
Oxidative Stress and Histopathological Changes in Gills and Kidneys of Cyprinus carpio following Exposure to Benzethonium Chloride, a Cationic Surfactant

 ,
,  ,
,  ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Fish
2.3. Sub-Lethal Toxicity Experiment
2.4. Analytical Control of BEC
2.5. Biochemical Investigations
2.5.1. Obtaining the Tissue Homogenate
2.5.2. Measurement of Enzyme Activities
2.5.3. Measurement of GSH Level
2.5.4. Measurement of Lipid Peroxidation
2.5.5. Statistical Analysis
2.6. Histological Staining
3. Results
3.1. Characterization of Water Parameters
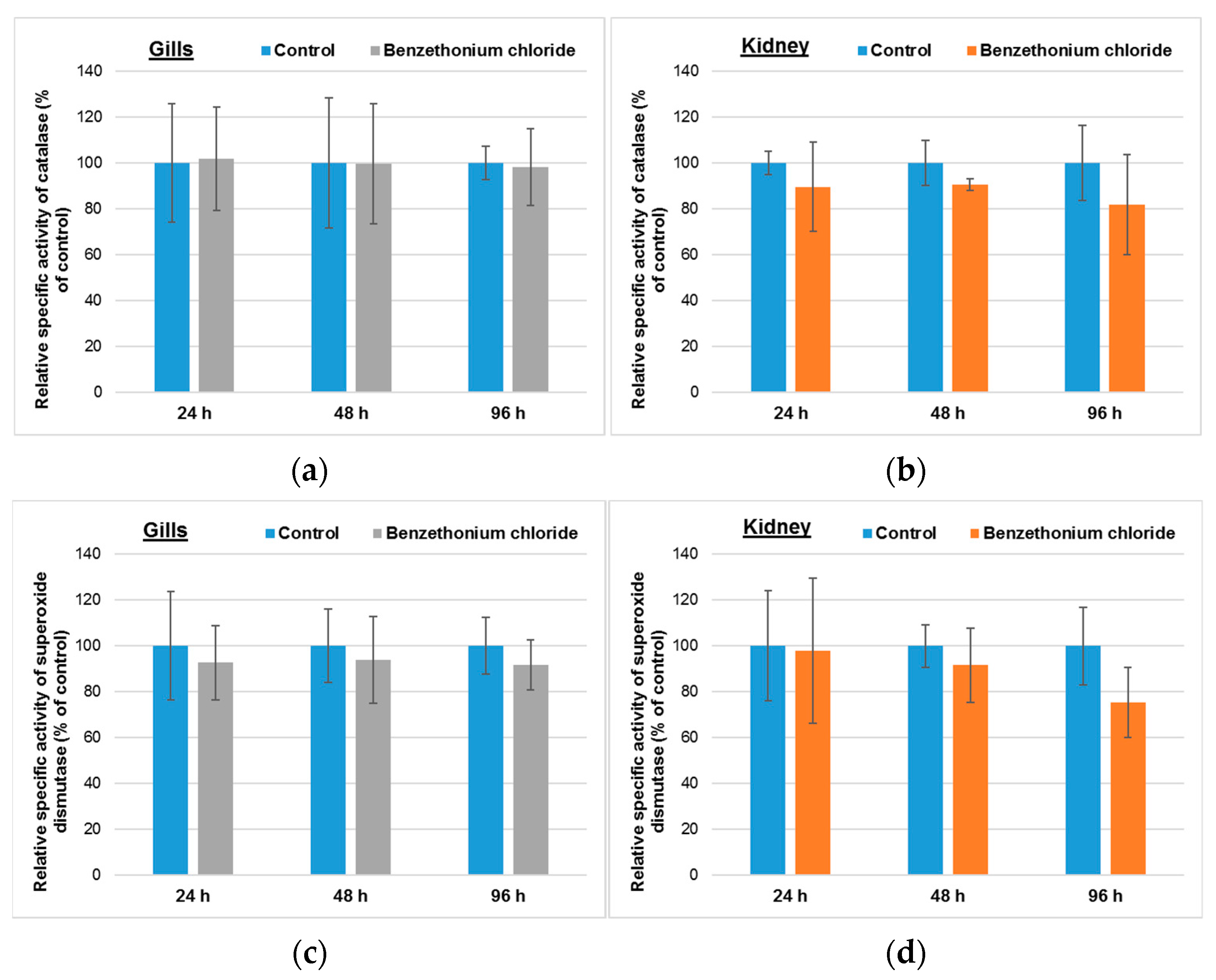
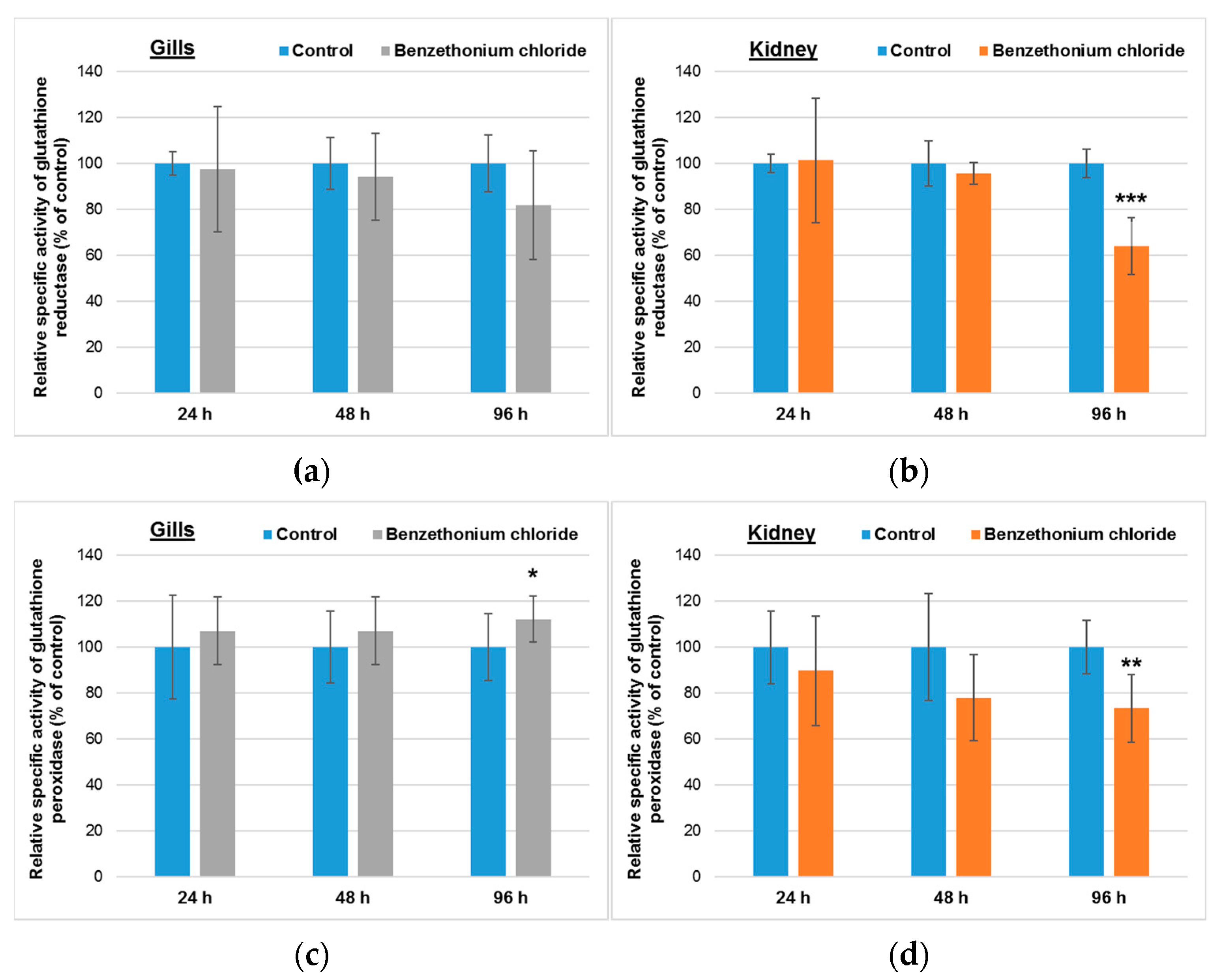
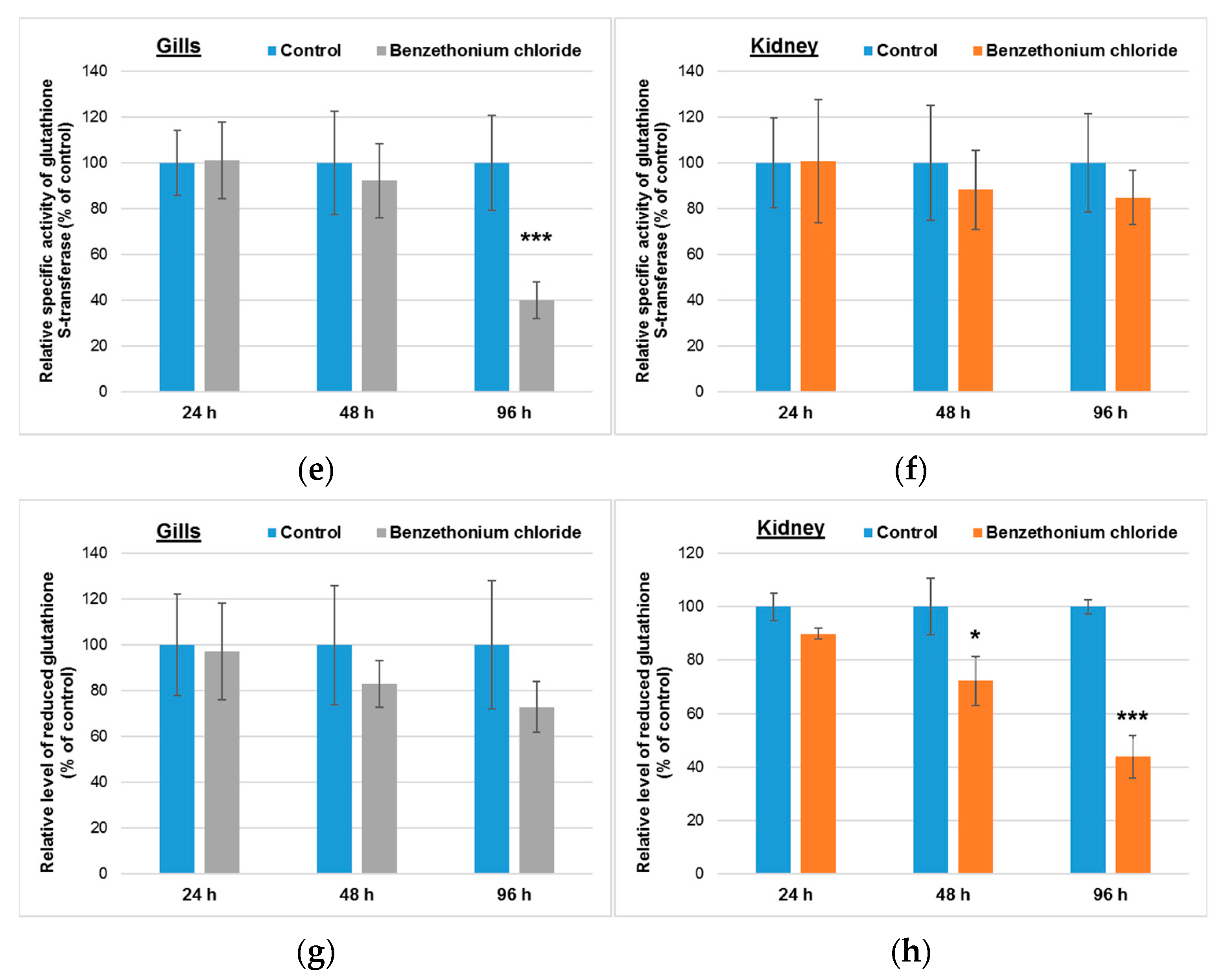
3.2. Effects Induced by BEC on Cellular Antioxidant System
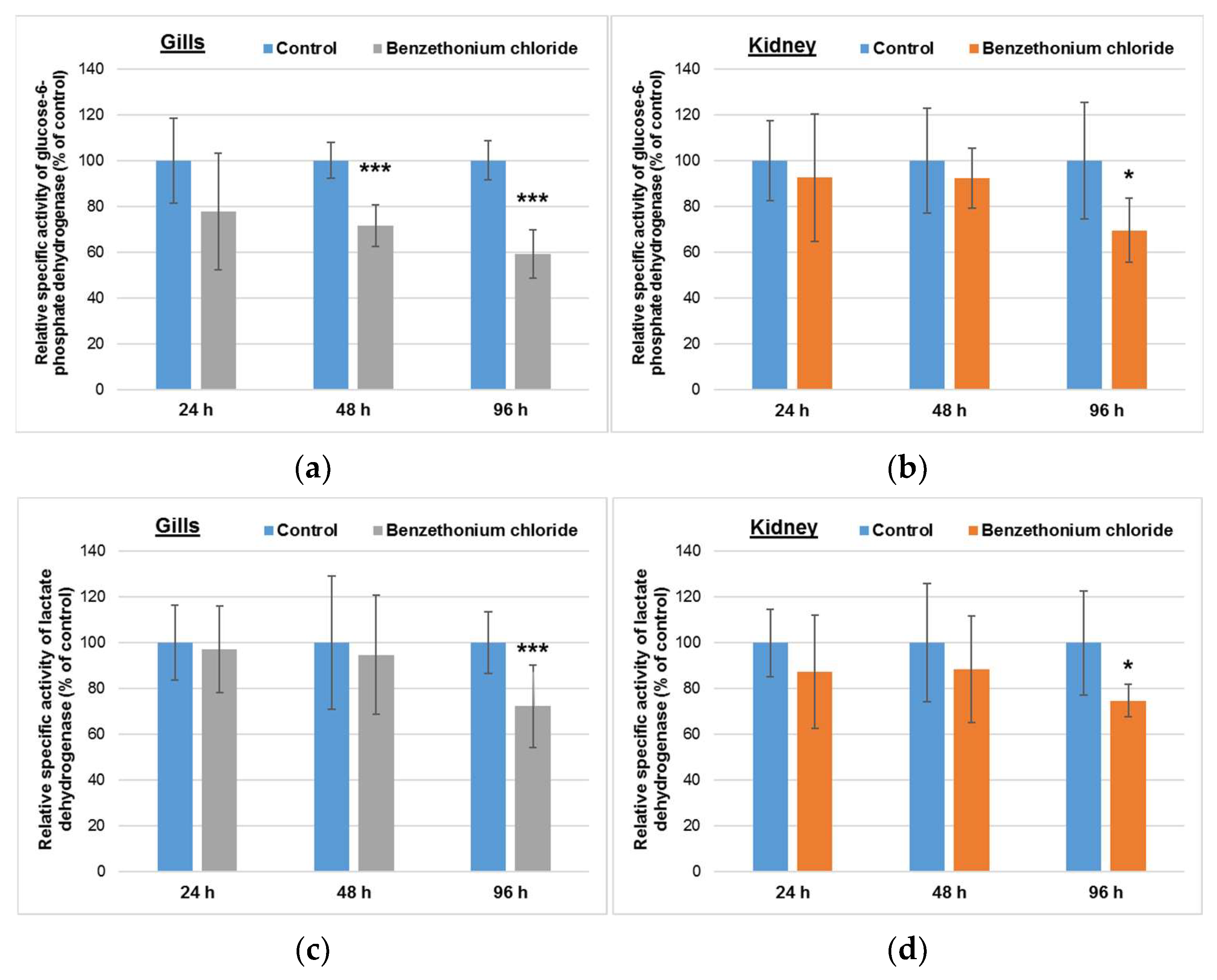
3.3. Effects Induced by BEC on Enzymes-Generating Reducing Equivalents
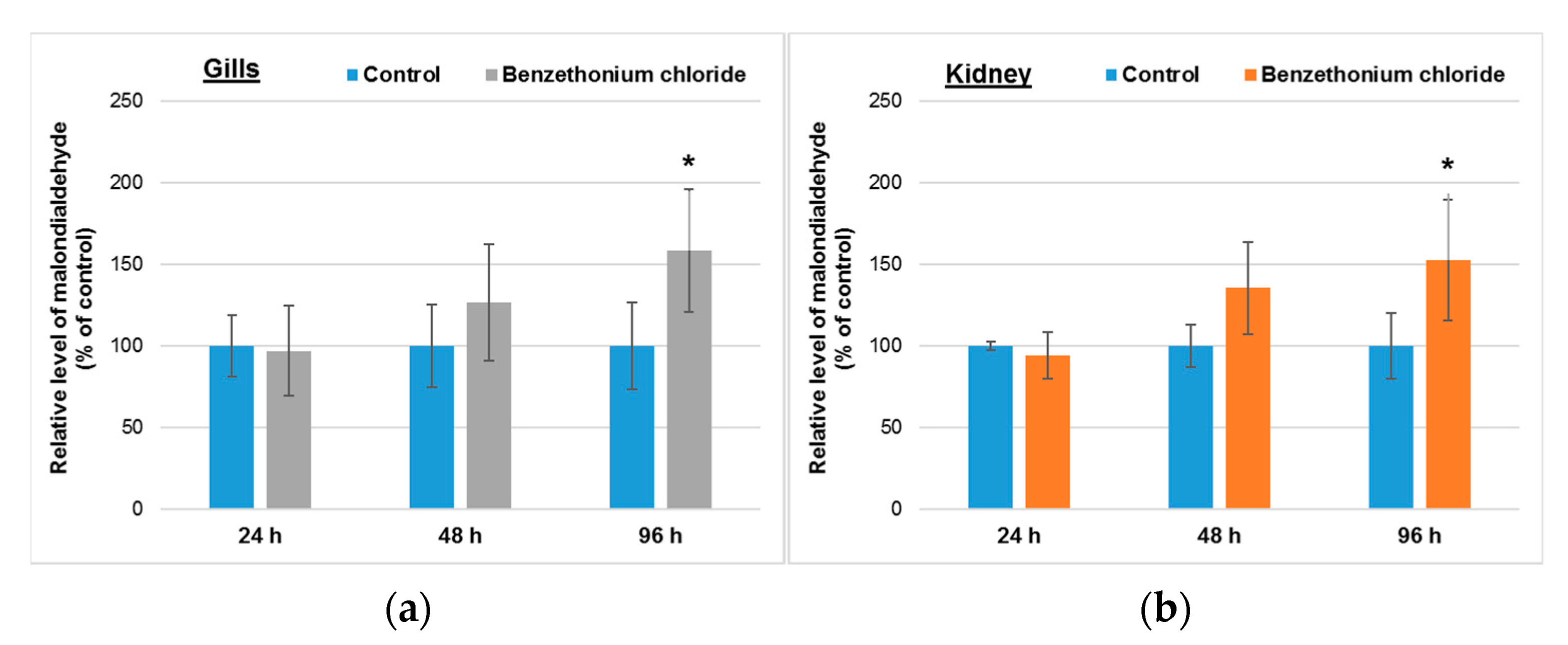
