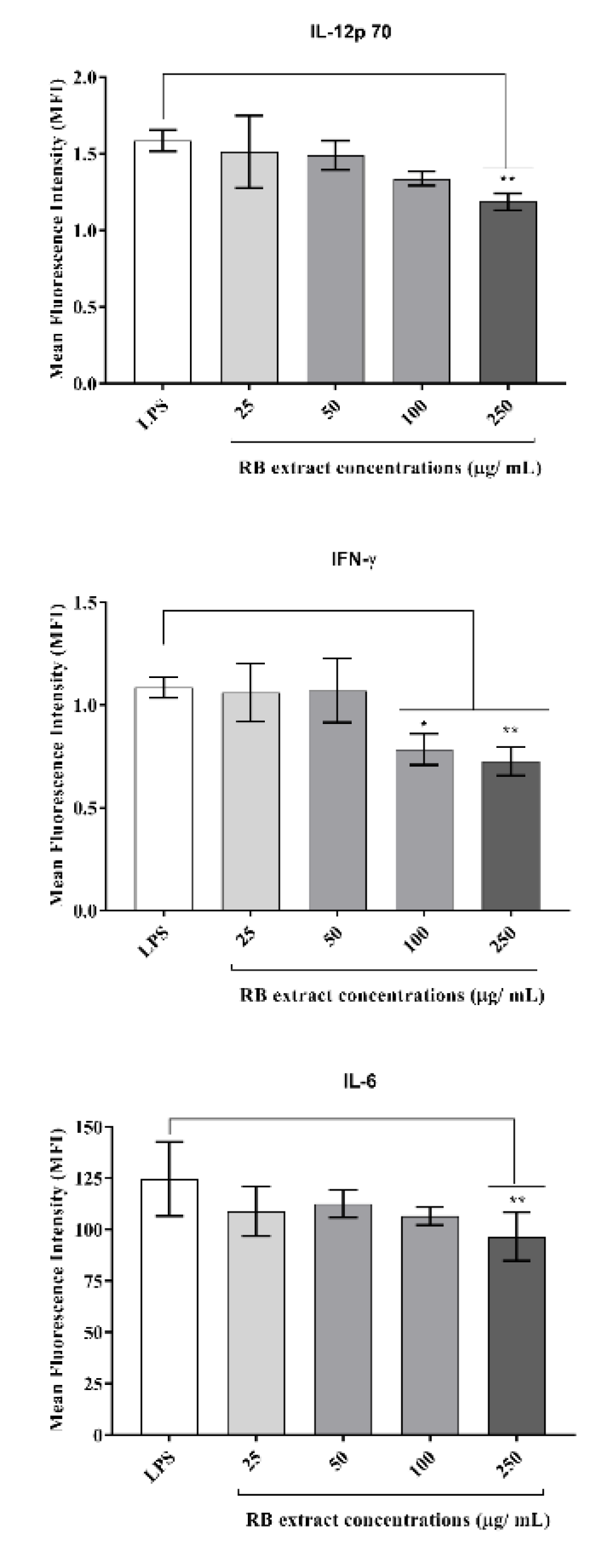
3.3.2. Inflammatory Cytokine Determination
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| COX | Cyclooxygenase |
| DMSO | Dimethyl sulfoxide |
| DMEM | Dulbecco’s modified Eagle’s medium |
| H2O2 | Hydrogen peroxide |
| IFN-γ | Interferon-γ |
| IL-10 | Interleukin-10 |
| IL-12p70 | Interleukin-12, p70 |
| IL-6 | Interleukin-6 |
| LPS | Lipopolysaccharide |
| LOX | Lipoxygenase |
| MDA | Malondialdehyde |
| MAPK | Mitogen-activated protein kinase |
| MCP-1 | Monocyte chemoattractant protein 1 |
| NOS | Nitric oxide synthase |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| RB | Rice bran |
| STAT3 | Signal transducer and activator of transcription 3 |
| SD | Standard deviation |
| TBA | Thiobarbituric acid |
| TNF-α | Tumor necrosis factor-α |
References
- He, F.; Zuo, L. Redox roles of reactive oxygen species in cardiovascular diseases. Int. J. Mol. Sci. 2015, 16, 27770–27780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saji, N.; Francis, N.; Schwarz, L.J.; Blanchard, C.L.; Santhakumar, A.B. Rice bran derived bioactive compounds modulate risk factors of cardiovascular disease and type 2 diabetes mellitus: An updated review. Nutrients 2019, 11, 2736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korns, D.R.; Frasch, S.C.; Fernandez-Boyanapalli, R.; Henson, P.M.; Bratton, D.L. Modulation of macrophage efferocytosis in inflammation. Front. Immunol. 2011, 2, 57. [Google Scholar] [CrossRef] [Green Version]
- Ellulu, M.S. Obesity, cardiovascular disease, and role of vitamin C on inflammation: A review of facts and underlying mechanisms. Inflammopharmacology 2017, 25, 313–328. [Google Scholar] [CrossRef] [PubMed]
- Poffenberger, M.C.; Straka, N.; El Warry, N.; Fang, D.; Shanina, I.; Horwitz, M.S. Lack of IL-6 during coxsackievirus infection heightens the early immune response resulting in increased severity of chronic autoimmune myocarditis. PLoS ONE 2009, 4, e6207. [Google Scholar] [CrossRef] [PubMed]
- Slatter, D.; Bolton, C.; Bailey, A. The importance of lipid-derived malondialdehyde in diabetes mellitus. Diabetologia 2000, 43, 550–557. [Google Scholar] [CrossRef]
- Porter, N.A.; Caldwell, S.E.; Mills, K.A. Mechanisms of free radical oxidation of unsaturated lipids. Lipids 1995, 30, 277–290. [Google Scholar] [CrossRef]
- Uchida, K. Role of reactive aldehyde in cardiovascular diseases. Free Radic. Boil. Med. 2000, 28, 1685–1696. [Google Scholar] [CrossRef]
- Naseem, K.M. The role of nitric oxide in cardiovascular diseases. Mol. Asp. Med. 2005, 26, 33–65. [Google Scholar] [CrossRef]
- Abeywardena, M.Y.; Leifert, W.R.; Warnes, K.E.; Varghese, J.N.; Head, R.J. Cardiovascular Boilogy of interleukin-6. Curr. Pharm. Des. 2009, 15, 1809–1821. [Google Scholar] [CrossRef] [Green Version]
- Niu, J.; Kolattukudy, P.E. Role of MCP-1 in cardiovascular disease: Molecular mechanisms and clinical implications. Clin. Sci. 2009, 117, 95–109. [Google Scholar] [CrossRef] [Green Version]
