Metabolite Profiling and Chemometric Study for the Discrimination Analyses of Geographic Origin of Perilla (Perilla frutescens) and Sesame (Sesamum indicum) Seeds

 , and
, and
Abstract
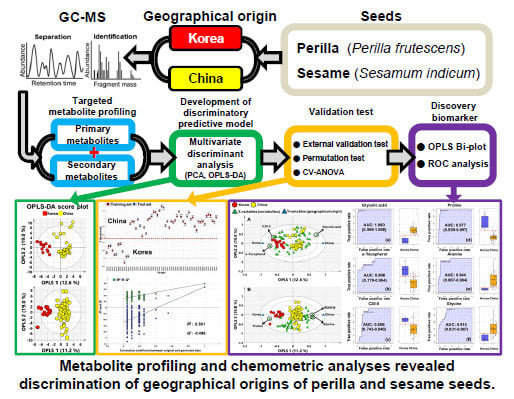
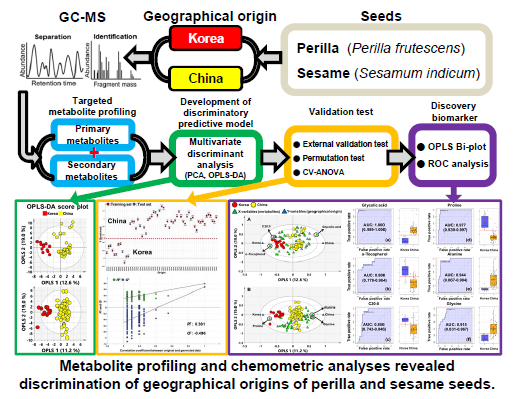
:
1. Introduction
2. Materials and Methods
2.1. Sample and Chemicals
2.2. Extraction and Analysis of Hydrophilic Compounds
2.3. Extraction and Analysis of Lipophilic Compounds
2.4. Statistical Analysis
3. Results
3.1. Metabolite Profiling of Perilla and Sesame Seeds
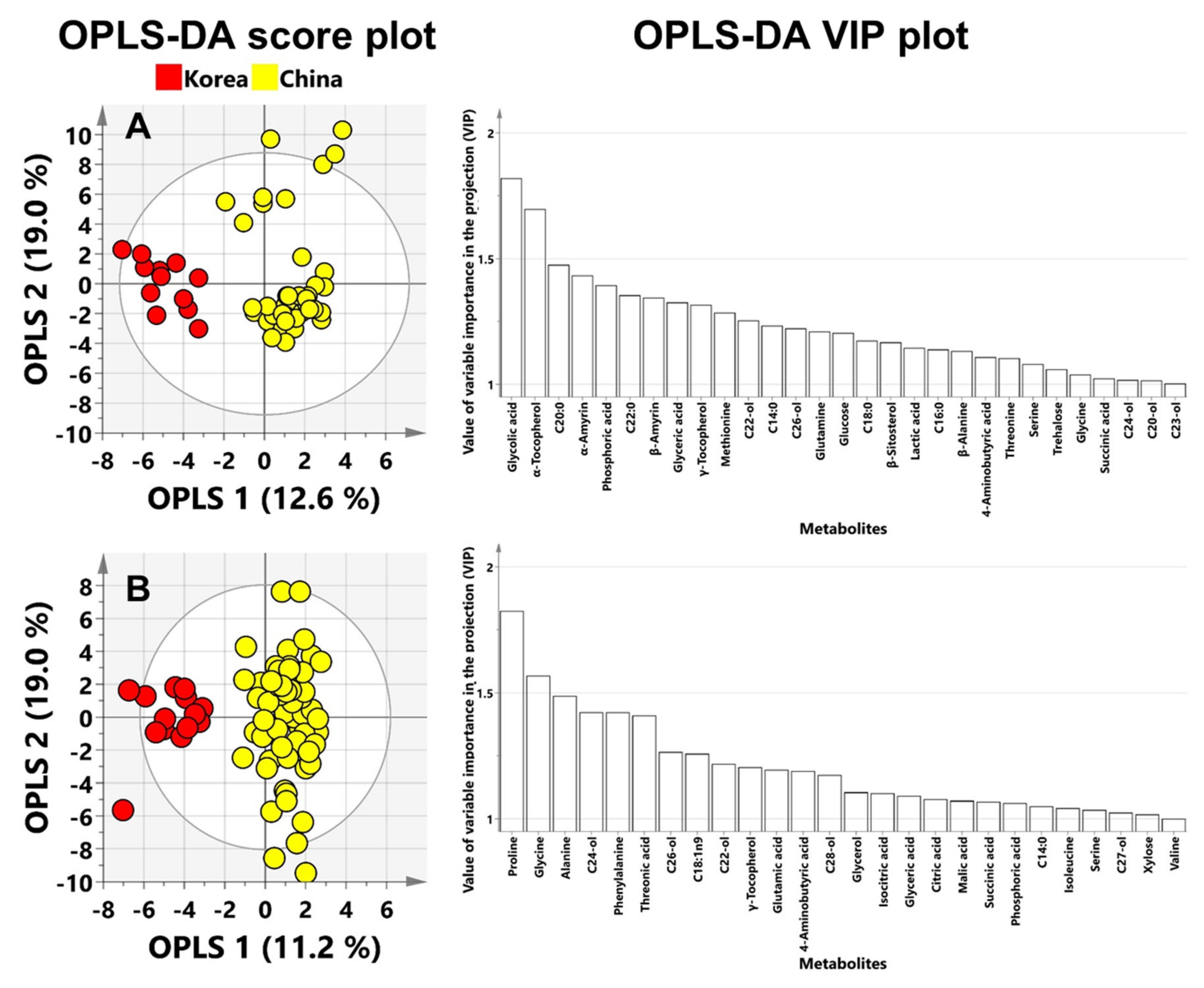
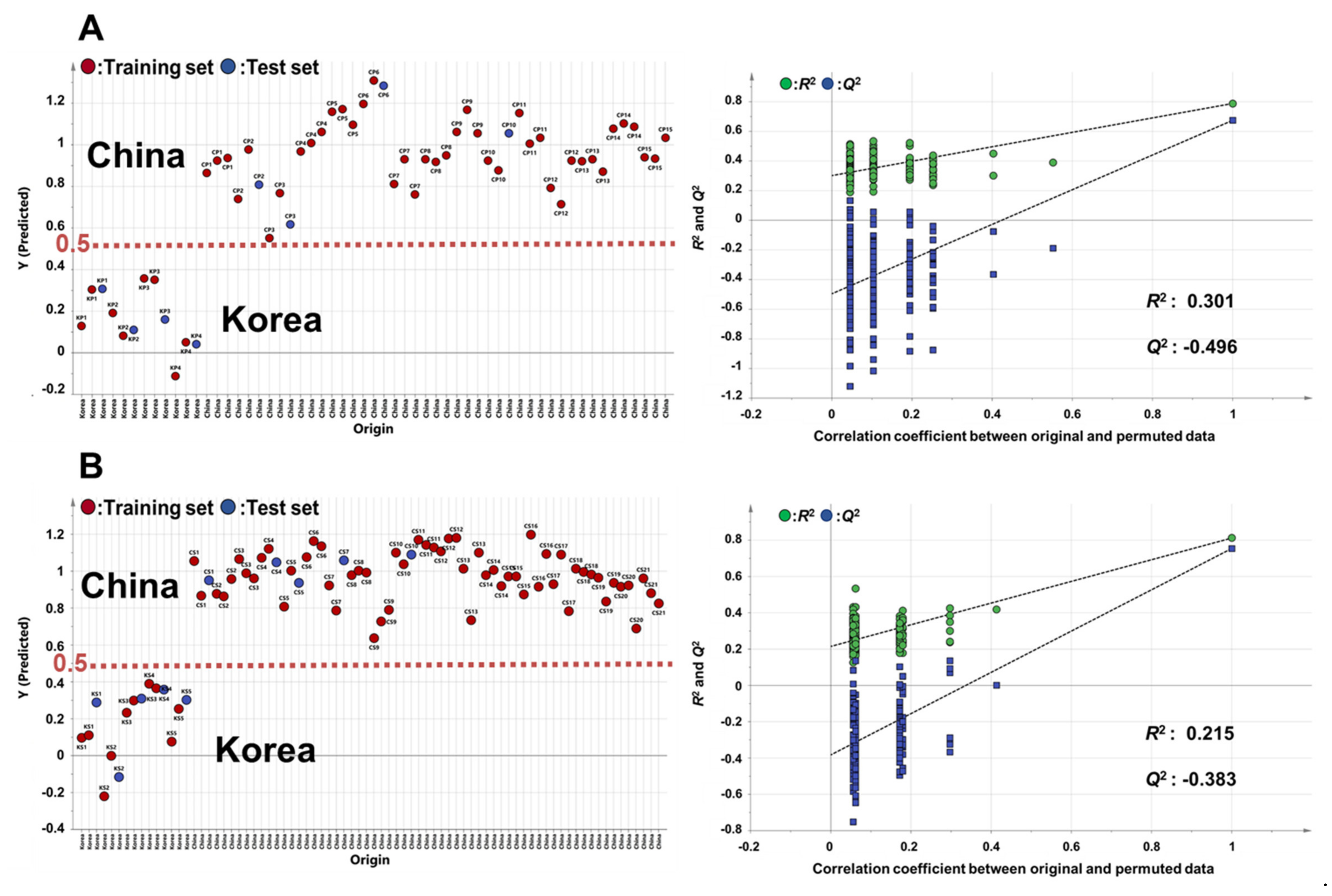
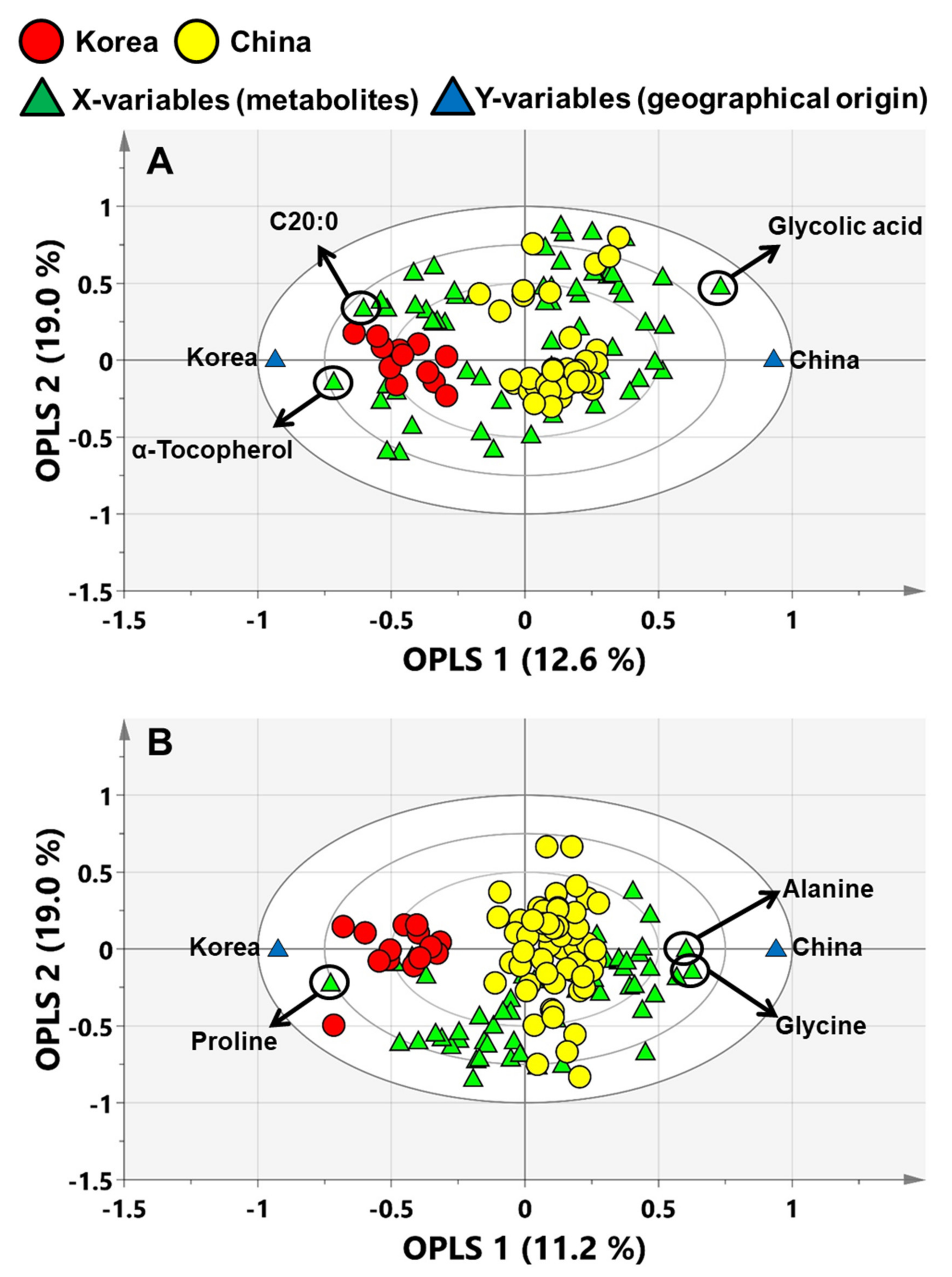
3.2. PCA and OPLS-DA for Geographic Discrimination of Perilla and Sesame Seeds
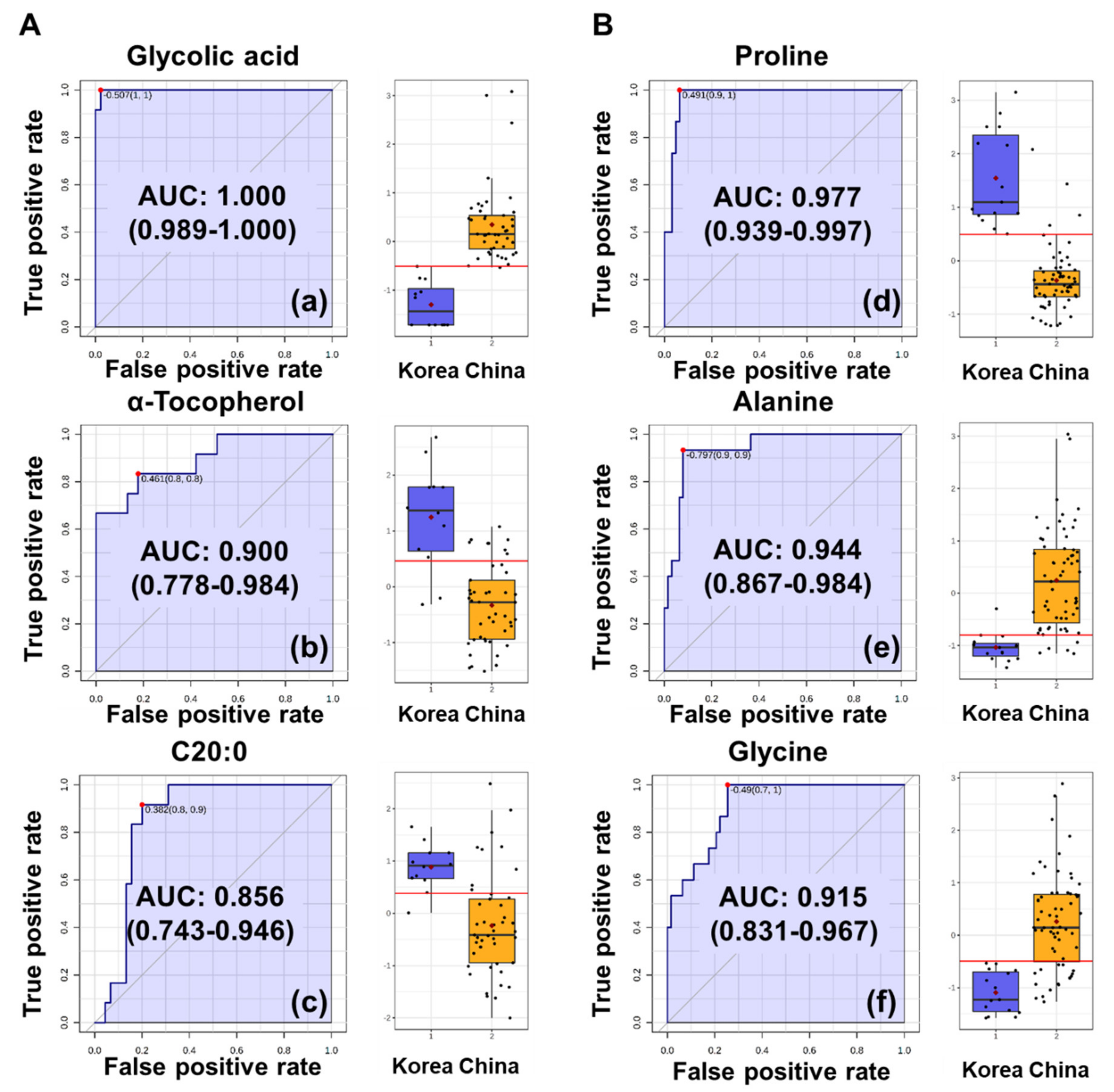
3.3. Potential Biomarkers for the Discrimination of Perilla and Sesame Seeds Based on Their Geographic Origins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kim, J.K.; Park, S.-Y.; Na, J.-K.; Seong, E.S.; Yu, C.Y. Metabolite profiling based on lipophilic compounds for quality assessment of perilla (Perilla frutescens) cultivars. J. Agric. Food Chem. 2012, 60, 2257–2263. [Google Scholar] [CrossRef]
- Shen, J.; Luo, F.; Lin, Q. Policosanol: Extraction and biological functions. J. Funct. Foods 2019, 57, 351–360. [Google Scholar] [CrossRef]