3.4. Effects Induced by BEC on Lipid Peroxidation
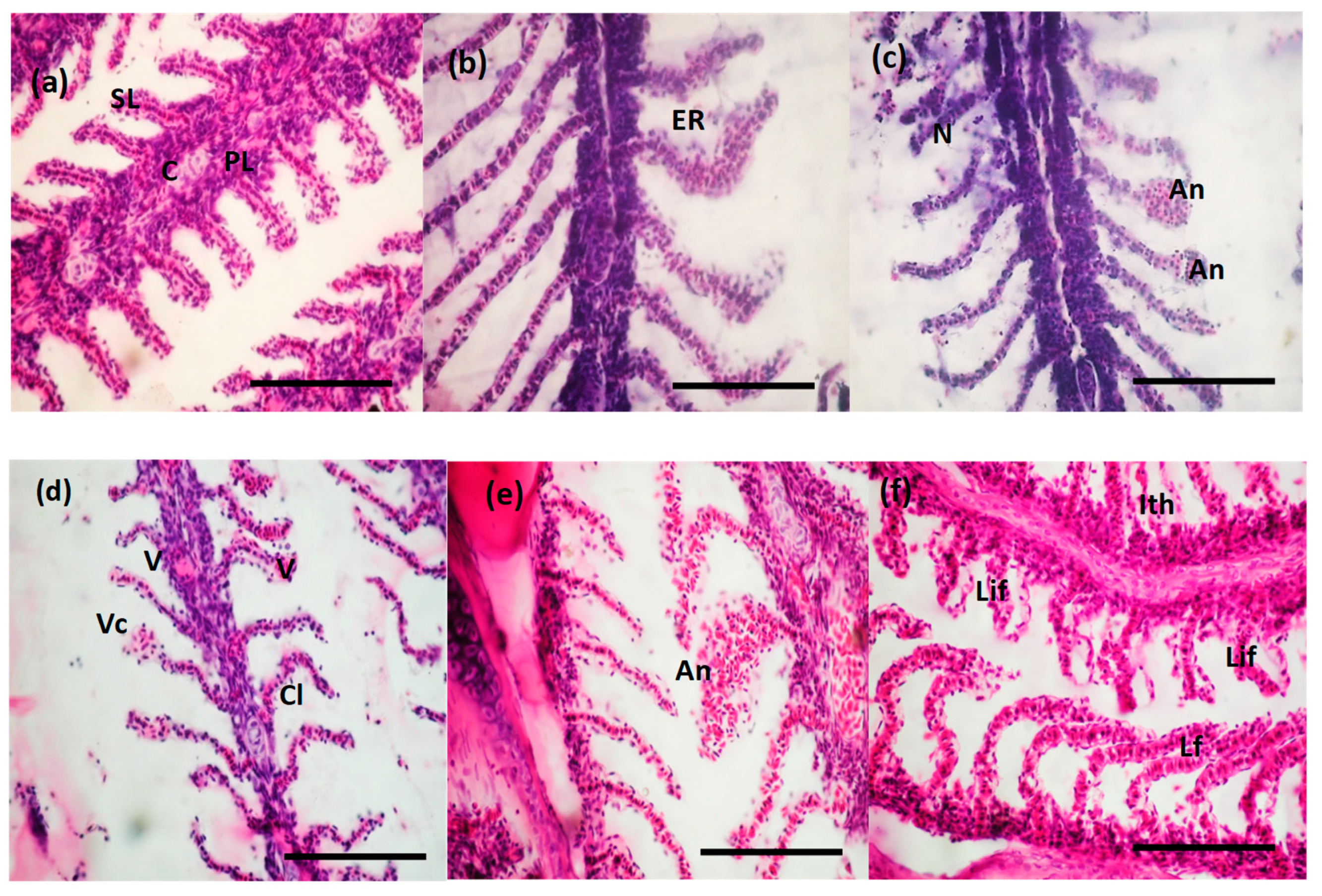
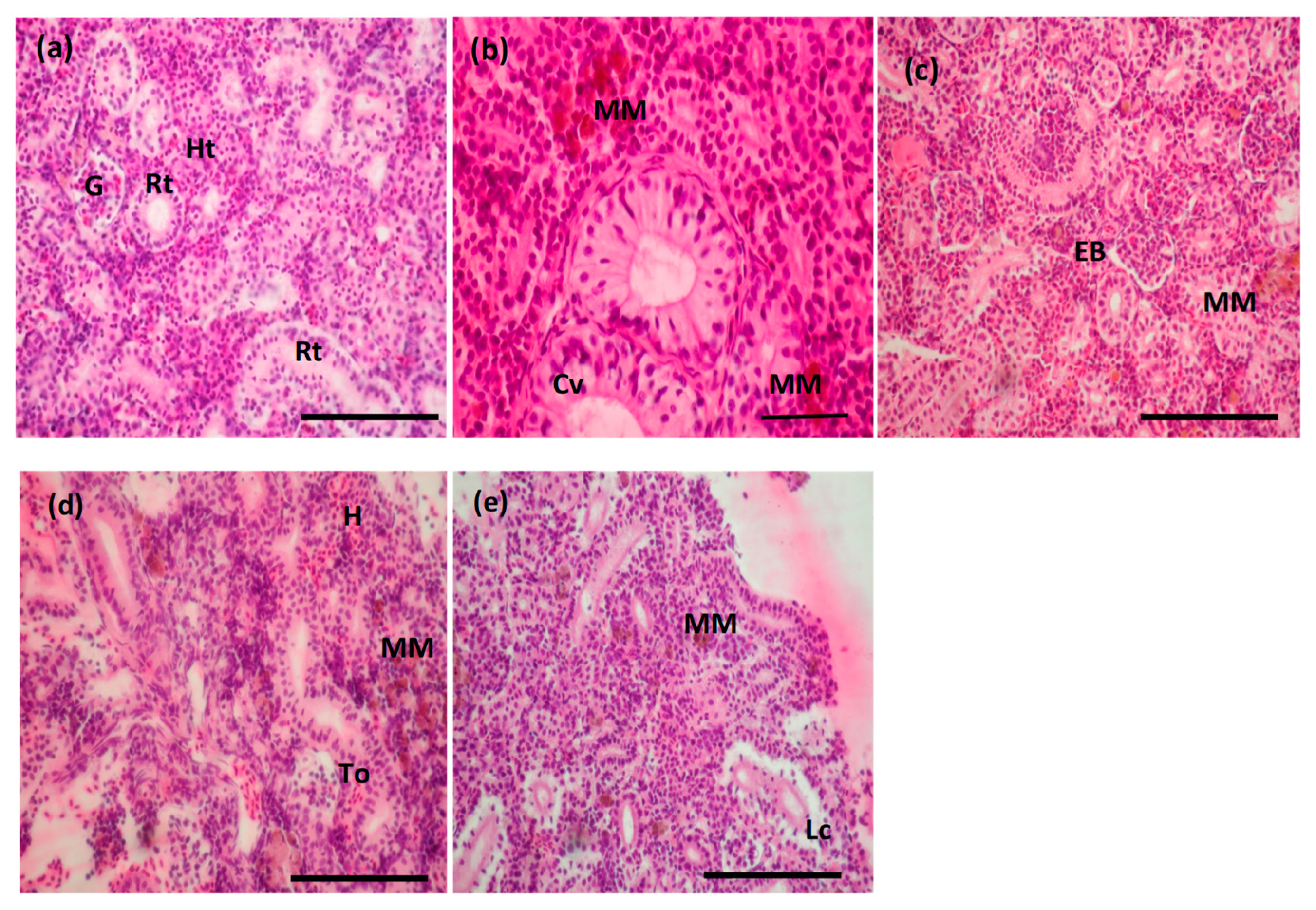
3.5. Histopathological Changes Induced by BEC
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ivancovic, T.; Hrenovic, J. Surfactants in the environment. Arh. Hig. Rada Toksikol. 2010, 61, 95–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradhan, A.; Bhattacharyya, A. Quest for an eco-friendly alternative surfactant: Surface and foam characteristics of natural surfactants. J. Clean Prod. 2017, 150, 127–134. [Google Scholar] [CrossRef]
- Brunelli, E.; Talarico, E.; Corapi, B.; Perrotta, I.; Tripepi, S. Effects of a sublethal concentration of sodium lauryl sulphate on the morphology and Na+/K+ ATPase activity in the gill of the ornate wrasse (Thalassoma pavo). Ecotoxicol. Environ. Saf. 2008, 71, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Muñoz, D.; Gómez-Parra, A.; Blasco, J.; Sarasquete, C.; González-Mazo, E. Oxidative stress and histopathology damage related to the metabolism of dodecylbenzene sulfonate in Senegalese sole. Chemosphere 2009, 74, 1216–1223. [Google Scholar] [CrossRef]
- Mandal, R.; Mandal, D.; Mishra, N.; Bahadur, A. Effect of surfactants on phosphatase level of fresh water fish Labeo rohita. J. Environ. Biol. 2010, 31, 395–398. [Google Scholar] [PubMed]
- Mustapha, D.S.; Bawa-Allah, K.A. Differential toxicities of anionic and nonionic surfactants in fish. Environ. Sci. Pollut. Res. 2020, 27, 16754–16762. [Google Scholar] [CrossRef]
- Dani, U.; Bahadur, A.; Kuperkar, K. Biotoxicity and tissue-specific oxidative stress induced by Gemini surfactant as a protocol on fingerlings of Cirrhinus mrigala (Ham.): An integrated experimental and theoretical methodology. Ecotoxicol. Environ. Saf. 2019, 183, 1–9. [Google Scholar] [CrossRef]
- Johnson, P.; Trybala, A.; Starov, V.; Pinfield, V.J. Effect of synthetic surfactants on the environment and the potential for substitution by biosurfactants. Adv. Colloid Interface Sci. 2021, 288, 102340. [Google Scholar] [CrossRef]
- DeLeo, P.C.; Huynh, C.; Pattanayek, M.; Clark Schmid, K.; Pechacek, N. Assessment of ecological hazards and environmental fate of disinfectant quaternary ammonium compounds. Ecotoxicol. Environ. Saf. 2020, 206, 111116. [Google Scholar] [CrossRef]
- Agrawal, K.; Agnihotri, G.; Shrivas, K.; Mundhara, G.L.; Patel, K.S.; Hofmann, P. Determination of cationic surfactants in environmental samples by flow injection analysis. Microchim. Acta 2004, 147, 273–278. [Google Scholar] [CrossRef]
- Zhang, C.; Cui, F.; Zeng, G.M.; Jiang, M.; Yang, Z.Z.; Yu, Z.G.; Zhu, M.Y.; Shen, L.Q. Quaternary ammonium compounds (QACs): A review on occurrence, fate and toxicity in the environment. Sci. Total Environ. 2015, 15, 518–519. [Google Scholar] [CrossRef] [PubMed]
