

Alpha Lipoic Acid and Monoisoamyl-DMSA Combined Treatment Ameliorates Copper-Induced Neurobehavioral Deficits, Oxidative Stress, and Inflammation



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material & Methods
2.1. Animals
2.2. Chemicals
2.3. Chelation Studies
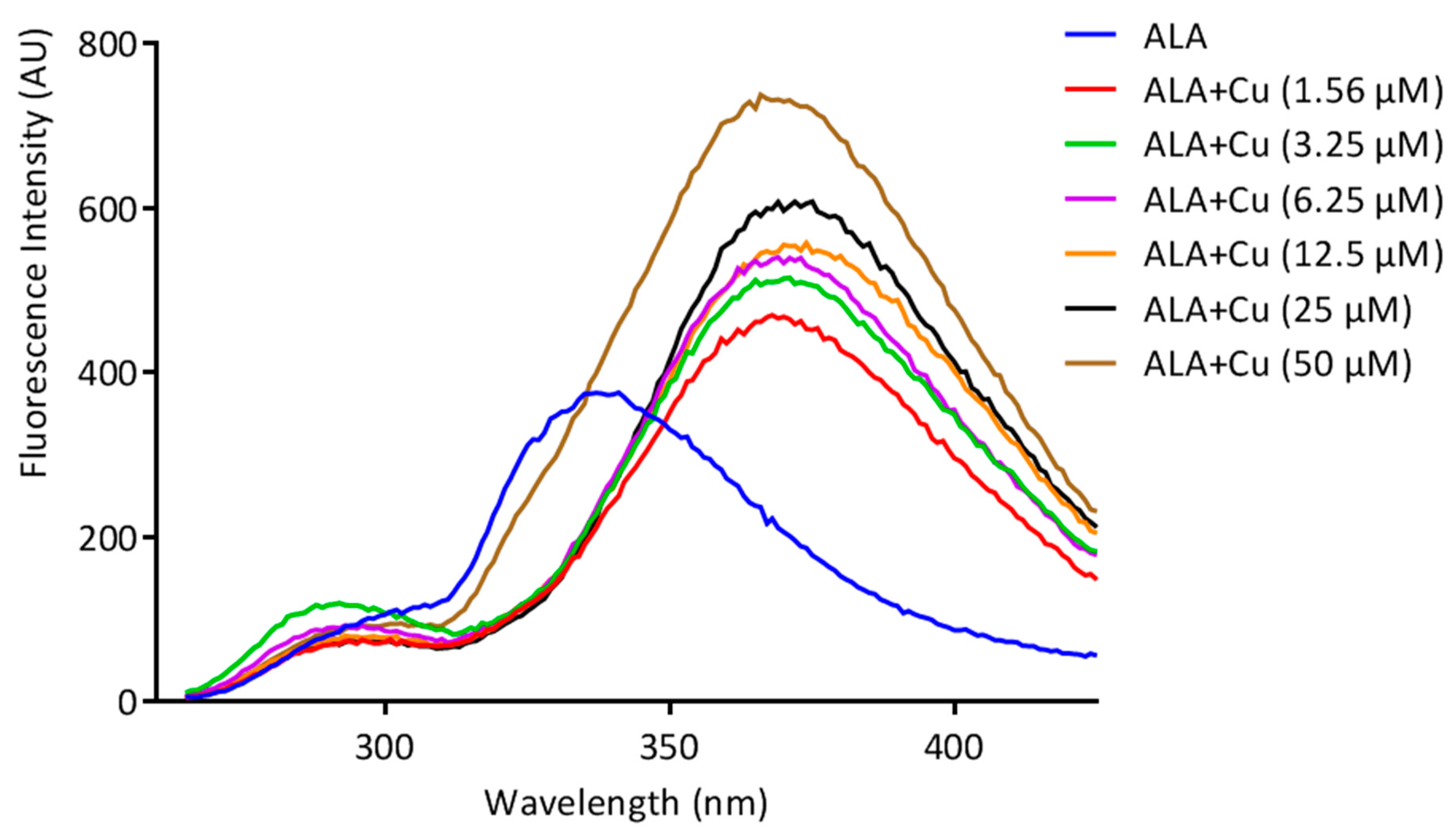
2.3.1. Fluorescence Spectroscopy
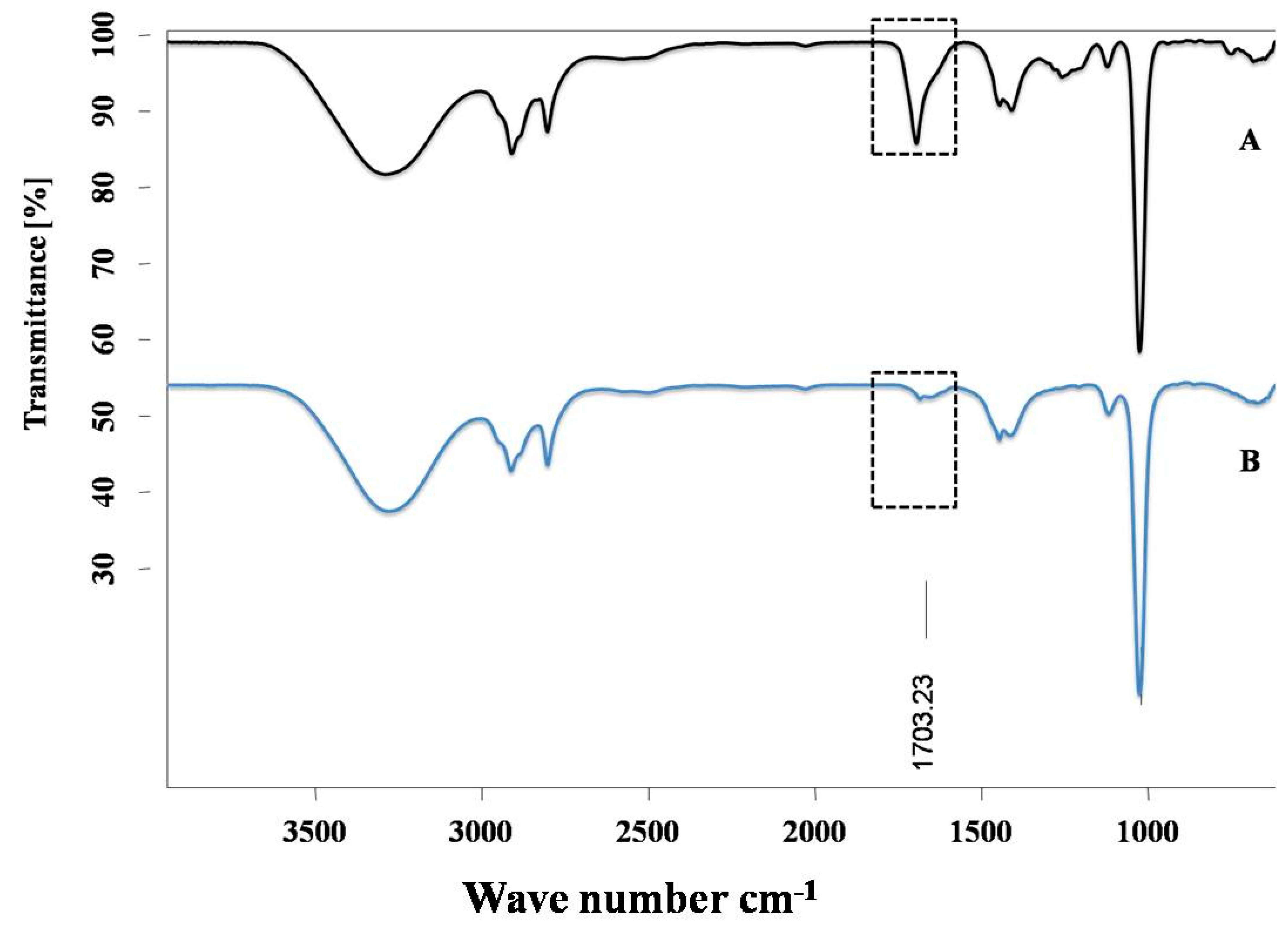
2.3.2. IR Spectroscopy
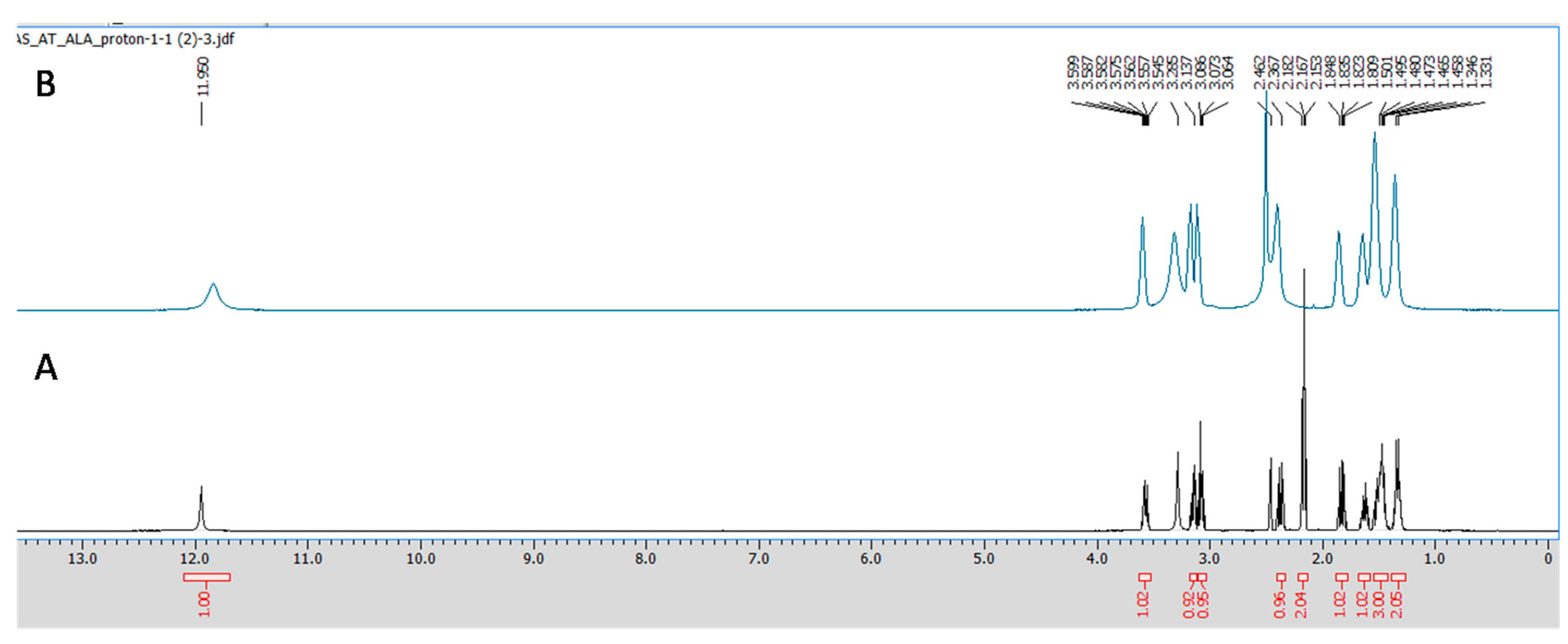
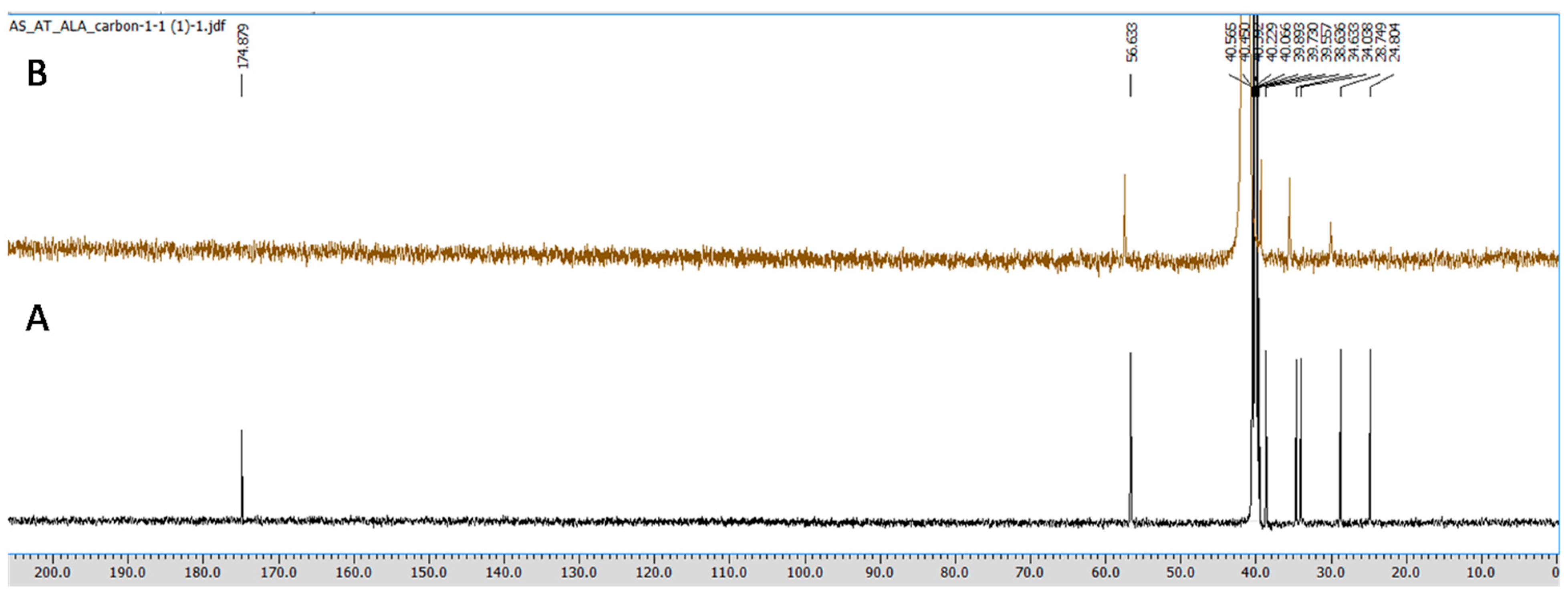
2.3.3. NMR Spectroscopy
2.3.4. Mass Spectroscopy
2.4. Experimental Design
- Group IA (n = 6): Saline (Control)
- Group IB (n = 6): MiADMSA (75 mg/kg, once, daily p.o.)
- Group IC (n = 6): ALA (75 mg/kg, once daily, p.o.)
- Group IIA, Cu-pre-exposed (n = 6): Saline (Copper sulfate 20 mg/kg)
- Group IIB, Cu-pre-exposed (n = 6): MiADMSA (75 mg/kg once daily, p.o.)
- Group IIC, Cu-pre-exposed (n = 6): ALA (75 mg/kg, once daily, p.o.)
- Group IID, Cu-pre-exposed (n = 6): ALA + MiADMSA (both 75 mg/kg, once daily, p.o.)