- Lee, K.-R.; Kim, K.-H.; Kim, J.B.; Hong, S.-B.; Jeon, I.; Kim, H.U.; Lee, M.H.; Kim, J.K. High accumulation of γ-linolenic acid and Stearidonic acid in transgenic perilla (Perilla frutescens var. frutescens) seeds. BMC Plant Biol. 2019, 19, 120. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Kim, E.; Shin, B.K.; Seo, J.-A.; Kim, Y.-S.; Lee, D.Y.; Choi, H.-K. NMR-Based metabolic profiling discriminates the geographical origin of raw sesame seeds. Food Control 2020, 112, 107–113. [Google Scholar] [CrossRef]
- Pathak, N.; Rai, A.; Kumari, R.; Bhat, K. Value addition in sesame: A perspective on bioactive components for enhancing utility and profitability. Pharmacogn. Rev. 2014, 8, 147–155. [Google Scholar]
- Jeon, H.; Kim, I.H.; Lee, C.; Choi, H.D.; Kim, B.H.; Akoh, C.C. Discrimination of origin of sesame oils using fatty acid and lignan profiles in combination with canonical discriminant analysis. J. Am. Oil Chem. Soc. 2013, 90, 337–347. [Google Scholar] [CrossRef]
- Williamson, K.S.; Morris, J.B.; Pye, Q.N.; Kamat, C.D.; Hensley, K. A survey of sesamin and composition of tocopherol variability from seeds of eleven diverse sesame (Sesamum indicum L.) genotypes using HPLC-PAD-ECD. Phytochem. Anal. 2008, 19, 311–322. [Google Scholar] [CrossRef]
- Castro-Puyana, M.; Herrero, M. Metabolomics approaches based on mass spectrometry for food safety, quality and traceability. TrAC Trends Anal. Chem. 2013, 52, 74–87. [Google Scholar] [CrossRef]
- Castro-Puyana, M.; Pérez-Míguez, R.; Montero, L.; Herrero, M. Reprint of: Application of mass spectrometry-based metabolomics approaches for food safety, quality and traceability. TrAC Trends Anal. Chem. 2017, 96, 62–78. [Google Scholar] [CrossRef]