- National Industrial Chemicals Notification and Assessment Scheme (NICNAS); Full Public Report, Stepantex Esterquat, File No: STD/1027; National Occupational Health and Safety Commission: Canberra, Australia, 2003.
- Gheorghe, S.; Lucaciu, I.; Paun, I.; Stoica, C.; Stanescu, E. Environmental exposure and effects of some micropollutants found in the Romanian surface waters. J. Environ. Prot. Ecol. 2014, 15, 878–888. [Google Scholar]
- Gheorghe, S.; Lucaciu, I.; Paun, I.; Stoica, C.; Stanescu, E. Ecotoxicological behavior of some cationic and amphoteric surfactants (biodegradation, toxicity and risk assessment). In Biodegradation—Life Science; Chamy, R., Rosenkranz, F., Eds.; IntechOpen: London, UK, 2013; pp. 83–114. [Google Scholar]
- Reregistration Eligibility Decision (RED) for Alkyl Dimethyl Benzyl Ammonium Chloride (ADBAC); EPA739-R-06-009; U.S. Environmental Protection Agency (USEPA): Washington, DC, USA, 2006.
- Compendium, Containing Abstracts, Discussions, and Conclusions of CIR Cosmetic Ingredient Safety Assessments; Cosmetic Ingredient Review: Washington, DC, USA, 2006.
- Clara, M.; Scharf, S.; Sche_knecht, C.; Gans, O. Occurrence of selected surfactants in untreated and treated sewage. Water Res. 2007, 41, 4339–4348. [Google Scholar] [CrossRef]
- Czerwinski, S.E.; Cozean, J.; Cozean, C. Novel water-based antiseptic lotion demonstrates rapid, broad-spectrum kill compared with alcohol antiseptic. J. Infect. Public Health 2014, 7, 199–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Registration, Evaluation, Authorization and Restriction of Chemicals (REACH); Regulation no. 1907/2006; European Agency for Safety and Health at Work: Bilbao, Spain, 2022.