2.5. Assessment of Neurobehavioral Functions
2.5.1. Open Field Activity
2.5.2. Rotarod Test
2.5.3. Elevated plus Maze
2.5.4. Forced Swim Test (FST)
2.5.5. Nobel Object Recognition Test
2.5.6. Passive Avoidance Test
2.6. Estimation of Brain AChE Activity
2.7. Evaluation of Brain Oxidative Stress Markers
2.7.1. Reactive Oxygen Species
2.7.2. Thiobarbituric Acid Reactive Substances
2.7.3. Nitrite Levels
2.7.4. Superoxide Dismutase Activity
2.7.5. Catalase Activity
2.7.6. Total Protein Measurement
2.8. Measurement of Neuroinflammatory Markers (TNF-α, & IL-1β) by ELISA
2.9. Determination of Brain Cu Levels
2.10. Statistical Analysis
3. Results
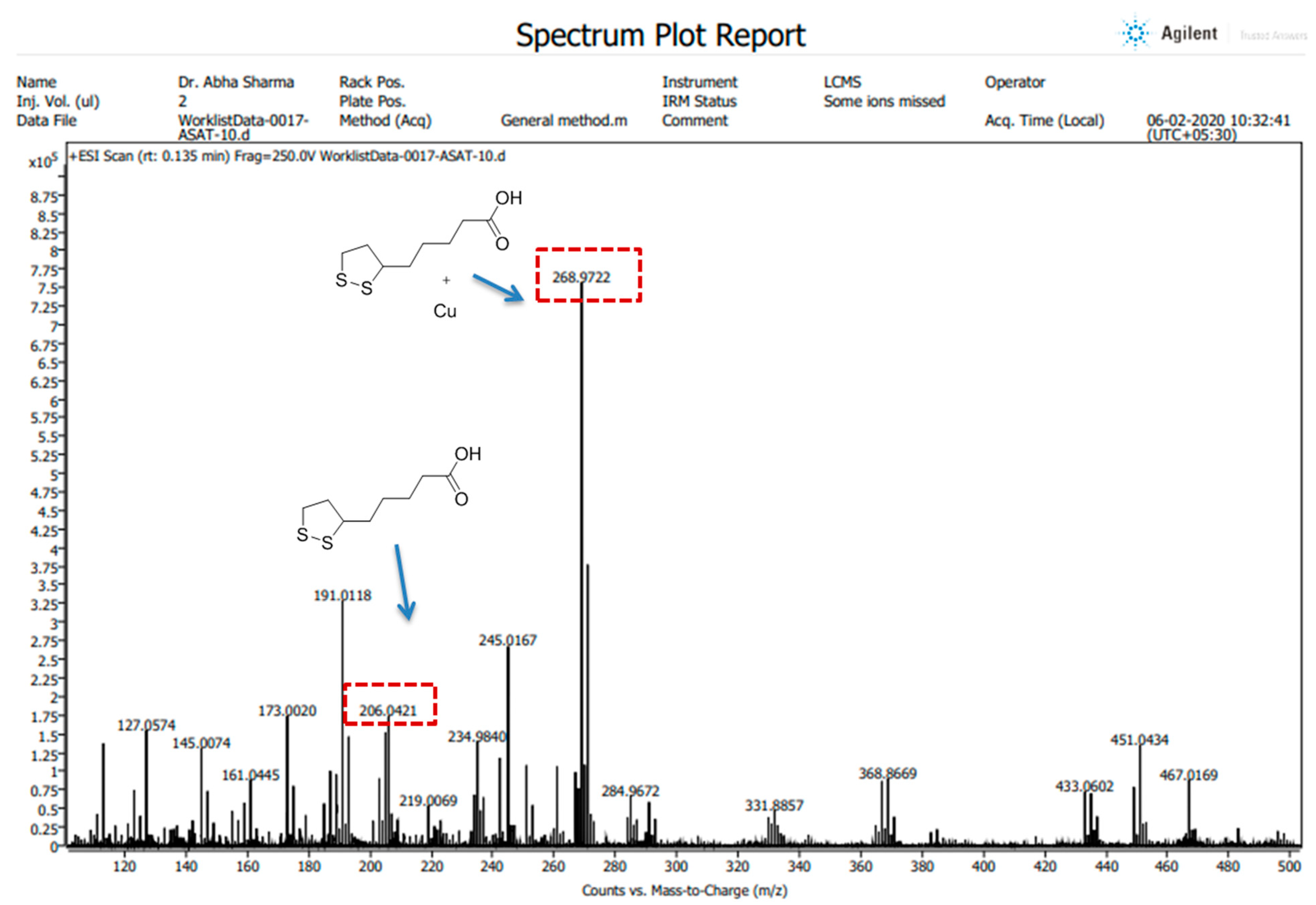
3.1. ALA/Cu Chelation Studies
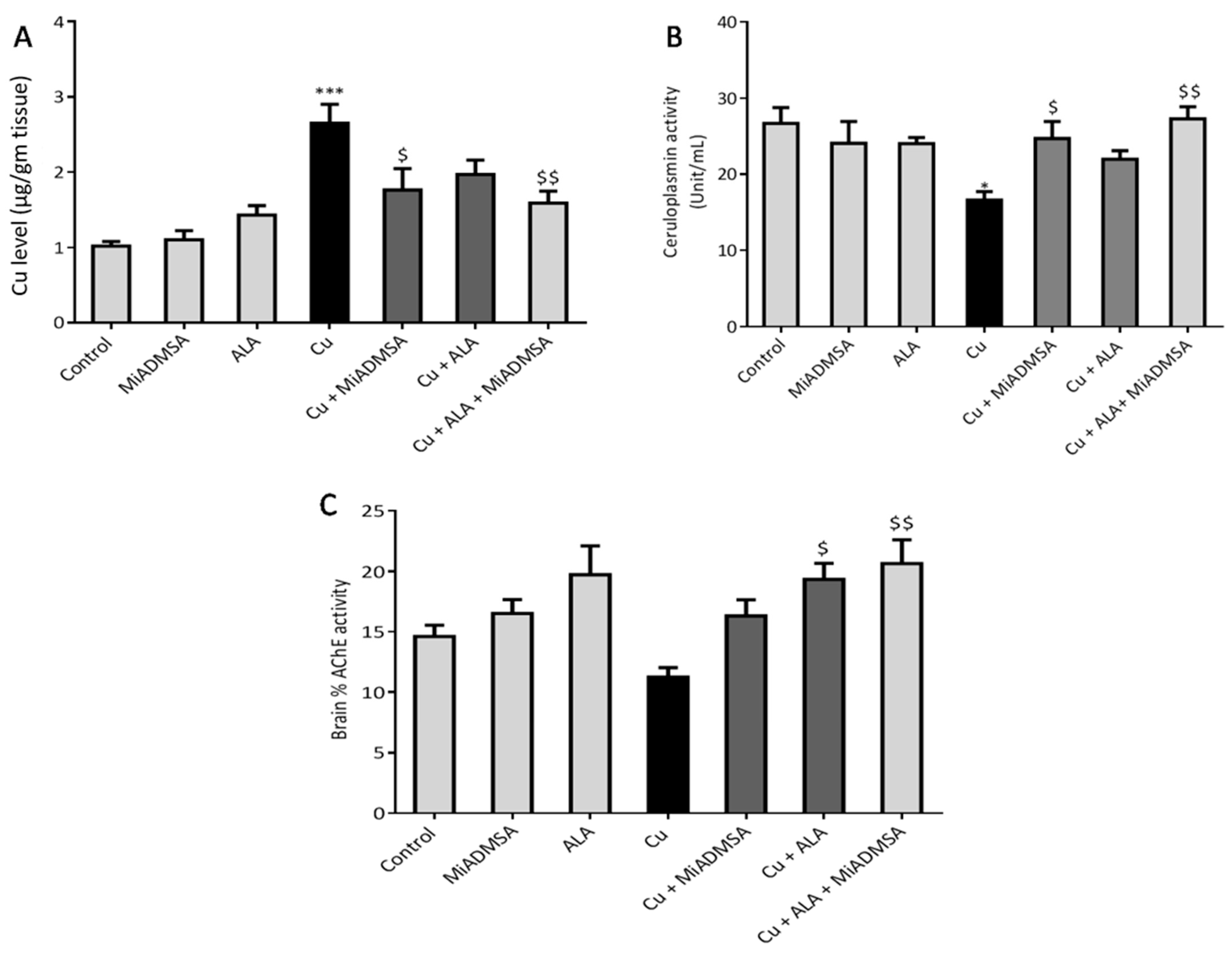
3.2. Effect of ALA& MiADMSA on Brain Cu, Serum Ceruloplasmin Levels, and AChE Levels
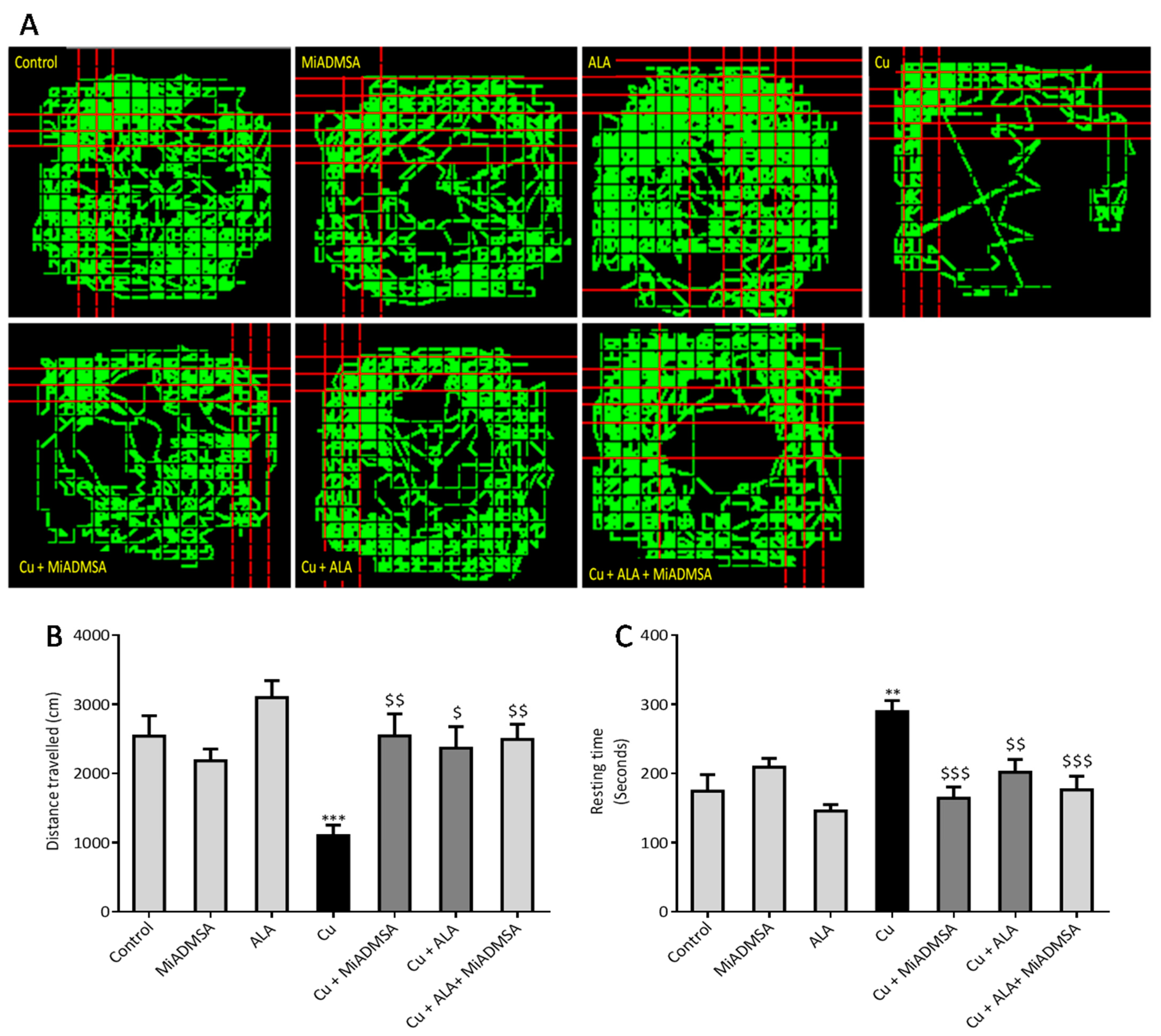
3.3. Effect of ALA and MiADMSA on Locomotors Activity
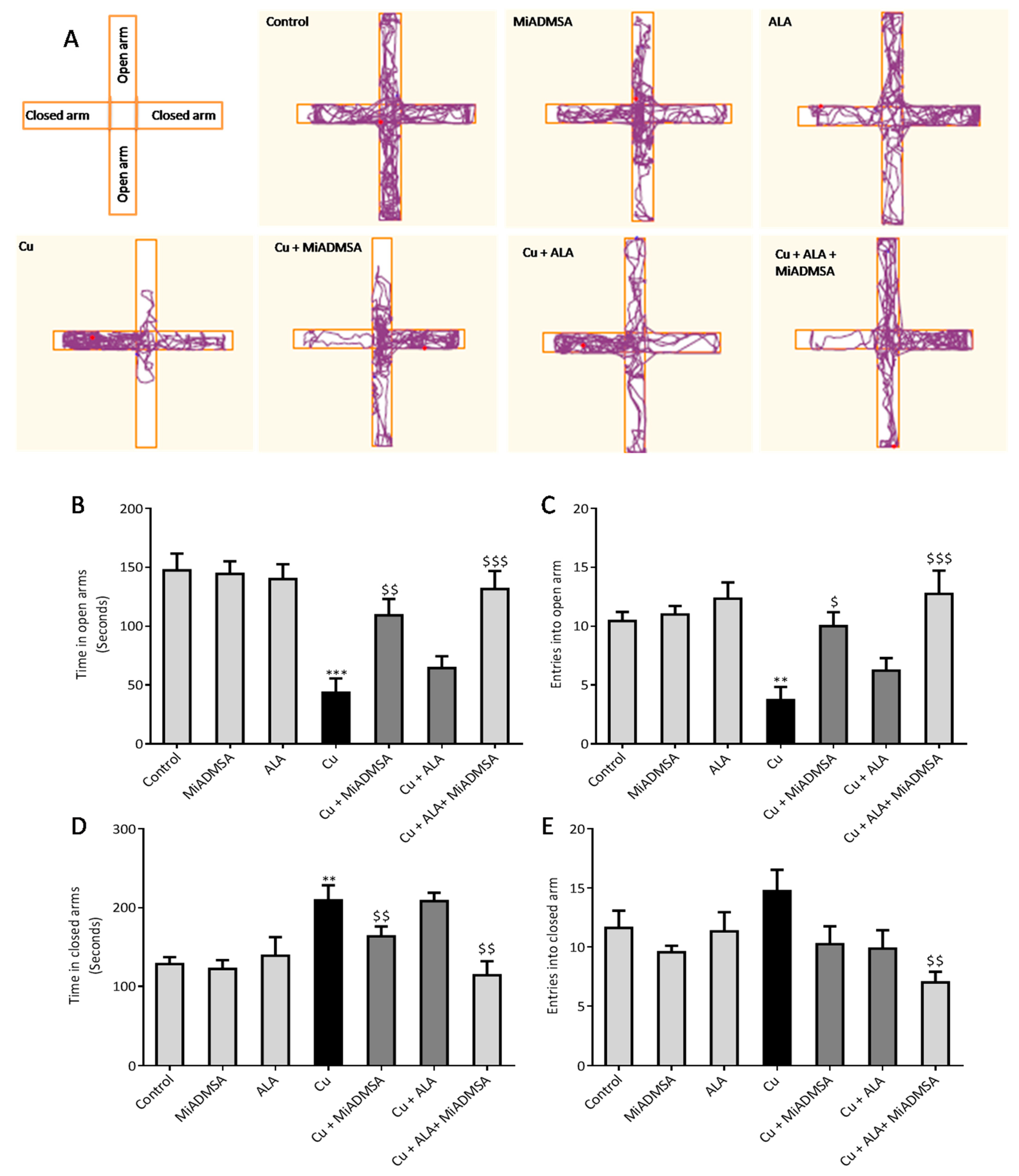
3.4. Effect of ALA and MiADMSA on EPM Performance
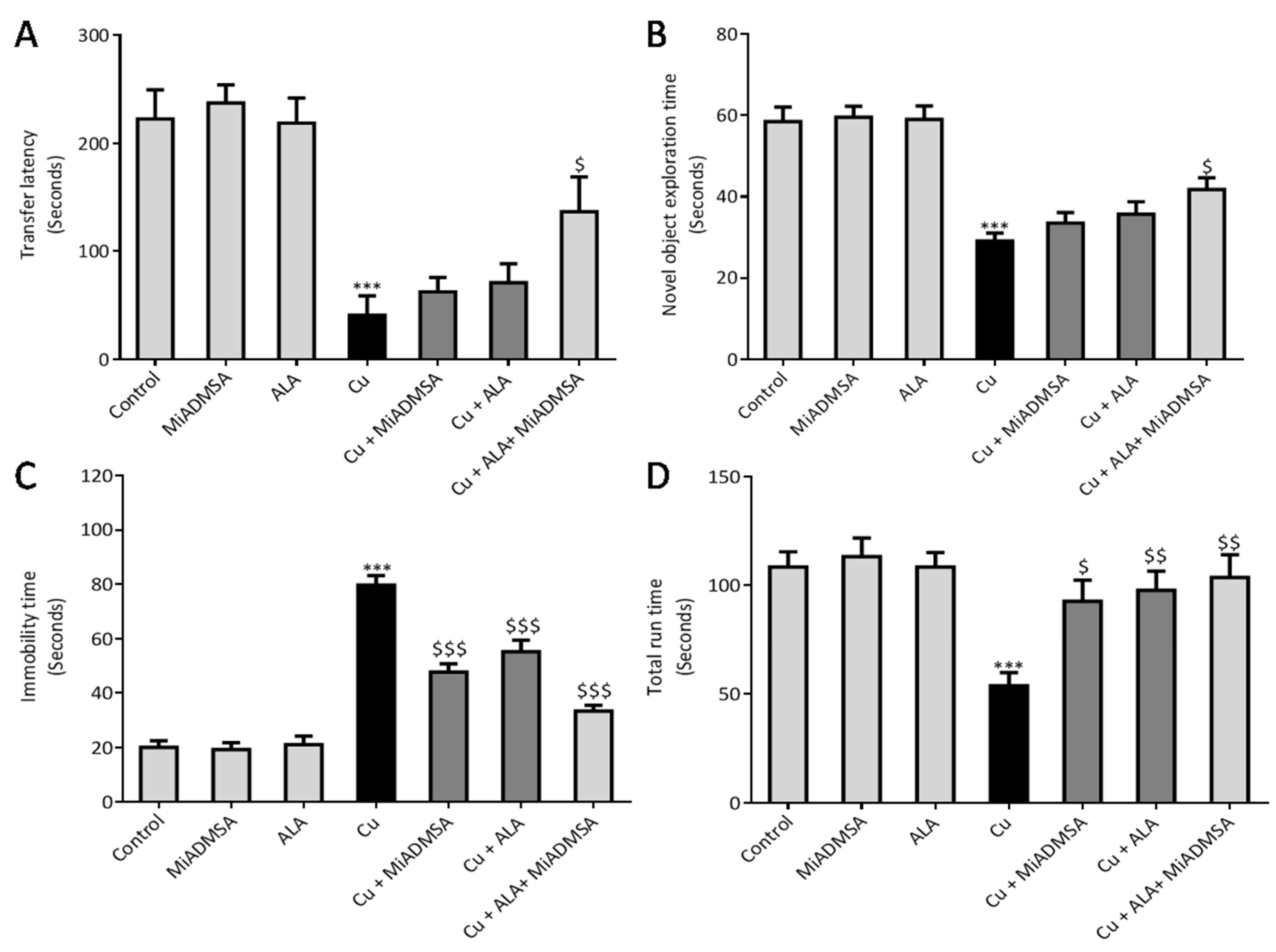
3.5. Effect of ALA and MiADMSA on Memory Functions and Other Neurobehavioral Activities
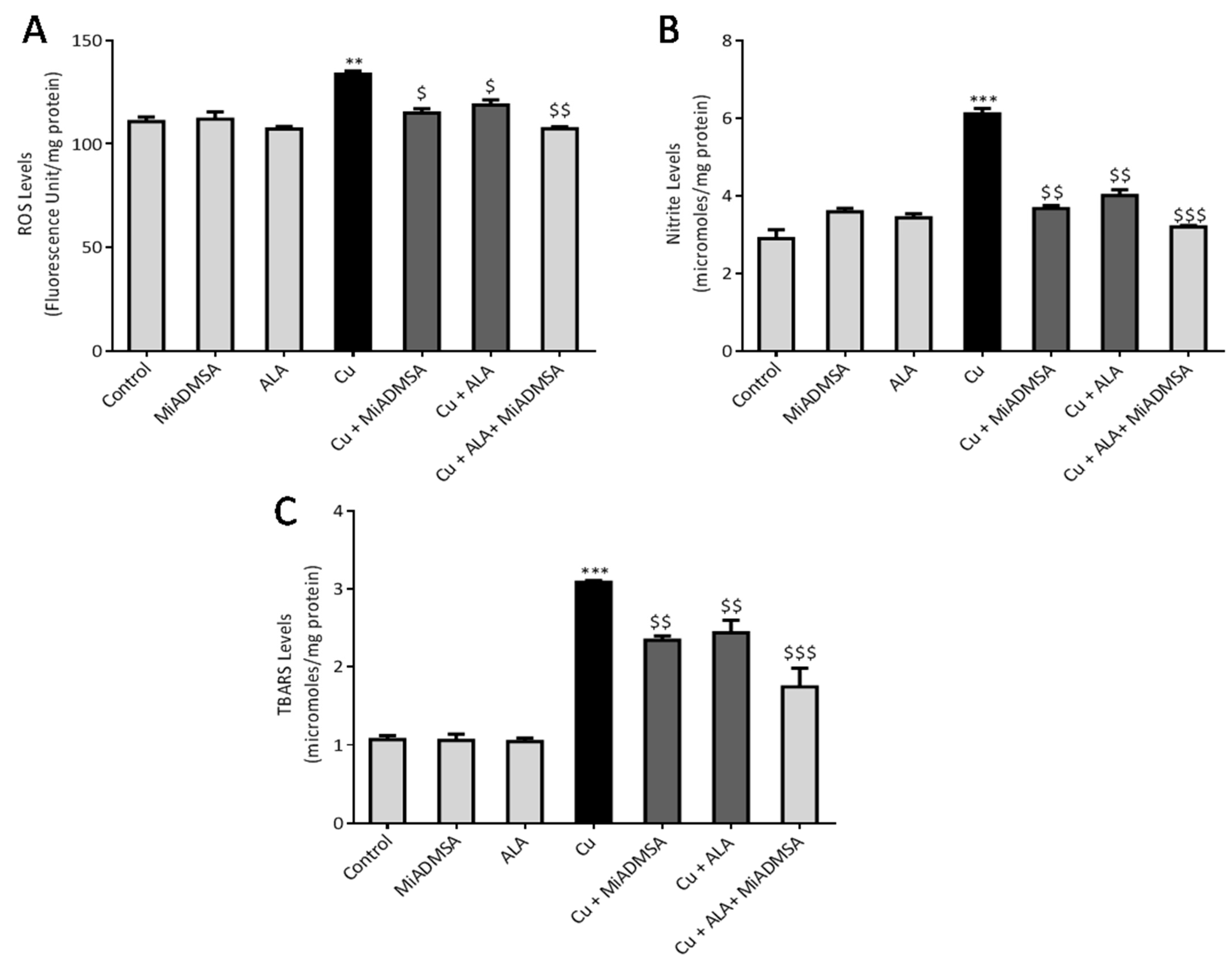
3.6. MiADMSA and ALA Therapy Mitigates the Oxidative Stress
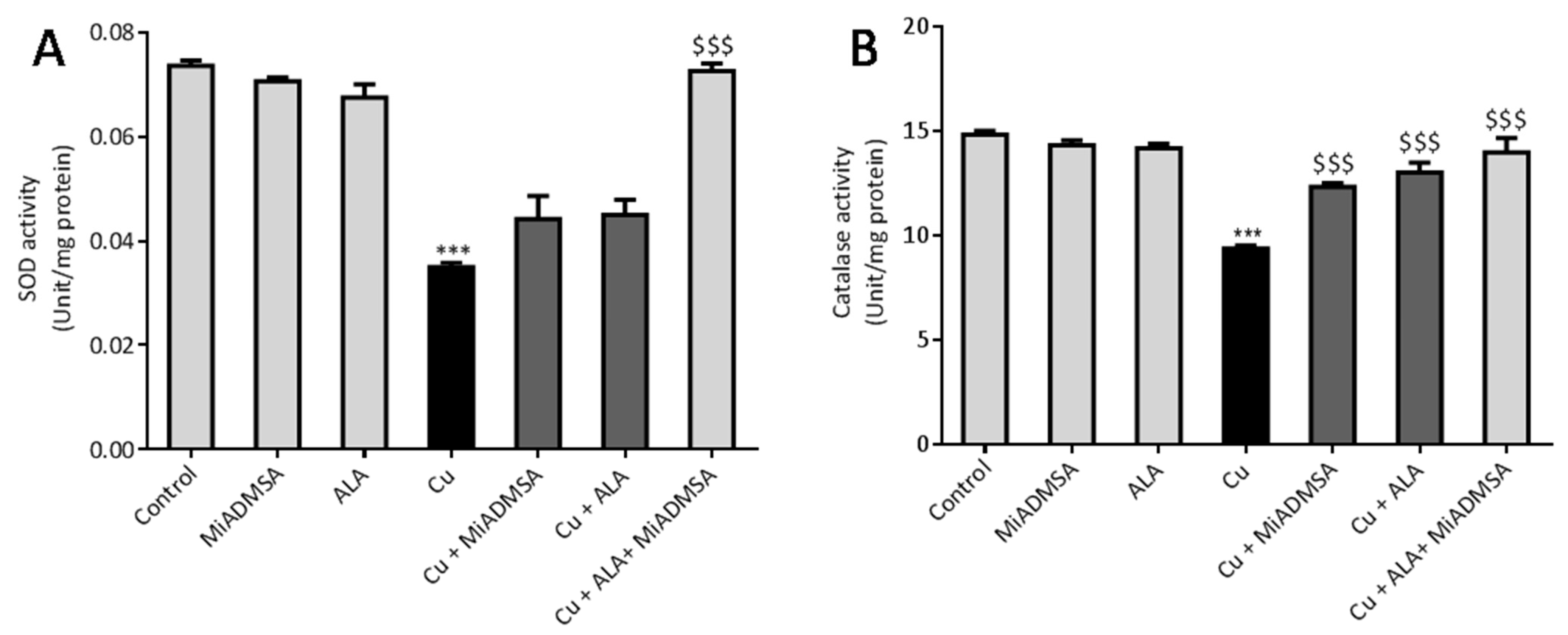
3.7. MiADMSA and ALA Restored the Altered Antioxidant Enzymes
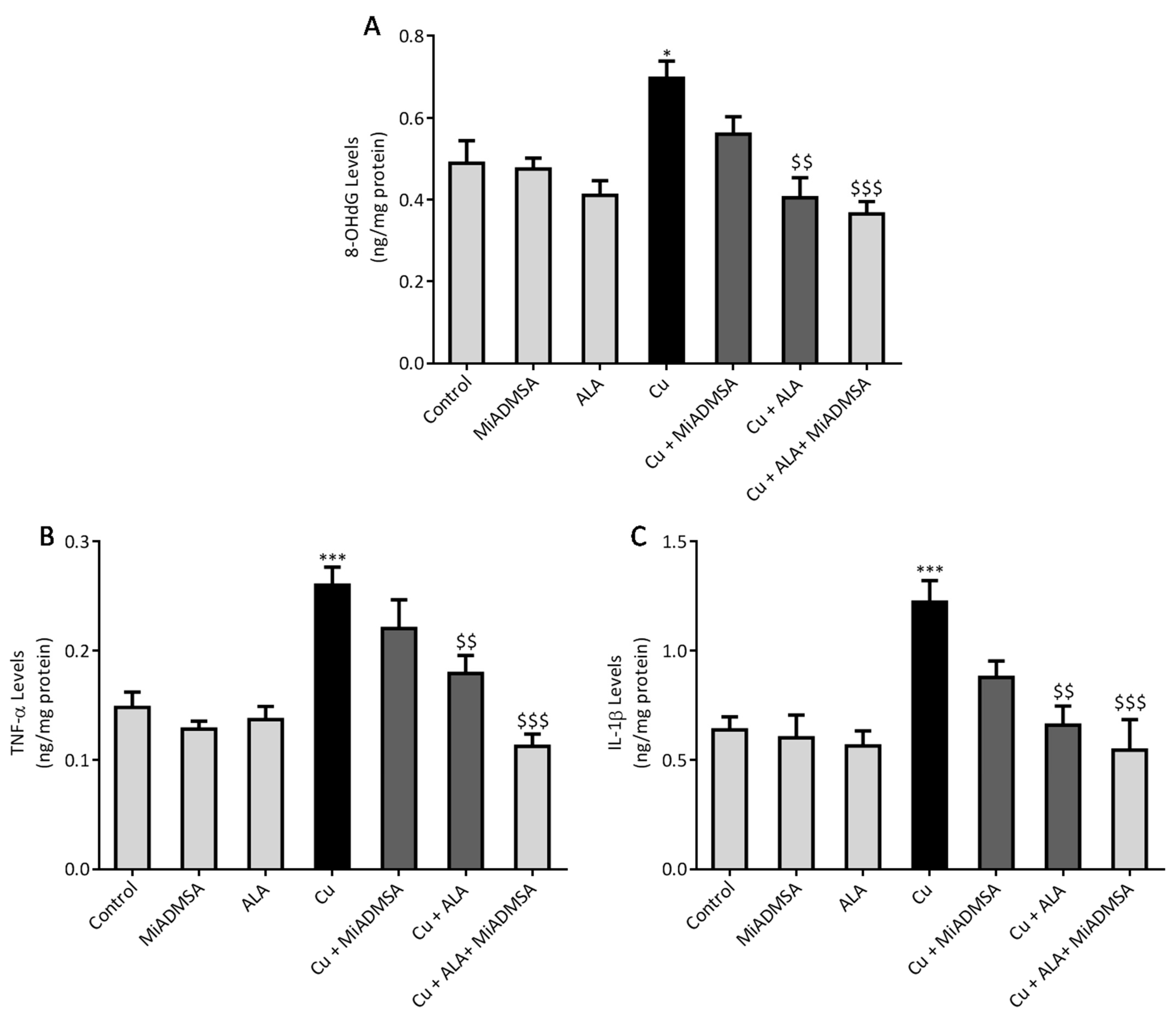
3.8. Effect of ALA and MiADMSA on Brain Inflammatory Markers
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| AChE | Acetyl cholinesterase |
| AD | Alzheimer’s disease |
| ALA | Alpha lipoic acid |
| Cu | Copper |
| DPA | D-Penicillamine |
| DTNB | 5,5′-Dithiobis(2-nitrobenzoic acid) |
| ELISA | Enzyme-linked immunosorbent assay |
| EPM | Elevated plus maze |
| FST | Forced swim test |
| GSH | Glutathione |
| HRMS | High-resolution mass spectrometry |
| IR | Infrared spectroscopy |
| MiADMSA | Monoisoamyl 2,3-dimercaptosuccinic acid |
| NF-κB | Nuclear factor-κb |
| NBT | Nitro blue tetrazolium |
| NO | Nitric oxide |
| NMR | Nuclear magnetic resonance |
| NORT | Novel object recognition test |
| 8-OHdG | 8-hydroxy-2′-deoxyguanosine |
| PACS | Passive avoidance task |
| PMS | Phenazine methosulfate |
| ROS | Reactive oxygen species |
| RPM | Revolutions per minute |
| SOD | Superoxide dismutase |