- Salvo, A.; Rotondo, A.; Mangano, V.; Grimaldi, M.; Stillitano, I.; D’Ursi, A.M.; Dugo, G.; Rastrelli, L. High-Resolution magic angle spinning nuclear magnetic resonance (HR-MAS-NMR) as quick and direct insight of almonds. Nat. Prod. Res. 2017, 34, 71–77. [Google Scholar] [CrossRef]
- Kim, T.J.; Park, J.G.; Ahn, S.K.; Kim, K.W.; Choi, J.; Kim, H.Y.; Ha, S.-H.; Seo, W.D.; Kim, J.K. Discrimination of adzuki bean (Vigna angularis) geographical origin by targeted and non-targeted metabolite profiling with gas chromatography time-of-flight mass spectrometry. Metabolites 2020, 10, 112. [Google Scholar] [CrossRef] [Green Version]
- Park, C.H.; Yeo, H.J.; Park, S.-Y.; Kim, J.K.; Park, S.U. Comparative phytochemical analyses and metabolic profiling of different phenotypes of Chinese cabbage (Brassica Rapa ssp. Pekinensis). Foods 2019, 8, 587. [Google Scholar] [CrossRef] [Green Version]
- Putri, S.P.; Irifune, T.; Fukusaki, E. GC/MS based metabolite profiling of Indonesian specialty coffee from different species and geographical origin. Metabolomics 2019, 15, 126. [Google Scholar] [CrossRef]
- Mo, C.; Lim, J.; Kwon, S.W.; Lim, D.K.; Kim, M.S.; Kim, G.; Kang, J.; Kwon, K.-D.; Cho, B.-K. Hyperspectral imaging and partial least square discriminant analysis for geographical origin discrimination of white rice. J. Biosyst. Eng. 2017, 42, 293–300. [Google Scholar]
- Luykx, D.M.; Van Ruth, S.M. An overview of analytical methods for determining the geographical origin of food products. Food Chem. 2008, 107, 897–911. [Google Scholar] [CrossRef]
