
What Caused Declines in Intertidal Invertebrate Populations around Fukushima Daiichi Nuclear Power Plant after the 2011 Great East Japan Earthquake, Tsunami, and Nuclear Disaster?

Abstract
:1. Possible Impacts to Marine Organisms around Fukushima Daiichi Nuclear Power Plant after the 2011 Great East Japan Earthquake, Tsunami, and Nuclear Disaster
2. Effects of Radionuclides and Ionizing Radiation on Aquatic Organisms
3. The Gap between Effects Observed in the Laboratory and Those in Aquatic Environments
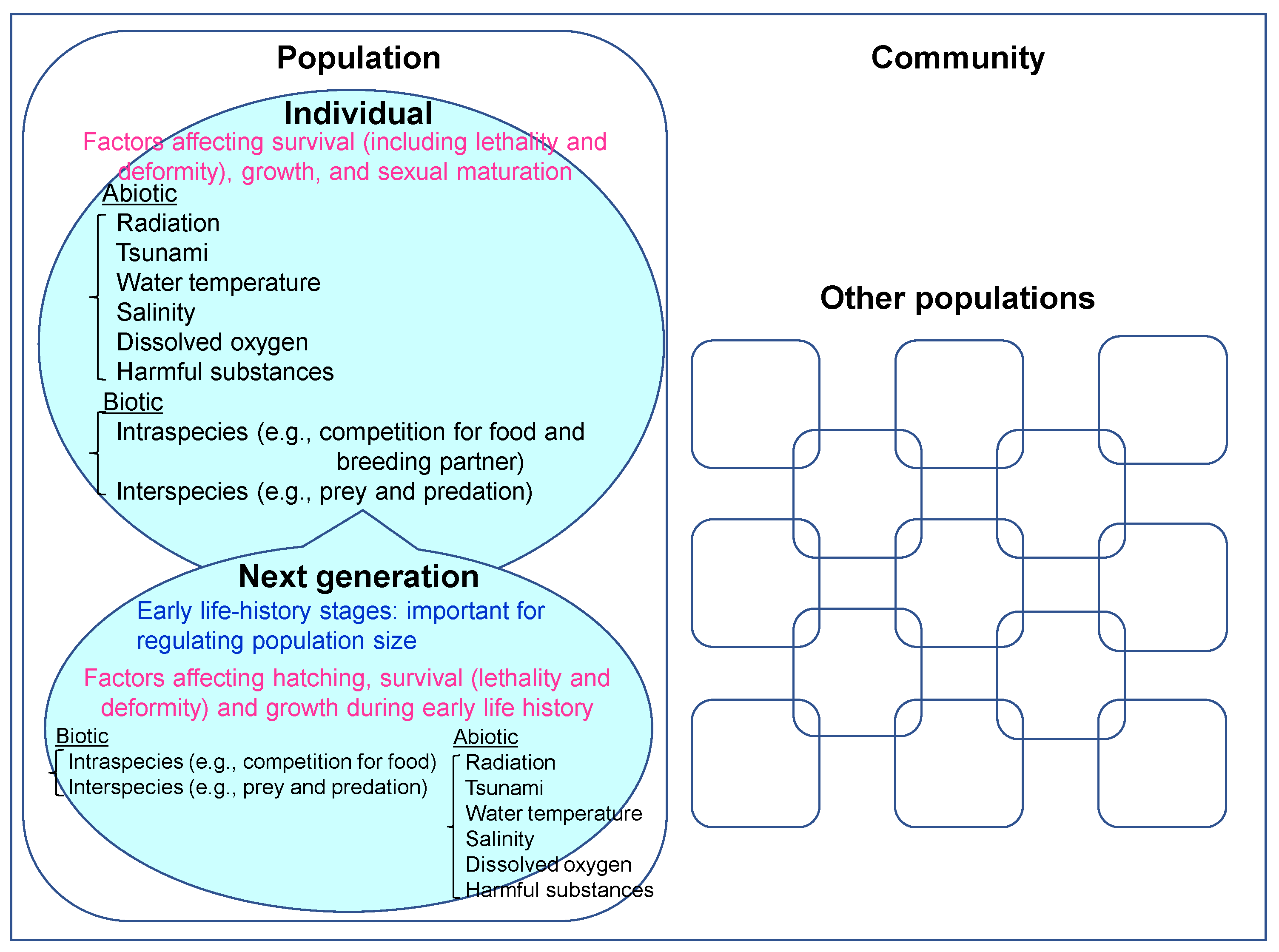
4. Effects of Biotic and Abiotic Factors Other than Ionizing Radiation on Aquatic Organisms
5. Existing Knowledge on Combined Effects of Ionizing Radiation and Other Environmental Factors
6. The Environment Is a Complex System: The Necessity of Evaluating Combined Effects of Ionizing Radiation and Other Biotic and Abiotic Environmental Factors for Population-Level Assessments
- (1)
- Limitations on extrapolation from the effects observed in aquatic organisms exposed at relatively high doses/dose rates in the laboratory to ecologically relevant effects observed in aquatic organisms in the field, which could be exposed chronically at much lower doses/dose rates.