| TBARS | Thiobarbituric acid reactive substances |
| TNF-α | Tumour necrosis factor-alpha |
| WD | Wilson’s disease |
References
- Festa, R.A.; Thiele, D.J. Copper: An essential metal in biology. Curr. Biol. 2011, 21, R877–R883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackerman, C.M.; Chang, C.J. Copper signaling in the brain and beyond. Int. J. Biol. Chem. 2018, 293, 4628–4635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, V.; Kaler, S.G. Role of copper in human neurological disorders. Am. J. Clin. Nutr. 2008, 88, 855S–858S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathys, Z.K.; White, A.R. Copper and Alzheimer’s Disease. Adv. Neurobiol. 2017, 18, 199–216. [Google Scholar]
- Członkowska, A.; Litwin, T.; Dusek, P.; Ferenci, P.; Lutsenko, S.; Medici, V.; Rybakowski, J.K.; Weiss, K.H.; Schilsky, M.L. Wilson disease. Nat. Rev. Dis. Primers 2018, 4, 21. [Google Scholar] [CrossRef]
- Yang, F.; Liao, J.; Yu, W.; Pei, R.; Qiao, N.; Han, Q.; Hu, L.; Li, Y.; Guo, J.; Pan, J.; et al. Copper induces oxidative stress with triggered NF-κB pathway leading to inflammatory responses in immune organs of the chicken. Ecotoxicol. Environ. Saf. 2020, 200, 110715. [Google Scholar] [CrossRef]
- Pham, A.N.; Xing, G.; Miller, C.J.; Waite, T.D. Fenton-like copper redox chemistry revisited: Hydrogen peroxide and superoxide mediation of copper-catalyzed oxidant production. J. Catal. 2013, 301, 54–64. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2017, 94, 909–950. [Google Scholar] [CrossRef] [Green Version]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signaling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Kitazawa, M.; Hsu, H.W.; Medeiros, R. Copper Exposure Perturbs Brain Inflammatory Responses and Impairs Clearance of Amyloid-Beta. Toxicol. Sci. 2016, 152, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Flora, S.J.; Pachauri, V. Chelation in metal intoxication. Int. J. Environ. Res. Public Health 2010, 7, 2745–2788. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Kim, Y.S.; Kumar, V. Heavy metal toxicity: An update of chelating therapeutic strategies. J. Trace Elem. Med. Biol. 2019, 54, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Lawson, M.K.; Valko, M.; Cronin, M.T.D. Jomová, K. Chelators in Iron and Copper Toxicity. Curr. Pharmacol. Rep. 2016, 2, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Thakur, A.; Patwa, J.; Pant, S.; Sharma, A.; Flora, S.J.S. Interaction study of monoisoamyl dimercaptosuccinic acid with bovine serum albumin using biophysical and molecular docking approaches. Sci. Rep. 2021, 11, 4068. [Google Scholar] [CrossRef] [PubMed]