- Girndt, M.; Köhler, H. Interleukin-10 (IL-10): An update on its relevance for cardiovascular risk. Nephrol. Dial. Transplant. 2003, 18, 1976–1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrari, R. The role of TNF in cardiovascular disease. Pharmacol. Res. 1999, 40, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Huo, X.; Zhang, Y.; Wang, Q.; Zhang, Y.; Xu, X. Cardiovascular endothelial inflammation by chronic coexposure to lead (Pb) and polycyclic aromatic hydrocarbons from preschool children in an e-waste recycling area. Environ. Pollut. 2019, 246, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.M.; Cummings, E.; Pantos, C.; Singh, J. Protein kinase C and cardiac dysfunction: A review. Heart Fail. Rev. 2017, 22, 843–859. [Google Scholar] [CrossRef] [Green Version]
- Voloshyna, I.; Littlefield, M.J.; Reiss, A.B. Atherosclerosis and interferon-γ: New insights and therapeutic targets. Trends Cardiovasc. Med. 2014, 24, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Shattat, G.F. A review article on hyperlipidemia: Types, treatments and new drug targets. Biomed. Pharmacol. J. 2015, 7, 399–409. [Google Scholar] [CrossRef]
- Callcott, E.T.; Blanchard, C.L.; Oli, P.; Santhakumar, A.B. Pigmented Rice-Derived Phenolic Compounds Reduce Biomarkers of Oxidative Stress and Inflammation in Human Umbilical Vein Endothelial Cells. Mol. Nutr. Food Res. 2018, 62, 1800840. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxidative Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [Green Version]
- Saji, N.; Schwarz, L.J.; Santhakumar, A.B.; Blanchard, C.L. Stabilization treatment of rice bran alters phenolic content and antioxidant activity. Cereal Chem. 2020, 97, 281–292. [Google Scholar] [CrossRef]
- Saji, N.; Francis, N.; Blanchard, C.L.; Schwarz, L.J.; Santhakumar, A.B. Rice Bran Phenolic Compounds Regulate Genes Associated with Antioxidant and Anti-Inflammatory Activity in Human Umbilical Vein Endothelial Cells with Induced Oxidative Stress. Int. J. Mol. Sci. 2019, 20, 4715. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Bai, D.; Wei, Z.; Zhang, Y.; Huang, Y.; Deng, H.; Huang, X. Curcumin attenuates oxidative stress in RAW264. 7 cells by increasing the activity of antioxidant enzymes and activating the Nrf2-Keap1 pathway. PLoS ONE 2019, 14, e0216711. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.-S.; Hsu, W.-H.; Huang, J.-J.; Wu, S.-C. Antioxidant and anti-inflammatory effects of pigeon pea (Cajanus cajan L.) extracts on hydrogen peroxide-and lipopolysaccharide-treated RAW264. 7 macrophages. Food Funct. 2012, 3, 1294–1301. [Google Scholar] [CrossRef]
- Hwang, K.-A.; Hwang, Y.-J.; Song, J. Antioxidant activities and oxidative stress inhibitory effects of ethanol extracts from Cornus officinalis on raw 264.7 cells. BMC Complementary Altern. Med. 2016, 16, 196. [Google Scholar]
- Zhang, Q.; Luna-Vital, D.; de Mejia, E.G. Anthocyanins from colored maize ameliorated the inflammatory paracrine interplay between macrophages and adipocytes through regulation of NF-κB and JNK-dependent MAPK pathways. J. Funct. Foods 2019, 54, 175–186. [Google Scholar] [CrossRef]