- (2)
- Insufficient consideration of the combined effects of ionizing radiation with other abiotic and biotic environmental factors together with the life-history traits of the species examined, for assessment of population-level effects.
- (3)
- The misapprehension that population-level effects can be assessed by simply extending the individual-level effects on adults to the next generation.
- (1)
- Decline in population densities as well as the number of species. Radionuclides (including short- and long-lived ones) are candidates as causal factors, as discussed by Horiguchi et al. [3]. Even if there was no mass mortality in intertidal invertebrate populations near FDNPP immediately after the nuclear accident, it is possible that long-term survival of individuals exposed to radiation might have been difficult with the possible depletion of prey and the unchanging, harsh intertidal environment along the coast of Fukushima. Although boric acid and hydrazine, which were also thought to have been released from FDNPP into the sea at the time of the accident, might also be causal factors, we believe it unlikely that they would have caused mass mortality in intertidal invertebrate populations near FDNPP after the nuclear accident, because seawater already contains much boron, and hydrazine decomposes rapidly in sunlight.
- (2)
- Delayed recovery from the decline in population densities and the number of species. Several heavy metals and a possible increase in pH resulting from the cement used in the construction to cover the contaminated bottom sediment in the harbor at FDNPP may be causal factors reducing the survival rates of invertebrate larvae recruiting from remote areas to the intertidal zone near FDNPP [4]. The possible increase in turbidity due to the construction may also be a factor [4].
- (3)
- Continuous sexual maturation in the rock shell population. It is unlikely that radionuclides that leaked from FDNPP or high seawater temperatures are causal factors for these observed effects, as discussed in Horiguchi et al. [5]. However, it is possible that some unknown harmful substance(s) that may have leaked from FDNPP may be related to the induction/promotion and continued sexual maturation, or the failure to terminate maturation, in the rock shell population near FDNPP throughout the year. Unknown changes in physiological characteristics of the rock shell near FDNPP may be involved. There might also be a trade-off in the allocation of available energy resources between reproduction and somatic growth, because changes in life-history traits and population dynamics of marine organisms in response to environmental change are known to occur in nature, although the causes underlying such responses and the mechanisms remain unidentified [72].
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Garnier-Laplace, J.; Beaugelin-Seiller, K.; Hinton, T.G. Fukushima wildlife dose reconstruction signals ecological consequences. Environ. Sci. Technol. 2011, 45, 5077–5078. [Google Scholar] [CrossRef]
- Vives i Batlle, J. Exposures and Effects in the Marine Environment after the Fukushima Accident. ICRP 2013 Proceedings. Available online: https://www.icrp.org/docs/ICRP2013%20S6%20Vives%20i%20Batlle.pdf (accessed on 2 March 2022).