- Bindu, P.C.; Babu, P. Surfactant-induced lipid peroxidation in a tropical euryhaline teleost Oreochromis mossambicus (Tilapia) adapted to fresh water. Indian J. Exp. Biol. 2001, 39, 1118–1122. [Google Scholar]
- Nakata, K.; Tsuchido, T.; Matsumura, Y. Antimicrobial cationic surfactant, cetyltrimethylammonium bromide, induces superoxide stress in Escherichia coli cells. J. Appl. Microbiol. 2010, 110, 568–579. [Google Scholar] [CrossRef]
- Gheorghe, S.; Mitroi, D.N.; Stan, M.S.; Staicu, C.A.; Cicirma, M.; Lucaciu, I.E.; Nita-Lazar, M.; Dinischiotu, A. Evaluation of sub-lethal toxicity of benzethonium chloride in Cyprinus carpio liver. Appl. Sci. 2020, 10, 8485. [Google Scholar] [CrossRef]
- Abel, P.D. Toxicity of synthetic detergents to fish and aquatic invertebrates. J. Fish Biol. 1974, 6, 279–298. [Google Scholar] [CrossRef]
- Rosen, M.; Li, F.; Morrall, S.W.; Versteeg, D.J. The relationship between the interfacial properties of surfactants and their toxicity to aquatic organisms. Environ. Sci. Technol. 2001, 35, 954–959. [Google Scholar] [CrossRef]
- Svobodová, Z.; Lloyd, R.; Máchová, J.; Vykusová, B. Water Quality and Fish Health; EIFAC Technical Paper. No. 54; FAO: Rome, Italy, 1993. [Google Scholar]
- Livingstone, D.R. Oxidative stress in aquatic organisms in relation to pollution and aquaculture. Revue. Med. Vet. 2003, 154, 427–430. [Google Scholar]
- Chowdhury, M.J.; Blust, R. A mechanistic model for the uptake of waterborne strontium in the common carp (Cyprinus carpio L.). Environ. Sci. Technol. 2001, 35, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Sharples, F.; Anestidou, L.; Beil, K.; Fletcher, C.H.; Haycraft, R.; Zurlo, J. Guide for the Use and Care of Laboratory Animals. In National Research Council (US) Committee or the Update of the Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Fish Toxicity Testing Framework. In Environment Directorate Joint Meeting of The Chemicals Committee and The Working Party on Chemicals, Pesticides and Biotechnology, Organisation for Economic Co-operation and Development; Series on Testing and Assessment no. 171, ENV/JM/MONO (2012)16; OECD Publishing: Paris, France, 2012.
- Section 2—Effects on biotic Systems, Organisation for Economic Co-operation and Development—Guideline for the testing of chemicals. In Fish, Acute Toxicity Testing; Test Guideline No. 203; OECD Publishing: Paris, France, 2019.
- Lowry, O.H.; Rosenbrough, N.J.; Farr, A.L.; Randall, B.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Aebi, H. Catalase. In Methods of Enzymatic Analysis; Bergmeyer, H.V., Ed.; Academic Press: New York, NY, USA, 1984; pp. 673–677. [Google Scholar]
- Paoletti, F.; Mocali, A. Determination of superoxide dismutase activity by purely chemical system based on NADP(H) oxidation. Methods Enzymol. 1990, 186, 209–221. [Google Scholar] [PubMed]
- Beutler, E. Red Cell Metabolism: A Manual of Biochemical Method, 3rd ed.; Grune and Stratton: Orlando, FL, USA, 1984; pp. 68–73. [Google Scholar]
- Goldberg, D.M.; Spooner, R.J. Glutathione reductase. In Methods of Enzymatic Analysis, 3rd ed.; Bergmeyer, H.V., Ed.; Verlag Chemie: Weinheim, Germany, 1983; pp. 258–265. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Stanca, L.; Petrache, S.N.; Serban, A.I.; Staicu, A.C.; Sima, C.; Munteanu, M.C.; Zarnescu, O.; Dinu, D.; Dinischiotu, A. Interaction of silicon-based quantum dots with gibel carp liver: Oxidative and structural modifications. Nanoscale Res. Lett. 2013, 8, 254. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Moneim, A.M.; Al-Kahtani, M.A.; Elmenshawy, O.M. Histopathological biomarkers in gills and liver of Oreochromisniloticus from polluted wetland environments, Saudi Arabia. Chemosphere 2012, 88, 1028–1035. [Google Scholar] [CrossRef]
- Mishra, A.K.; Mohanty, B. Acute toxicity impacts of hexavalent chromium on behavior and histopathology of gill, kidney and liver of the fresh water fish, Channa punctatus (Bloch). Environ. Toxicol. Pharmacol. 2008, 26, 136–141. [Google Scholar] [CrossRef]
- Sweidan, A.H.; El-Bendary, N.; Hegazy, O.M.; Hassanien, A.E.; Snasel, V. Water pollution detection system based on fish gills as a biomarker. Procedia Comput. Sci. 2015, 65, 601–611. [Google Scholar] [CrossRef] [Green Version]
- Kierkegaard, A.; Chen, C.; Armitage, J.M.; Arnot, J.A.; Droge, S.; McLachlan, M.S. Tissue distribution of several series of cationic surfactants in rainbow trout (Oncorhynchus mykiss) following exposure via water. Environ. Sci. Technol. 2020, 54, 4190–4199. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, J.B.; De Canales, M.L.G.; Sarasquete, C. Histopathological changes induced by lindane (-HCH) in various organs of fishes. Sci. Mar. 2003, 67, 53–61. [Google Scholar]
- Droge, S.T.J.; Armitage, J.M.; Amot, J.A.; Fitzsimmons, P.N.; Nichols, J.W. Biotransformation Potential of Cationic Surfactants in Fish Assessed with Rainbow Trout Liver S9 Fractions. Env. Toxicol. Chem. 2021, 40, 3123–3136. [Google Scholar] [CrossRef] [PubMed]