- Flora, S.; Mehta, A.; Gautam, P.; Jatav, P.; Pathak, U. Essential metal status, prooxidant/antioxidant effects of MiADMSA in male rats: Age-related effects. Biol. Trace Elem. Res. 2007, 120, 235–247. [Google Scholar] [CrossRef]
- Quamar, S.; Kumar, J.; Mishra, A.; Flora, S. Oxidative stress and neurobehavioural changes in rats following copper exposure and their response to MiADMSA and d-penicillamine. Toxicol. Res. Appl. 2019, 3, 2397847319844782. [Google Scholar] [CrossRef]
- Patwa, J.; Flora, S.J.S. MiADMSA abrogates chronic copper-induced hepatic and immunological changes in Sprague Dawley rats. Food Chem. Toxicol. 2020, 145, 111692. [Google Scholar] [CrossRef]
- Patwa, J.; Thakur, A.; Sharma, A.; Flora, S.J.S. Monoisoamyl DMSA reduced copper-induced neurotoxicity by lowering 8-OHdG level, amyloid beta and Tau protein expressions in Sprague-Dawley rats. Metallomics 2020, 12, 1428–1448. [Google Scholar] [CrossRef]
- Molz, P.; Schröder, N. Potential Therapeutic Effects of Lipoic Acid on Memory Deficits Related to Aging and Neurodegeneration. Front. Pharmacol. 2017, 8, 849. [Google Scholar] [CrossRef] [Green Version]
- Maczurek, A.; Hager, K.; Kenklies, M.; Sharman, M.; Martins, R.; Engel, J.; Carlson, D.A.; Münch, G. Lipoic acid as an anti-inflammatory and neuroprotective treatment for Alzheimer’s disease. Adv. Drug Deliv. Rev. 2008, 60, 1463–1470. [Google Scholar] [CrossRef]
- Pande, M.; Flora, S.J. Lead induced oxidative damage and its response to combined administration of alpha-lipoic acid and succimers in rats. Toxicology 2002, 177, 187–196. [Google Scholar] [CrossRef]
- Wang, T.; Cheng, J.; Wang, S.; Wang, X.; Jiang, H.; Yang, Y.; Wang, Y.; Zhang, C.; Liang, W.; Feng, H. α-Lipoic acid attenuates oxidative stress and neurotoxicity via the ERK/Akt-dependent pathway in the mutant hSOD1 related Drosophila model and the NSC34 cell line of amyotrophic lateral sclerosis. Brain Res. Bull. 2018, 140, 299–310. [Google Scholar] [CrossRef]