- Horiguchi, T.; Yoshii, H.; Mizuno, S.; Shiraishi, H. Decline in intertidal biota after the 2011 Great East Japan Earthquake and Tsunami and the Fukushima nuclear disaster: Field observations. Sci. Rep. 2016, 6, 20416. [Google Scholar] [CrossRef] [Green Version]
- Horiguchi, T.; Kodama, K.; Kume, G.; Kang, I.J. Chapter 6, Delayed recovery from declines in the population densities and species richness of intertidal invertebrates near Fukushima Daiichi Nuclear Power Plant. In Low-Dose Radiation Effects on Animals and Ecosystems, Long-Term Study on the Fukushima Nuclear Accident; Fukumoto, M., Ed.; Springer: Tokyo, Japan, 2019; pp. 65–88. [Google Scholar]
- Horiguchi, T.; Kawamura, K.; Ohta, Y. Consecutive sexual maturation observed in a rock shell population in the vicinity of the Fukushima Daiichi Nuclear Power Plant, Japan. Sci. Rep. 2021, 11, 560. [Google Scholar] [CrossRef] [PubMed]
- Kodama, K.; Aramaki, T.; Horiguchi, T. Current status of the megabenthic community in coastal Fukushima Prefecture, Japan, in the wake of the Great East Japan Earthquake. Mar. Environ. Res. 2018, 140, 358–374. [Google Scholar] [CrossRef]
- Etoh, H. Effects by internal irradiation in fish. In Radioactivity and Fish; Egami, N., Ed.; Koseisha-Koseikaku: Tokyo, Japan, 1972; pp. 298–316. (In Japanese) [Google Scholar]
- Ophel, I.L.; Hoppenheit, M.; Ichikawa, R.; Klimov, A.G.; Kobayashi, S.; Nishiwaki, Y.; Saiki, M. Chapter 2, Effects of ionizing radiation on aquatic organisms. In Effects of Ionizing Radiation on Aquatic Organisms and Ecosystems; International Atomic Energy Agency (IAEA) Technical Report Series, No. 172; International Atomic Energy Agency: Vienna, Austria, 1976; pp. 55–86. Available online: https://inis.iaea.org/collection/NCLCollectionStore/_Public/07/278/7278755.pdf (accessed on 2 March 2022).
- Lebedeva, G.D.; Sinevid, S.G. The effect of radioactive strontium upon survival and propagation in Daphnia magna. Dokl. Akad. Nauk SSSR 1958, 122, 586. (In Russian) [Google Scholar]
- Telichenko, M.M. Chronic effects of small doses of 238U, 232Th and 89+90Sr on a number of generations of Daphnia magna Straus. Nauch. Dokl. Vyssh. Shk. 1958, 1, 114. (In Russian) [Google Scholar]
- Polikarpov, G.G.; Ivanov, V.N. The effect of 90Sr–90Y on the developing anchovy eggs. Vopr. Ikhtiol. 1961, 1, 583. [Google Scholar]
- Polikarpov, G.G.; Ivanov, V.N. The harmful effect of 90Sr–90Y in the early development of the red mullet, the green wrasse, the horse mackerel and the Black Sea anchovy. Dokl. Akad. Nauk SSSR 1962, 144, 219. [Google Scholar]
- Fedorov, A.F.; Podymakhin, V.N.; Shchitenko, N.T.; Chumachenko, V.V. The influence of low radioactive contamination of water on the development of Pleuronectes platessa L. Vopr. Ikhtiol. 1964, 4, 579. (In Russian) [Google Scholar]
- Brown, V.M.; Templeton, W.L. Resistance of fish embryos to chronic irradiation. Nature 1964, 203, 1257. [Google Scholar] [CrossRef]
- Neustroev, G.V.; Podymakhin, V.N. On the rate of development of salmon (Salmo salar L.) roe under conditions of radioactive pollution of the hydrosphere with 90Sr + 90Y. Radiobiologiya 1966, 6, 321. [Google Scholar]
- Kulikov, N.V.; Timofeeva, N.A.; Lyubimova, S.A. Influence of ionizing radiation on the early stages on the development of Limnea stagnalis. Radiobiologiya 1966, 6, 908. [Google Scholar]
- Hiyama, Y.; Shimizu, M.; Suyama, I. Radiation effects on hatching of fish eggs. In Report on the Influence of Low Concentrations of Radionuclides in Water to Aquatic Organisms; Nuclear Safety Research Association of Japan: Tokyo, Japan, 1970. (In Japanese) [Google Scholar]
- Egami, N.; Etoh, H.; Hyodo-Taguchi, Y.; Tanaka, Y.; Yoshimura, N. Effects of low radioactive effluent on fisheries propagation. In Report on the Influence of Low Concentrations of Radionuclides in Water to Aquatic Organisms; Nuclear Safety Research Association of Japan: Tokyo, Japan, 1970. (In Japanese) [Google Scholar]
- Kosheleva, V.V. Change in peripheral blood in the embryos and larvae of Atlantic salmon under the influence of radioactive contamination of the water and X-rays. In Effect of Ionizing Radiation on the Organism; Sorokin, B.P., Ed.; AEC-tr-7418; US Atomic Energy Comission: Washington, DC, USA, 1973; p. 75. [Google Scholar]