- Vadalà, R.; Mottese, A.F.; Bua, G.D.; Salvo, A.; Mallamace, D.; Corsaro, C.; Vasi, S.; Giofrè, S.V.; Alfa, M.; Cicero, N.; et al. Statistical analysis of mineral concentration for the geographic identification of garlic samples from Sicily (Italy), Tunisia and Spain. Foods 2016, 5, 20. [Google Scholar] [CrossRef] [Green Version]
- Dossa, K.; Wei, X.; Zhang, Y.; Fonceka, D.; Yang, W.; Diouf, D.; Liao, B.; Cissé, N.; Zhang, X. Analysis of genetic diversity and population structure of sesame accessions from Africa and Asia as major centers of its cultivation. Genes 2016, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Ghimire, B.K.; Yoo, J.H.; Yu, C.Y.; Chung, I.-M. GC–MS analysis of volatile compounds of Perilla frutescens Britton var. Japonica accessions: Morphological and seasonal variability. Asian Pac. J. Trop. Med. 2017, 10, 643–651. [Google Scholar]
- Korea Statistical Information Service. Available online: http://kosis.kr/statHtml/statHtml.do?orgId=101&tblId=DT_1ET0293&conn_path=I2 (accessed on 26 May 2020).
- Korea Agricultural Trade Information. Available online: http://www.kati.net/statistics/regionalPerformanceByProduct.do (accessed on 26 May 2020).
- Horacek, M.; Hansel-Hohl, K.; Burg, K.; Soja, G.; Okello-Anyanga, W.; Fluch, S. Control of origin of sesame oil from various countries by stable isotope analysis and DNA based markers—A pilot study. PLoS ONE 2015, 10, e0123020. [Google Scholar] [CrossRef]