- Smirnova, J.; Kabin, E.; Järving, I.; Bragina, O.; Tõugu, V.; Plitz, T.; Palumaa, P. Copper(I)-binding properties of de-coppering drugs for the treatment of Wilson disease. α-Lipoic acid as a potential anti-copper agent. Sci. Rep. 2018, 8, 1463. [Google Scholar] [CrossRef]
- Kumar, J.; Sathua, K.B.; Flora, S.J.S. Chronic copper exposure elicit neurotoxic responses in rat brain: Assessment of 8-hydroxy-2-deoxyguanosine activity, oxidative stress and neurobehavioral parameters. Cell. Mol. Biol. 2019, 65, 27–35. [Google Scholar] [CrossRef]
- Masuo, Y.; Matsumoto, Y.; Morita, S.; Noguchi, J. A novel method for counting spontaneous motor activity in the rat. Brain Res. Brain Res. Protoc. 1997, 1, 321–326. [Google Scholar] [CrossRef]
- Shiotsuki, H.; Yoshimi, K.; Shimo, Y.; Funayama, M.; Takamatsu, Y.; Ikeda, K.; Takahashi, R.; Kitazawa, S.; Hattori, N. A rotarod test for evaluation of motor skill learning. J. Neurosci. Methods 2010, 189, 180–185. [Google Scholar] [CrossRef]
- Walf, A.A.; Frye, C.A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef] [Green Version]
- Slattery, D.A.; Cryan, J.F. Using the rat forced swim test to assess antidepressant-like activity in rodents. Nat. Protoc. 2012, 7, 1009–1014. [Google Scholar] [CrossRef]
- Antunes, M.; Biala, G. The novel object recognition memory: Neurobiology, test procedure, and its modifications. Cogn. Process. 2012, 13, 93–110. [Google Scholar] [CrossRef] [Green Version]
- Elrod, K.; Buccafusco, J.J. An evaluation of the mechanism of scopolamine-induced impairment in two passive avoidance protocols. Pharmacol. Biochem. Behav. 1988, 29, 15–21. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Feather-Stone, R.M., Jr. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Degli Esposti, M. Measuring mitochondrial reactive oxygen species. Methods (S.-Diego, Calif.) 2002, 26, 335–340. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Giustarini, D.; Rossi, R.; Milzani, A.; Dalle-Donne, I. Nitrite and nitrate measurement by Griess reagent in human plasma: Evaluation of interferences and standardization. Methods Enzymol. 2008, 440, 361–380. [Google Scholar] [PubMed]
- Kakkar, P.; Das, B.; Viswanathan, P.N. A modified spectrophotometric assay of superoxide dismutase. Indian J. Biochem. Biophys. 1984, 21, 130–132. [Google Scholar]