- Kosheleva, V.V. Accumulation of radioactive isotopes by the developing eggs of the Atlantic salmon. In Effect of Ionizing Radiation on the Organism; Sorokin, B.P., Ed.; AEC-tr-7418; U.S. AEC: Washington, DC, USA, 1973; p. 7. [Google Scholar]
- Nelson, V.A. Effects of 90Sr plus 90Y, 65Zn and 51Cr on the larvae of the Pacific oyster, Crassostrea gigas. In The Columbia River Estuary and Adjacent Ocean Waters; Pruler, A.T., Alverson, D.L., Eds.; University of Washington Press: Seattle, WA, USA, 1972; p. 819. [Google Scholar]
- Tsytsugina, V.G. Effect of incorporated radionuclides on chromosome apparatus of ocean fish. In Marine Radioecology; Polikarpov, G.G., Ed.; AEC-tr-7299; US Atomic Energy Commission: Washington, DC, USA, 1972; pp. 157–165. [Google Scholar]
- Tsytsugina, V.G. Effect of incorporated radionuclides on the chromosomal apparatus of marine fish. In Effect of Ionizing Radiation on the Organism; Sorokin, B.P., Ed.; AEC-tr-7418; US Atomic Energy Commission: Washington, DC, USA, 1973; p. 157. [Google Scholar]
- Guthrie, J.E.; Brust, R.A. Sterilization of Aedes aegypti (L.) as a result of chronic exposure to low doses of beta radiation. Nature 1971, 232, 345. [Google Scholar] [CrossRef]
- Guthrie, J.E.; Brust, R.A. Effects of chronic radiation exposure on mosquitoes. I: Effects of rearing in 90Sr + 90Y solutions. In Radionuclides in Ecosystems, Proceedings of the 3rd National Symposium on Radioecology, 10–12 May 1971, Oak Ridge, TN, USA; Nelson, D.J., Ed.; CONF-710501; US Atomic Energy Commission: Washington, DC, USA, 1973; p. 1174. [Google Scholar]
- Scott, A.G. Radiation Dose to Mosquito Larvae Reared in Radioactive Media; Rep. AECL-3789; Atomic Energy of Canada Ltd.: Chalk River, ON, Canada, 1972. [Google Scholar]
- Strand, J.A.; Fujihara, M.P.; Templeton, W.L.; Tangen, F.G. Suppression of Chondrococcus columnaris immune response in rainbow trout sub-lethally exposed to tritiated water during embryogenesis. In Radioactive Contamination of the Marine Environment, Proceedings of an International Conference, 10–14 July 1972, Seattle, WA, USA; IAEA: Vienna, Austria, 1973; p. 543. [Google Scholar]
- Strand, J.A.; Templeton, W.L.; Tangen, F.G. Accumulation and retention of tritium (tritiated water) in embryonic and larval fish, and radiation effect. In Radionuclides in Ecosystems, Proceedings of the 3rd National Symposium on Radioecology, 10–12 May 1971, Oak Ridge, TN, USA; Nelson, D.J., Ed.; CONF-710501; US Atomic Energy Commission: Washington, DC, USA, 1973; p. 445. [Google Scholar]
- Williams, R.B.; Murdoch, M.B. Long-term effects of cesium-137 on a copepod. In Annual Report of Bureau of Commercial Fisheries Radiobiological Laboratory Beaufort, NC for the Fiscal Year Ending 30 June 1963; Circ. 204; Fisheries and Wildlife Services: Washington, DC, USA, 1965; p. 28. [Google Scholar]
- White, J.C.; Angelovic, J.W.; Engel, D.W.; Davis, E.M. Interactions of radiation, salinity and temperature on estuarine organisms. Effects on brine shrimp. In Annual Report of Bureau of Commercial Fisheries Radiobiological Laboratory Beaufort; Circ. 270; Fisheries and Wildlife Services: Washington, DC, USA, 1967; p. 33. [Google Scholar]
- Hallopeau, C. Recherches de l’action éventuelle de pollutions radioactives (Cs-137, mélange de produits de fission) sur le développement et la reproduction d ‘Artemia salina. In Rapports et Procès-Verbaux des Réunions Commission Internationale pour l’Exploration Scientifique de la Mer Méditerranée; International Commission for the Scientific Exploration of the Mediterranean Sea: Monte Carlo, Monaco, 1969; Volume 19, p. 961. [Google Scholar]
- Migalovskij, I.P. Development of Atlantic salmon eggs under conditions of radioactive contamination of water by strontium-90–yttrium-90 and cerium-144. In Effect of Ionizing Radiation on the Organism; Sorokin, B.P., Ed.; AEC-tr-7418; US Atomic Energy Commission: Washington, DC, USA, 1973; p. 16. [Google Scholar]
- Migalovskij, I.P. Development of fish eggs and the early period of gametogenesis in the embryos and larvae of the Atlantic salmon under conditions of radioactive contamination of water. In Effect of Ionizing Radiation on the Organism; AEC-tr-7418; Sorokin, B.P., Ed.; US Atomic Energy Commission: Washington, DC, USA, 1973; p. 36. [Google Scholar]