- Choi, Y.H.; Hong, C.K.; Kim, M.; Jung, S.O.; Park, J.; Oh, Y.H.; Kwon, J.-H. Multivariate analysis to discriminate the origin of sesame seeds by multi-element analysis inductively coupled plasma-mass spectrometry. Food Sci. Biotechnol. 2017, 26, 375–379. [Google Scholar] [CrossRef]
- Sa, K.J.; Park, D.H.; Lim, S.E.; Ma, S.J.; Lee, J.K. Geographic variation of morphological characters among cultivated types of perilla crop and their weedy types in Korea and China. Plant Breed. Biotechnol. 2019, 7, 313–325. [Google Scholar] [CrossRef]
- Bang, H.J.; Kim, C.T.; Kim, B.H. Liquid and gas chromatographic analyses of triacylglycerols for Asian sesame oil traceability. Eur. J. Lipid Sci. Technol. 2014, 116, 1354–1362. [Google Scholar] [CrossRef]
- Jin, G.; Kim, J.; Lee, Y.; Kim, J.; Akoh, C.C.; Chun, H.S.; Ahn, S.; Kim, B.H. A nuclear magnetic resonance spectroscopy approach to discriminate the geographic origin of roasted Asian sesame oils. J. Oleo Sci. 2017, 66, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.J.; Choi, J.; Kim, K.W.; Ahn, S.K.; Ha, S.H.; Choi, Y.; Park, N.I.; Kim, J.K. Metabolite profiling of peppers of various colors reveals relationships between tocopherol, carotenoid, and phytosterol content. J. Food Sci. 2017, 82, 2885–2893. [Google Scholar] [CrossRef]
- Kim, T.J.; Lee, K.B.; Baek, S.-A.; Choi, J.; Ha, S.-H.; Lim, S.-H.; Park, S.-Y.; Yeo, Y.; Park, S.U.; Kim, J.K. Determination of lipophilic metabolites for species discrimination and quality assessment of nine leafy vegetables. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 909–918. [Google Scholar] [CrossRef]