- Góth, L. A simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta 1991, 196, 143–151. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Sathua, K.B.; Patwa, J.K.; Flora, S.J.S. Oxidative/ Nitrosative Stress, 8-OHdG and MMP-9: The Possible Co-Links and Early Sign of Arsenic Induced Urinary Bladder Carcinogenesis in Experimental Rats. Free Radic. Antioxid. 2019, 9, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Esmieu, C.; Guettas, D.; Conte-Daban, A.; Sabater, L.; Faller, P.; Hureau, C. Copper-Targeting Approaches in Alzheimer’s Disease: How To Improve the Fallouts Obtained from in Vitro Studies. Inorg. Chem. 2019, 58, 13509–13527. [Google Scholar] [CrossRef] [Green Version]
- Naqvi, S.; Kumar, P.; Flora, S.J.S. Comparative efficacy of Nano and Bulk Monoisoamyl DMSA against arsenic-induced neurotoxicity in rats. Biomed. Pharmacol. 2020, 132, 110871. [Google Scholar] [CrossRef]
- Bai, L.; Xu, Y.; Li, G.; Tian, S.; Li, L.; Tao, F.; Deng, A.; Wang, S.; Wang, L. A Highly Selective Turn-on and Reversible Fluorescent Chemosensor for Al3+ Detection Based on Novel Salicylidene Schiff Base-Terminated PEG in Pure Aqueous Solution. Polymers 2019, 11, 573. [Google Scholar] [CrossRef] [Green Version]
- Lica, G.C.; Zelakiewicz, B.S.; Tong, Y.Y. Electrochemical and NMR characterization of octanethiol-protected Au nanoparticles. J. Electroanal. Chem. 2003, 554–555, 127–132. [Google Scholar] [CrossRef]
- Bjørklund, G.; Crisponi, G.; Nurchi, V.M.; Cappai, R.; Buha Djordjevic, A.; Aaseth, J. A Review on Coordination Properties of Thiol-Containing Chelating Agents Towards Mercury, Cadmium, and Lead. Molecules 2019, 24, 3247. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, N.; Flora, G.; Kushwaha, P.; Flora, S.J. Alpha-lipoic acid protects oxidative stress, changes in cholinergic system and tissue histopathology during co-exposure to arsenic-dichlorvos in rats. Environ. Toxicol. Pharmacol. 2014, 37, 7–23. [Google Scholar] [CrossRef]
- Sensi, S.L.; Granzotto, A.; Siotto, M.; Squitti, R. Copper and Zinc Dysregulation in Alzheimer’s Disease. Trends Pharmacol. Sci. 2018, 39, 1049–1063. [Google Scholar] [CrossRef]
- Coelho, F.C.; Squitti, R.; Ventriglia, M.; Cerchiaro, G.; Daher, J.P.; Rocha, J.G.; Rongioletti, M.C.A.; Moonen, A.-C. Agricultural Use of Copper and Its Link to Alzheimer’s Disease. Biomolecules 2020, 10, 897. [Google Scholar] [CrossRef]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [Green Version]