- Mashneva, N.J. Effect of complex mixtures of radioisotopes on developing eggs of freshwater fish after one-time contamination of water bodies. Gig. Sanit. 1971, 36, 118. (In Russian) [Google Scholar]
- Akita, Y.; Shiroya, T. Radiation effects on development of eggs of marine organisms. In Report on the Influence of Low Concentrations of Radionuclides in Water to Aquatic Organisms; Nuclear Safety Research Association of Japan: Tokyo, Japan, 1970. (In Japanese) [Google Scholar]
- Etoh, H.; Ueno, A.; Ichikawa, R. Influence of low concentrations of radionuclides in water to aquatic organisms. In Report on the Influence of Low Concentrations of Radionuclides in Water to Aquatic Organisms; Nuclear Safety Research Association of Japan: Tokyo, Japan, 1970. (In Japanese) [Google Scholar]
- Buggeln, R.G.; Held, E. Annual Report, Johnson Atoll Bioenvironmental Program, 1967–1969; Rep. NVO-269-2, Appendix F; US Atomic Energy Commission: Washington, DC, USA, 1968.
- Blaylock, B.G.; Ulrikson, G.V.; Rohwer, P.; Allen, C.P.; Griffth, N.A. Embryonic Development of Cyprinus carpio Eggs as a Biological Test. In Annual Progress Report for Period Ending 31 July 1970; Rep. No. ORNL-4634; Oak Ridge National Laboratory: Oak ridge, TN, USA, 1970; p. 9. [Google Scholar]
- Walden, S.J. Effects of tritiated water on the embryonic development of the three-spine stickleback Gasterosteus aculeatus L. In Radionuclides in Ecosystems, Proceedings of the 3rd National Symposium on Radioecology, 10–12 May 1971, Oak Ridge, TN, USA; Nelson, D.J., Ed.; CONF-710501; US Atomic Energy Commission: Washington, DC, USA, 1973; p. 1087. [Google Scholar]
- Erickson, R.C. Effects of chronic irradiation by tritiated water on Poecilia reticulata, the guppy. In Radionuclides in Ecosystems, Proceedings of the 3rd National Symposium on Radioecology, 10–12 May 1971, Oak Ridge, TN, USA; Nelson, D.J., Ed.; CONF-710501; US Atomic Energy Commission: Washington, DC, USA, 1973; p. 1091. [Google Scholar]
- Ichikawa, R.; Suyama, I. Effects of tritiated water on the embryonic development of marine fishes. Bull. Jap. Soc. Sei. Fish. 1974, 40, 819. [Google Scholar] [CrossRef] [Green Version]
- Fedorova, G.V. The biological effect of 14C on the fish in early development stages. Vopr. Ichtiol. 1972, 12, 198. (In Russian) [Google Scholar]
- Povinec, P.P.; Hirose, K.; Aoyama, M. Radiostrontium in the western north Pacific: Characteristics, behavior and the Fukushima impact. Environ. Sci. Technol. 2012, 46, 10356–10363. [Google Scholar] [CrossRef] [PubMed]
- Buesseler, K.; Aoyama, M.; Fukasawa, M. Impacts of the Fukushima nuclear power plants on marine radioactivity. Environ. Sci. Technol. 2011, 45, 9931–9935. [Google Scholar] [CrossRef]
- Aoyama, M.; Tsumune, D.; Uematsu, M.; Kondo, F.; Harajima, Y. Temporal variation of 134Cs and 137Cs activities in surface water at stations along the coastline near the Fukushima Dai-ichi Nuclear Power Plant accident site, Japan. Geochem. J. 2012, 46, 321–325. [Google Scholar] [CrossRef] [Green Version]
- Nuclear Regulation Authority, Japan. Temporal Trends on Concentrations of Radionuclides in Seawater at Sampling Points Adjacent to the Fukushima Daiichi Nuclear Power Plant. 2020. Available online: https://radioactivity.nsr.go.jp/ja/contents/9000/8141/24/engan.pdf (accessed on 2 March 2022). (In Japanese)
- Kanda, J. Deposition, discharge, spreading/dispersion and accumulation of radionuclides to the ocean In Contamination by Radionuclides in the Aquatic Environment; Kurokura, H., Ed.; Koseisha-Koseikaku: Tokyo, Japan, 2014; pp. 27–53. (In Japanese) [Google Scholar]
- Miyazawa, Y.; Masumoto, Y.; Varlamov, S.M.; Miyama, T.; Takigawa, M.; Honda, M.; Saino, T. Inverse estimation of source parameters of oceanic radioactivity dispersion models associated with the Fukushima accident. Biogeosciences 2013, 10, 2349–2363. [Google Scholar] [CrossRef] [Green Version]
- International Commission on Radiological Protection. Environmental Protection—The Concept and Use of Reference Animals and Plants; ICRP Publication 108. Ann. ICRP 38 (4–6); ICRP: Ottawa, ON, Canada, 2008. [Google Scholar]
- Vandenhove, H. Effects of Ionizing Radiation on Non-Human Biota. 2011. Available online: http://www.bvsabr.be/js/tinymce/plugins/moxiemanager/data/files/20110415/Vandenhove.pdf (accessed on 15 March 2022).