- Park, P.; Goins, R. In situ preparation of fatty acid methyl esters for analysis of fatty acid composition in foods. J. Food Sci. 1994, 59, 1262–1266. [Google Scholar] [CrossRef]
- Lee, C.M.; Trevino, B.; Chaiyawat, M. A simple and rapid solvent extractionmethod for determining total lipids in fish tissue. J. AOAC Int. 1996, 79, 487–492. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, L.; Byrne, T.; Johansson, E.; Trygg, J.; Vikström, C. Multi- and Megavariate Data Analysis Basic Principles and Applications; Umetrics Academy: Umea, Sweden, 2013; Volume 1. [Google Scholar]
- Adhikari, P.; Hwang, K.T.; Park, J.N.; Kim, C.K. Policosanol content and composition in perilla seeds. J. Agric. Food Chem. 2006, 54, 5359–5362. [Google Scholar] [CrossRef]
- Jung, D.M.; Lee, M.J.; Yoon, S.H.; Jung, M.Y. A gas chromatography-tandem quadrupole mass spectrometric analysis of policosanols in commercial vegetable oils. J. Food Sci. 2011, 76, C891–C899. [Google Scholar] [CrossRef]
- Skov, T.; Honoré, A.H.; Jensen, H.M.; Næs, T.; Engelsen, S.B. Chemometrics in foodomics: Handling data structures from multiple analytical platforms. TrAC Trends Anal. Chem. 2014, 60, 71–79. [Google Scholar] [CrossRef]
- South, P.F.; Walker, B.J.; Cavanagh, A.P.; Rolland, V.; Badger, M.; Ort, D.R. Bile acid sodium symporter BASS6 can transport glycolate and is involved in photorespiratory metabolism in Arabidopsis thaliana. Plant Cell 2017, 29, 808–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | X Variables Number | Scaling Method | R2X | R2Y | Q2 |