- Farr, S.A.; Price, T.O.; Banks, W.A.; Ercal, N.; Morley, J.E. Effect of alpha-lipoic acid on memory, oxidation, and lifespan in SAMP8 mice. J. Alzheimer’s Dis. 2012, 32, 447–455. [Google Scholar] [CrossRef]
- Karimani, A.; Ramezani, N.; Afkhami Goli, A.; Nazem Shirazi, M.H.; Nourani, H.; Jafari, A.M. Subchronic neurotoxicity of diazinon in albino mice: Impact of oxidative stress, AChE activity, and gene expression disturbances in the cerebral cortex and hippocampus on mood, spatial learning, and memory function. Toxicol. Rep. 2021, 8, 1280–1288. [Google Scholar] [CrossRef]
- Eskici, G.; Axelsen, P.H. Copper and Oxidative Stress in the Pathogenesis of Alzheimer’s Disease. Biochemistry 2012, 51, 6289–6311. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Moini, H.; Packer, L.; Saris, N.-E.L. Antioxidant and Prooxidant Activities of α-Lipoic Acid and Dihydrolipoic Acid. Toxicol. App. Pharmacol. 2002, 182, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Persichini, T.; Percario, Z.; Mazzon, E.; Colasanti, M.; Cuzzocrea, S.; Musci, G. Copper activates the NF-kappaB pathway in vivo. Antioxid. Redox Signal. 2006, 8, 1897–1904. [Google Scholar] [CrossRef] [PubMed]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patwa, J.; Thakur, A.; Flora, S.J.S. Alpha Lipoic Acid and Monoisoamyl-DMSA Combined Treatment Ameliorates Copper-Induced Neurobehavioral Deficits, Oxidative Stress, and Inflammation. Toxics 2022, 10, 718. https://doi.org/10.3390/toxics10120718
Patwa J, Thakur A, Flora SJS. Alpha Lipoic Acid and Monoisoamyl-DMSA Combined Treatment Ameliorates Copper-Induced Neurobehavioral Deficits, Oxidative Stress, and Inflammation. Toxics. 2022; 10(12):718. https://doi.org/10.3390/toxics10120718
Chicago/Turabian StylePatwa, Jayant, Ashima Thakur, and Swaran Jeet Singh Flora. 2022. "Alpha Lipoic Acid and Monoisoamyl-DMSA Combined Treatment Ameliorates Copper-Induced Neurobehavioral Deficits, Oxidative Stress, and Inflammation" Toxics 10, no. 12: 718. https://doi.org/10.3390/toxics10120718
APA StylePatwa, J., Thakur, A., & Flora, S. J. S. (2022). Alpha Lipoic Acid and Monoisoamyl-DMSA Combined Treatment Ameliorates Copper-Induced Neurobehavioral Deficits, Oxidative Stress, and Inflammation. Toxics, 10(12), 718. https://doi.org/10.3390/toxics10120718