- United Nations Scientific Committee on the Effects of Atomic Radiation. Sources and Effects of Ionizing Radiation: UNSCEAR Report to the General Assembly, with Scientific Annex: Effects of Radiation on the Environment; United Nations: New York, NY, USA, 1996; Available online: https://www.unscear.org/docs/publications/1996/UNSCEAR_1996_Report.pdf (accessed on 23 March 2022).
- United Nations Scientific Committee on the Effects of Atomic Radiation. Sources and Effects of Ionizing Radiation: UNSCEAR Report to the General Assembly, with Scientific Annexes, Annex E: Effects of Ionizing Radiation on Non-Human Biota; United Nations: New York, NY, USA, 2008; Available online: http://www.unscear.org/docs/reports/2008/11-80076_Report_2008_Annex_E.pdf (accessed on 23 March 2022).
- United Nations Scientific Committee on the Effects of Atomic Radiation. Sources, Effects and Risks of Ionizing Radiation. Volume II: Scientific Annex B; United Nations: New York, NY, USA, 2020; Available online: https://www.unscear.org/docs/publications/2020/UNSCEAR_2020_21_Report_Vol.II.pdf (accessed on 23 March 2022).
- Adam-Guillermin, C.; Pereira, S.; Della-Vedova, C.; Hinton, T.; Garnier-Laplace, J. Genotoxic and reprotoxic effects of tritium and external gamma irradiation on aquatic animals. In Reviews of Environmental Contamination and Toxicology; Whitacre, D.M., Ed.; Springer: New York, NY, USA, 2012; pp. 67–103, Softcover; ISBN 978-1-4939-0136-4. [Google Scholar] [CrossRef]
- Feinendegen, L.E.; Bond, V.P.; Sondhaus, C.A. Relative biological effectiveness of ionizing radiations determined in tissue (RBE) fails in assessing comparative relative effectiveness in the tissue cells. Health Phys. 1996, 70, 852–858. [Google Scholar] [CrossRef]
- Bradshaw, C.; Kapustka, L.; Barnthouse, L.; Brown, J.; Ciffroy, P.; Forbes, V.; Geras’kin, S.; Kautsky, U.; Bréchignac, F. Using an Ecosystem Approach to complement protection schemes based on organism-level endpoints. J. Environ. Radioact. 2014, 136, 98–104. [Google Scholar] [CrossRef] [Green Version]
- International Union of Radioecology. IUR Report No. 7: Towards an Ecosystem Approach for Environment Protection with Emphasis on Radiological Hazards, 2nd ed.; International Union of Radioecology: Saint-Paul-lez-Durance, France, 2012; ISBN 978-0-9554994-4-9. Available online: https://www.iur-uir.org/en/publications/others-publications/id-15-iur-report-7-towards-an-ecosystem-approach-for-environment-protection-with-emphasis-on-radiological-hazards (accessed on 2 March 2022).