|---|---|---|---|---|---|
| Perilla | 57 | UV | 0.316 | 0.822 | 0.761 |
| Par | 0.473 | 0.575 | 0.480 | ||
| Sesame | 78 | UV | 0.303 | 0.844 | 0.799 |
| Par | 0.526 | 0.744 | 0.715 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, T.J.; Park, J.G.; Kim, H.Y.; Ha, S.-H.; Lee, B.; Park, S.U.; Seo, W.D.; Kim, J.K. Metabolite Profiling and Chemometric Study for the Discrimination Analyses of Geographic Origin of Perilla (Perilla frutescens) and Sesame (Sesamum indicum) Seeds. Foods 2020, 9, 989. https://doi.org/10.3390/foods9080989
Kim TJ, Park JG, Kim HY, Ha S-H, Lee B, Park SU, Seo WD, Kim JK. Metabolite Profiling and Chemometric Study for the Discrimination Analyses of Geographic Origin of Perilla (Perilla frutescens) and Sesame (Sesamum indicum) Seeds. Foods. 2020; 9(8):989. https://doi.org/10.3390/foods9080989
Chicago/Turabian StyleKim, Tae Jin, Jeong Gon Park, Hyun Young Kim, Sun-Hwa Ha, Bumkyu Lee, Sang Un Park, Woo Duck Seo, and Jae Kwang Kim. 2020. "Metabolite Profiling and Chemometric Study for the Discrimination Analyses of Geographic Origin of Perilla (Perilla frutescens) and Sesame (Sesamum indicum) Seeds" Foods 9, no. 8: 989. https://doi.org/10.3390/foods9080989
APA StyleKim, T. J., Park, J. G., Kim, H. Y., Ha, S. -H., Lee, B., Park, S. U., Seo, W. D., & Kim, J. K. (2020). Metabolite Profiling and Chemometric Study for the Discrimination Analyses of Geographic Origin of Perilla (Perilla frutescens) and Sesame (Sesamum indicum) Seeds. Foods, 9(8), 989. https://doi.org/10.3390/foods9080989