- Hegstad, K.; Langsrud, S.; Lunestad, B.T.; Scheie, A.A.; Sunde, M.; Yazdankhah, S.P. Does the wide use of quaternary ammonium compounds enhance the selection and spread of antimicrobial resistance and thus threaten our health? Microb. Drug Resist. 2010, 16, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Inácio, A.S.; Costa, G.N.; Domingues, N.S.; Santos, M.S.; Moreno, A.J.M.; Vaz, W.L.C.; Vieira, O.V. Mitochondrial dysfunction is the focus of quaternary ammonium surfactant toxicity to mammalian epithelial cells. Antimicrob. Agents Chemother. 2014, 57, 2631–2639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tierney, K.B.; Kennedy, C.J.; Gobas, F.; Gledhill, M.; Sekela, M. Organic contaminants and fish. In Organic Chemical Toxicology of Fishes; Tierney, K., Farrell, A.P., Brauner, C.J., Eds.; Elsevier: Heidelberg, Germany, 2014; pp. 1–52. [Google Scholar]
- Sheng, Y.; Gralla, E.B.; Schumacher, M.; Cascio, D.; Cabelli, D.E.; Valentine, J.S. Six-coordinate manganese (3+) in catalysis by yeast manganese superoxide dismutase. PNAS 2012, 109, 14314–14319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enomoto, R.; Suzuki, C.; Ohno, M.; Ohasi, T.; Futagami, R.; Ishikawa, K.; Komae, M.; Nishino, T.; Konishi, Y.; Lee, E. Cationic surfactants induce apoptosis in normal and cancer cells. Ann. N. Y. Acad. Sci. 2007, 1095, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Park, C.J.; Song, S.H.; Kim, D.H.; Gye, M.C. Developmental and acute toxicity of cetylpyridinium chloride in Bombina orientalis (Amphibia: Anura). Aquatic Toxicol. 2016, 177, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Feng, Y.; Ye, Y.L.; Leng, Y.; Shen, J.H. Intestinally targeted TGR5 agonists equipped with quaternary ammonium have an improved hypoglycemic effect and reduced gallbladder filling effect. Sci. Rep. 2016, 6, 28676. [Google Scholar] [CrossRef]
- Hermenean, A.; Damache, G.; Albu, P.; Ardelean, A.; Ardelean, G.; Ardelean, P.D.; Horge, M.; Nagy, T.; Braun, M.; Zsuga, M.; et al. Histopatological alterations and oxidative stress in liver and kidney of Leuciscus cephalus following exposure to heavy metals in the Tur River, North Western Romania. Ecotoxicol. Environ. Saf. 2015, 119, 198–205. [Google Scholar] [CrossRef]
- Ma, J.; Zhu, J.; Wang, W.; Ruan, P.; Rajeshkumar, S.; Li, X. Biochemical and molecular impacts of glyphosate-based herbicide on the gills of common carp. Environ Pollut. 2019, 252, 1288–1300. [Google Scholar] [CrossRef]
- Gomes, I.D.; Nascimento, A.A.; Sales, A.; Araújo, F.G. Can fish gill anomalies be used to assess water quality in freshwater Neotropical systems? Environ. Monit. Assess. 2012, 184, 5523–5531. [Google Scholar] [CrossRef]
- Corredor-Santamaría, W.; Torres-Tabares, A.; Velasco-Santamaría, Y.M. Biochemical and histological alterations in Aequidens metae (Pisces, Cichlidae) and Astyanax gr. bimaculatus (Pisces, Characidae) as indicators of river pollution. Sci. Total Environ. 2019, 692, 1234–1241. [Google Scholar] [CrossRef] [PubMed]
- Roy, D. Detergent-induced changes in the mapping of certain enzymes in various cell types of Rita rita. II. Gill epithelium. Ecotoxicol. Environ. Saf. 1989, 17, 67–74. [Google Scholar] [CrossRef]
- Byrne, P.; Speare, D.; Ferguson, H.W. Effects of a cationic detergent on the gills and blood chemistry of rainbow trout Salmo gairdneri. Dis. Aquat. Org. 1989, 6, 185–196. [Google Scholar] [CrossRef]
- Patel, R.P.; Mandal, D.; Bahadur, A. Subchronic toxicity of ionic surfactants on freshwater fish Labeo rohita (Hamilton). Asian Fish. Sci. 2009, 22, 1137–1148. [Google Scholar] [CrossRef]
- Camargo, M.M.P.; Martinez, C.B.R. Histopathology of gills, kidney and liver of a Neotropical fish caged in an urban stream. Neotrop. Ichthyol. 2007, 5, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Li, M.H. Effects of nonionic and ionic surfactants on survival, oxidative stress, and cholinesterase activity of planarian. Chemosphere 2008, 70, 1796–1803. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gheorghe, S.; Stan, M.S.; Mitroi, D.N.; Staicu, A.C.; Cicirma, M.; Lucaciu, I.E.; Nita-Lazar, M.; Dinischiotu, A. Oxidative Stress and Histopathological Changes in Gills and Kidneys of Cyprinus carpio following Exposure to Benzethonium Chloride, a Cationic Surfactant. Toxics 2022, 10, 227. https://doi.org/10.3390/toxics10050227
Gheorghe S, Stan MS, Mitroi DN, Staicu AC, Cicirma M, Lucaciu IE, Nita-Lazar M, Dinischiotu A. Oxidative Stress and Histopathological Changes in Gills and Kidneys of Cyprinus carpio following Exposure to Benzethonium Chloride, a Cationic Surfactant. Toxics. 2022; 10(5):227. https://doi.org/10.3390/toxics10050227
Chicago/Turabian StyleGheorghe, Stefania, Miruna S. Stan, Daniel N. Mitroi, Andrea C. Staicu, Marius Cicirma, Irina E. Lucaciu, Mihai Nita-Lazar, and Anca Dinischiotu. 2022. "Oxidative Stress and Histopathological Changes in Gills and Kidneys of Cyprinus carpio following Exposure to Benzethonium Chloride, a Cationic Surfactant" Toxics 10, no. 5: 227. https://doi.org/10.3390/toxics10050227
APA StyleGheorghe, S., Stan, M. S., Mitroi, D. N., Staicu, A. C., Cicirma, M., Lucaciu, I. E., Nita-Lazar, M., & Dinischiotu, A. (2022). Oxidative Stress and Histopathological Changes in Gills and Kidneys of Cyprinus carpio following Exposure to Benzethonium Chloride, a Cationic Surfactant. Toxics, 10(5), 227. https://doi.org/10.3390/toxics10050227