- Imanaka, T. Chapter 20, Comparison of radioactivity release and contamination from the Fukushima and Chernobyl Nuclear Power Plant accidents. In Low-Dose Radiation Effects on Animals and Ecosystems, Long-Term Study on the Fukushima Nuclear Accident; Fukumoto, M., Ed.; Springer: Tokyo, Japan, 2019; pp. 249–259. [Google Scholar]
- Buesseler, K.; Dai, M.; Aoyama, M.; Benitez-Nelson, C.; Charmasson, S.; Higley, K.; Maderich, V.; Masqué, P.; Morris, P.J.; Oughton, D.; et al. Fukushima Daiichi-derived radionuclides in the ocean: Transport, fate, and impacts. Annu. Rev. Mar. Sci. 2017, 9, 173–203. [Google Scholar] [CrossRef]
- New York State Department of Environmental Conservation Division of Fish, Wildlife and Marine Resources Bureau of Habitat. Assessment of the Risks to Fish and Wildlife from Exposure to Ionizing Radiation. 2014. Available online: https://www.dec.ny.gov/docs/fish_marine_pdf/radriskfw.pdf (accessed on 2 March 2022).
- Urabe, J.; Suzuki, T.; Nishita, T.; Makino, W. Correction: Immediate ecological impacts of the 2011 Tohoku Earthquake Tsunami on intertidal flat communities. PLoS ONE 2013, 8, 10. [Google Scholar] [CrossRef]
- Noda, T.; Iwasaki, A.; Fukaya, K. Recovery of rocky intertidal zonation: Two years after the 2011 Great East Japan Earthquake. J. Mar. Biol. Assoc. UK 2016, 96, 1549–1555. [Google Scholar] [CrossRef]
- Buhay, J.E. “Population dynamics of crustaceans”: Introduction to the symposium. Integr. Comp. Biol. 2011, 51, 577–579. [Google Scholar] [CrossRef] [Green Version]
- Kodama, K.; Horiguchi, T. Changes in the megabenthic community structure in relation to environmental variations in Tokyo Bay, Japan. In Advances in Environmental Research; Daniels, J.A., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2012; Volume 20, pp. 125–146. [Google Scholar]
- Trowbridge, C.D.; Little, C.; Plowman, C.Q.; Williams, G.A.; Pilling, G.M.; Morritt, D.; Rivera Vázquez, Y.; Dlouhy-Massengale, B.; Cottrell, D.M.; Stirling, P.; et al. No ‘silver bullet’: Multiple factors control population dynamics of European purple sea urchins in Lough Hyne Marine Reserve, Ireland. Estuar. Coast. Shelf Sci. 2019, 226, 106271. [Google Scholar] [CrossRef]
- Lee, J.H.; Kodama, K.; Shiraishi, H.; Horiguchi, T. Effect of water temperature on survival of early-life stages of marbled flounder Pseudopleuronectes yokohamae in Tokyo Bay, Japan. Mar. Environ. Res. 2017, 128, 107–113. [Google Scholar] [CrossRef]
- Kodama, K.; Tajima, Y.; Shimizu, T.; Ohata, S.; Shiraishi, H.; Horiguchi, T. Disturbance of recruitment success of mantis shrimp in Tokyo Bay associated with effects of hypoxia on the early life history. Mar. Pollut. Bull. 2014, 85, 433–438. [Google Scholar] [CrossRef]
- Kodama, K.; Lee, J.H.; Shiraishi, H.; Horiguchi, T. Changes in growth and reproductive traits of dragonet Callionymus valenciennei in Tokyo Bay, concurrent with decrease in stock size. Mar. Environ. Res. 2014, 93, 23–30. [Google Scholar] [CrossRef]
- Horiguchi, T.; Kojima, M.; Hamada, F.; Kajikawa, A.; Shiraishi, H.; Morita, M.; Shimizu, M. Impact of tributyltin and triphenyltin on ivory shell (Babylonia japonica) populations. Environ. Health Perspectives 2006, 114 (Suppl. 1), 13–19. [Google Scholar] [CrossRef] [PubMed]
- Hamada, F.; Kanazawa, T.; Yamamoto, E. Seed production of the ivory shell, Babylonia japonica Reeve. Bull. Tottori Prefect Fish Exp. Station 1988, 6, 110–116. (In Japanese) [Google Scholar]
- Hamada, F.; Kanazawa, T.; Yamamoto, E. Seed production of the ivory shell, Babylonia japonica Reeve. Bull. Tottori Prefect Fish Exp. Station 1989, 7, 103–109. (In Japanese) [Google Scholar]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef] [Green Version]
- Hyodo, Y. Effect of X-irradiation on the intestinal epithelium of the goldfish Carassius auratus. II. Influence of temperature on the development of histopathological changes in the intestine. Radiat. Res. 1965, 24, 133. [Google Scholar] [CrossRef]