- Yu, H.-S.; Lee, N.-K.; Choi, A.-J.; Choe, J.-S.; Bae, C.H.; Paik, H.-D. Anti-inflammatory potential of probiotic strain Weissella cibaria JW15 isolated from Kimchi through regulation of NF-κB and MAPKs pathways in LPS-induced RAW 264. 7 Cells. J. Microboil. Biotechnol. 2019, 29, 1022–1032. [Google Scholar] [CrossRef]
- Ben Lagha, A.; Andrian, E.; Grenier, D. Resveratrol attenuates the pathogenic and inflammatory properties of Porphyromonas gingivalis. Mol. Oral Microboil. 2019, 34, 118–130. [Google Scholar] [CrossRef]
- Taira, J.; Nanbu, H.; Ueda, K. Nitric oxide-scavenging compounds in Agrimonia pilosa Ledeb on LPS-induced RAW264. 7 macrophages. Food Chem. 2009, 115, 1221–1227. [Google Scholar] [CrossRef]
- Jeong, J.B.; Jeong, H.J. Rheosmin, a naturally occurring phenolic compound inhibits LPS-induced iNOS and COX-2 expression in RAW264. 7 cells by blocking NF-κB activation pathway. Food Chem. Toxicol. 2010, 48, 2148–2153. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Cao, W.; Zhu, Y. Immunoregulatory Functions of the IL-12 Family of Cytokines in Antiviral Systems. Viruses 2019, 11, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Wang, Q. Factors determining the formation and release of bioactive IL-12: Regulatory mechanisms for IL-12p70 synthesis and inhibition. Biochem. Biophys. Res. Commun. 2008, 372, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Harizi, H.; Gualde, N. The impact of eicosanoids on the crosstalk between innate and adaptive immunity: The key roles of dendritic cells. Tissue Antigens 2005, 65, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Tramsen, L.; Lehrnbecher, T. Natural killer cells in antifungal immunity. Front. Immunol. 2017, 8, 1623. [Google Scholar] [CrossRef] [PubMed]
- Greenhill, C.J.; Rose-John, S.; Lissilaa, R.; Ferlin, W.; Ernst, M.; Hertzog, P.J.; Mansell, A.; Jenkins, B.J. IL-6 trans-signaling modulates TLR4-dependent inflammatory responses via STAT3. J. Immunol. 2011, 186, 1199–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holst, B.; Williamson, G. Nutrients and phytochemicals: From bioavailability to bioefficacy beyond antioxidants. Curr. Opin. Biotechnol. 2008, 19, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P. Hormesis defined. Ageing Res. Rev. 2008, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saji, N.; Francis, N.; Schwarz, L.J.; Blanchard, C.L.; Santhakumar, A.B. The Antioxidant and Anti-Inflammatory Properties of Rice Bran Phenolic Extracts. Foods 2020, 9, 829. https://doi.org/10.3390/foods9060829
Saji N, Francis N, Schwarz LJ, Blanchard CL, Santhakumar AB. The Antioxidant and Anti-Inflammatory Properties of Rice Bran Phenolic Extracts. Foods. 2020; 9(6):829. https://doi.org/10.3390/foods9060829
Chicago/Turabian StyleSaji, Nancy, Nidhish Francis, Lachlan J. Schwarz, Christopher L. Blanchard, and Abishek B. Santhakumar. 2020. "The Antioxidant and Anti-Inflammatory Properties of Rice Bran Phenolic Extracts" Foods 9, no. 6: 829. https://doi.org/10.3390/foods9060829
APA StyleSaji, N., Francis, N., Schwarz, L. J., Blanchard, C. L., & Santhakumar, A. B. (2020). The Antioxidant and Anti-Inflammatory Properties of Rice Bran Phenolic Extracts. Foods, 9(6), 829. https://doi.org/10.3390/foods9060829