- Hyodo, Y. Development of intestinal damage after X-irradiation and 3H-thymidine into testinal epithelial cells of irradiated goldfish, Carassius auratus, at different temperatures. Radiat. Res. 1965, 26, 383. [Google Scholar] [CrossRef]
- Iwasaki, T. Sensitivity of Artemia eggs to the y (gamma)-irradiation. V. Biological after-effect of irradiation in relation to water content and temperature. Radiat. Res. 1965, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki, T. Sensitivity of Artemia eggs to the y (gamma)-irradiation. VII. Relationship between the degree of biological damage and the decay of free radicals in irradiated eggs. Int. J. Radiat. Biol. 1965, 9, 573. [Google Scholar]
- Angelovic, J.W.; White, J.C.; Engel, D.W. Influence of salinity on the response of estuarine animals to ionizing radiation. Assoc. Southeastern Biol. Bull. 1966, 13, 29. [Google Scholar]
- Angelovic, J.W.; White, J.C.; Davis, E.M. Interactions of ionizing radiation, salinity and temperature on the estuarine fish, Fundulus heteroclitus. In Proceedings of the 3rd National Symposium on Radioecology, CONF-670503, Oak Ridge, TN, USA, 10–12 May 1971; Nelson, D.J., Evans, F.C., Eds.; US Atomic Energy Commission: Washington, DC, USA, 1969; p. 131. [Google Scholar]
- Conte, F.P. Effects of ionizing radiation on osmoregulation in fish Oncorhynchus kisutch. Comp. Biochem. Physiol. 1965, 15, 293. [Google Scholar] [CrossRef]
- Vives i Batlle, J.; Aono, T.; Brown, J.E.; Hosseini, A.; Garnier-Laplace, J.; Sazykina, T.; Steenhuisen, F.; Strand, P. The impact of the Fukushima nuclear accident on marine biota: Retrospective assessment of the first year and perspectives. Sci. Total Environ. 2014, 487, 143–153. [Google Scholar] [CrossRef]
- Bréchignac, F.; Bradshaw, C.; Carroll, S.; Jaworska, A.; Kapustka, L.; Monte, L.; Oughton, D. Recommendations from the International Union of Radioecology to Improve Guidance on Radiation Protection. Integr. Environ. Assess. Manag. 2011, 7, 411–413. [Google Scholar] [CrossRef]
- Nakano, D.; Nagoshi, M. Growth and age of Thais clavigera (Küster), Prosobranch, in tidal zone around Shima Peninsula, Japan. In the 25th Anniversary Memorial Publication of Toba Aquarium; Toba Aquarium: Toba, Japan, 1980; pp. 87–92. (In Japanese) [Google Scholar]


| Group | (Krad) | (Gy) | Remarks # |
|---|---|---|---|
| Bacteria | 4.5–735 | 45–7350 | LD90 |
| Cyanobacteria | <400 to >1200 | <4000 to >12000 | LD90 |
| Other algae | 3–120 | 30–1200 | LD50 |
| Protozoa | <600 | <6000 | LD50 |
| Mollusks | 20–109 | 200–1090 | LD50/30 |
| Crustaceans | 1.5–56.6 | 15–566 | LD50/30 |
| Fishes | 1.1–5.6 | 11–56 | LD50/30 |






{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horiguchi, T.; Kodama, K. What Caused Declines in Intertidal Invertebrate Populations around Fukushima Daiichi Nuclear Power Plant after the 2011 Great East Japan Earthquake, Tsunami, and Nuclear Disaster? Toxics 2022, 10, 214. https://doi.org/10.3390/toxics10050214
Horiguchi T, Kodama K. What Caused Declines in Intertidal Invertebrate Populations around Fukushima Daiichi Nuclear Power Plant after the 2011 Great East Japan Earthquake, Tsunami, and Nuclear Disaster? Toxics. 2022; 10(5):214. https://doi.org/10.3390/toxics10050214
Chicago/Turabian StyleHoriguchi, Toshihiro, and Keita Kodama. 2022. "What Caused Declines in Intertidal Invertebrate Populations around Fukushima Daiichi Nuclear Power Plant after the 2011 Great East Japan Earthquake, Tsunami, and Nuclear Disaster?" Toxics 10, no. 5: 214. https://doi.org/10.3390/toxics10050214
APA StyleHoriguchi, T., & Kodama, K. (2022). What Caused Declines in Intertidal Invertebrate Populations around Fukushima Daiichi Nuclear Power Plant after the 2011 Great East Japan Earthquake, Tsunami, and Nuclear Disaster? Toxics, 10(5), 214. https://doi.org/10.3390/toxics10050214