
A Proposed Association between Improving Energy Metabolism of HepG2 Cells by Plant Extracts and Increasing Their Sensitivity to Doxorubicin

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Preparation of Plant Extracts
2.3. Cell Culture
2.4. Cytotoxicity Assay for Plant Extracts by MTT Assay
2.5. HepG2 Cell Treatment
2.6. Chemical Composition Analysis
2.6.1. UPLC/MS Analysis of Plant Extracts
2.6.2. GC/MS Analysis of Avocado Oil
2.7. Cytotoxicity Assay for Doxorubicin by MTT Using Untreated and Plant Extract-Pretreated HepG2 Cells
2.8. Colorimetric/ELISA Determination of Intracellular Proteins/Enzymes
2.9. Statistical Analysis
3. Results
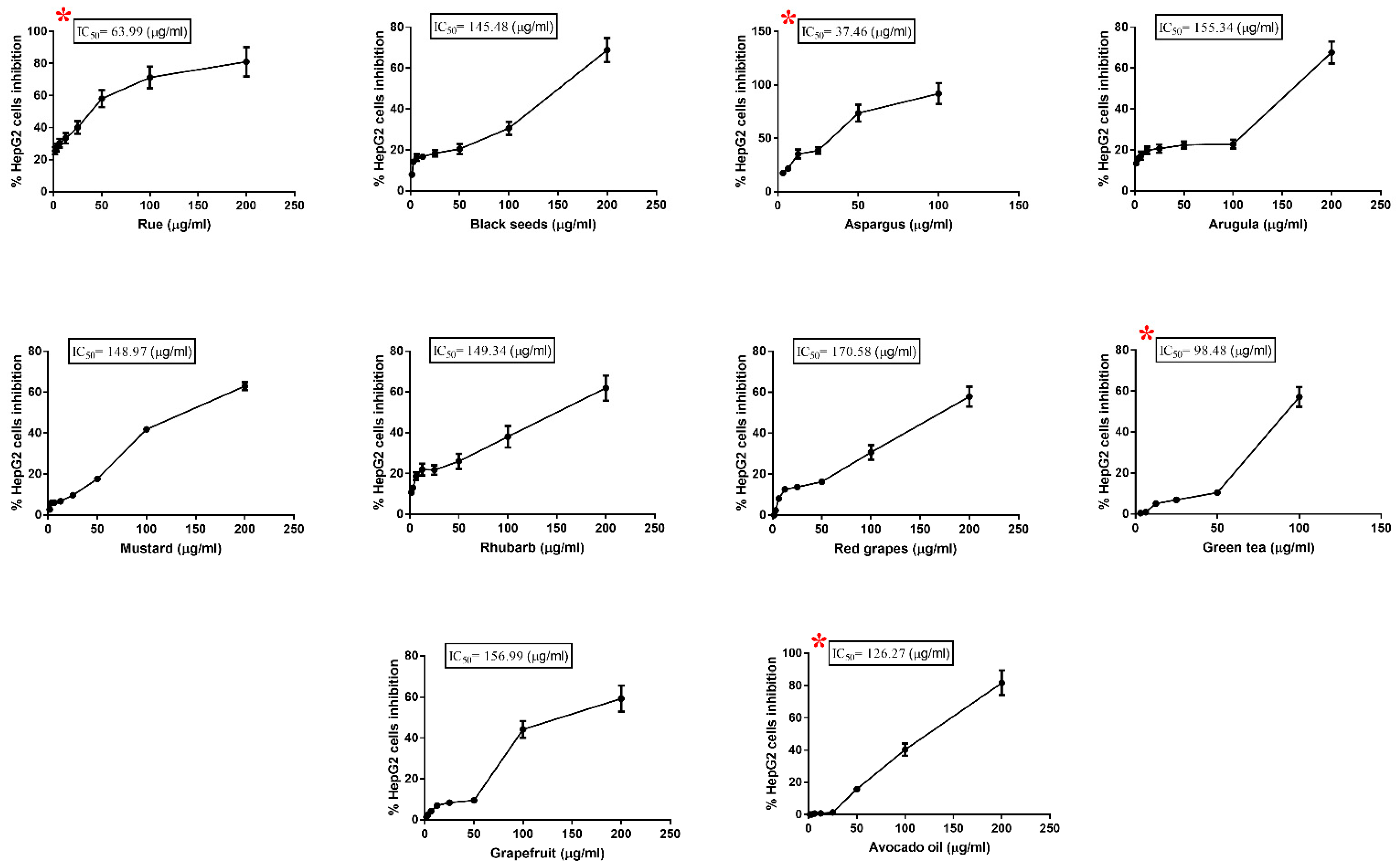
3.1. Cytotoxic Effect of Plant Extracts on HepG2 Cell
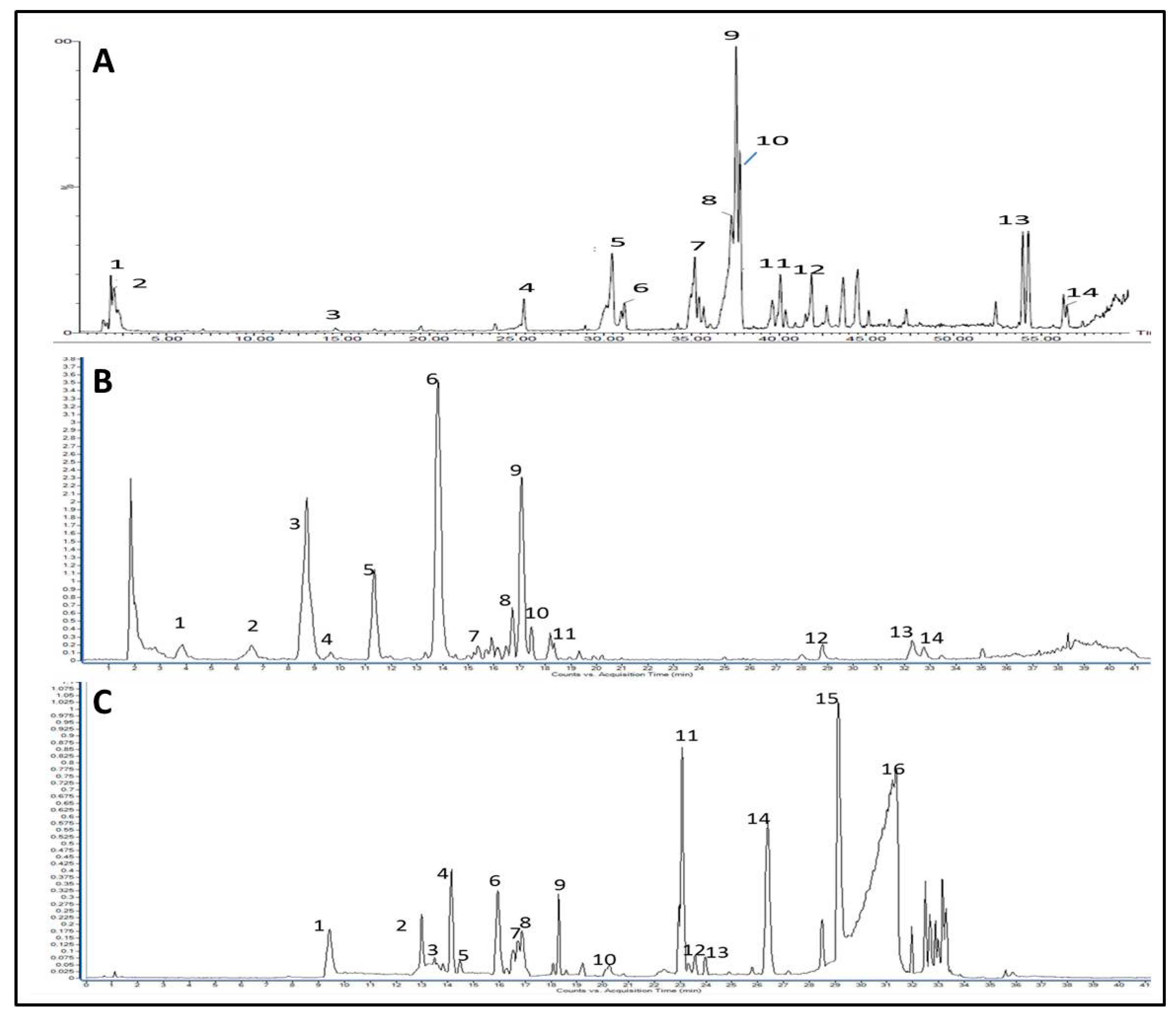
3.2. Identification of Major Phytoconstituents of the Investigated Extracts
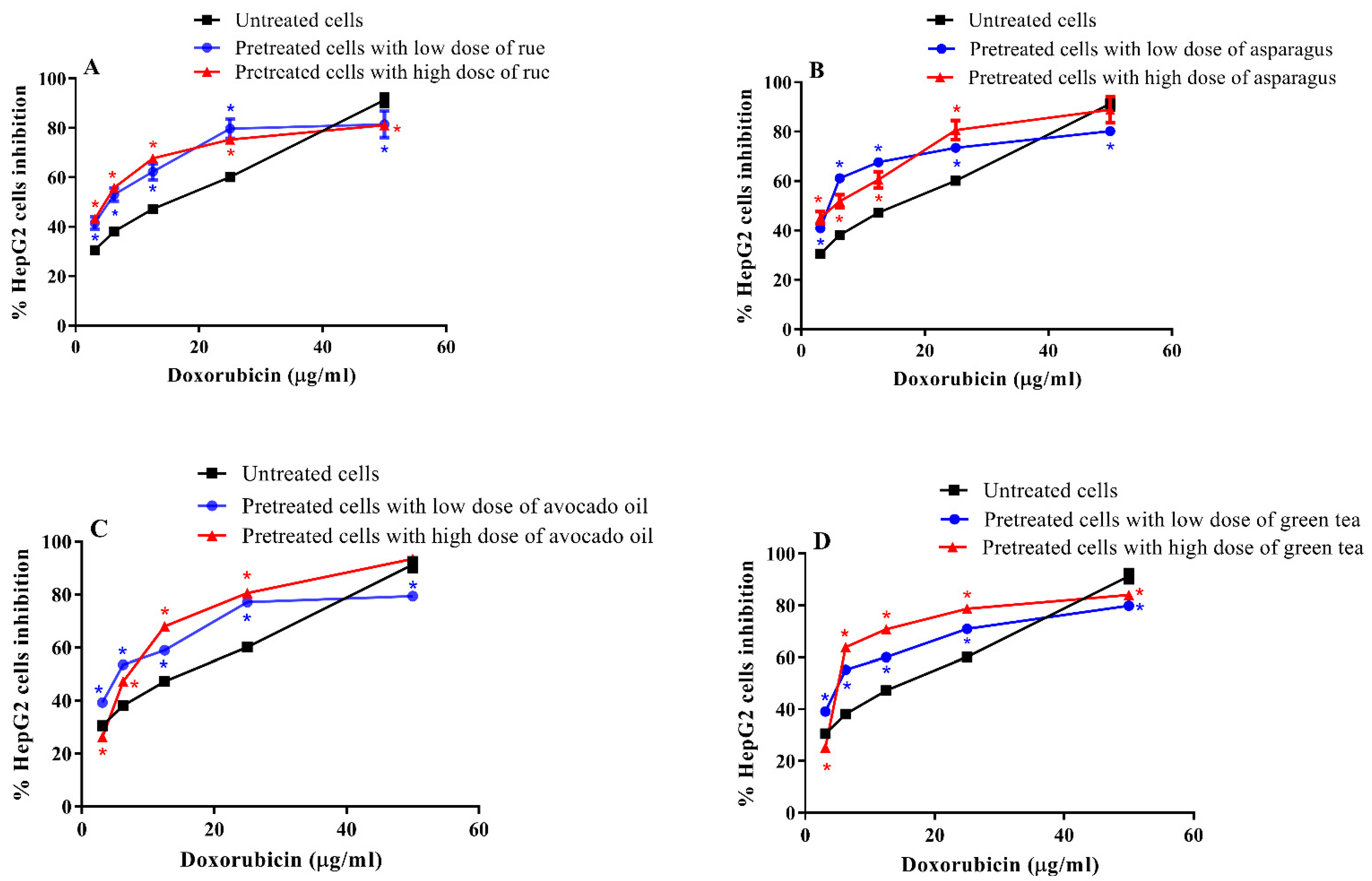
3.3. Cytotoxic Effect of Doxorubicin on Pretreated and Untreated HepG2 Cells
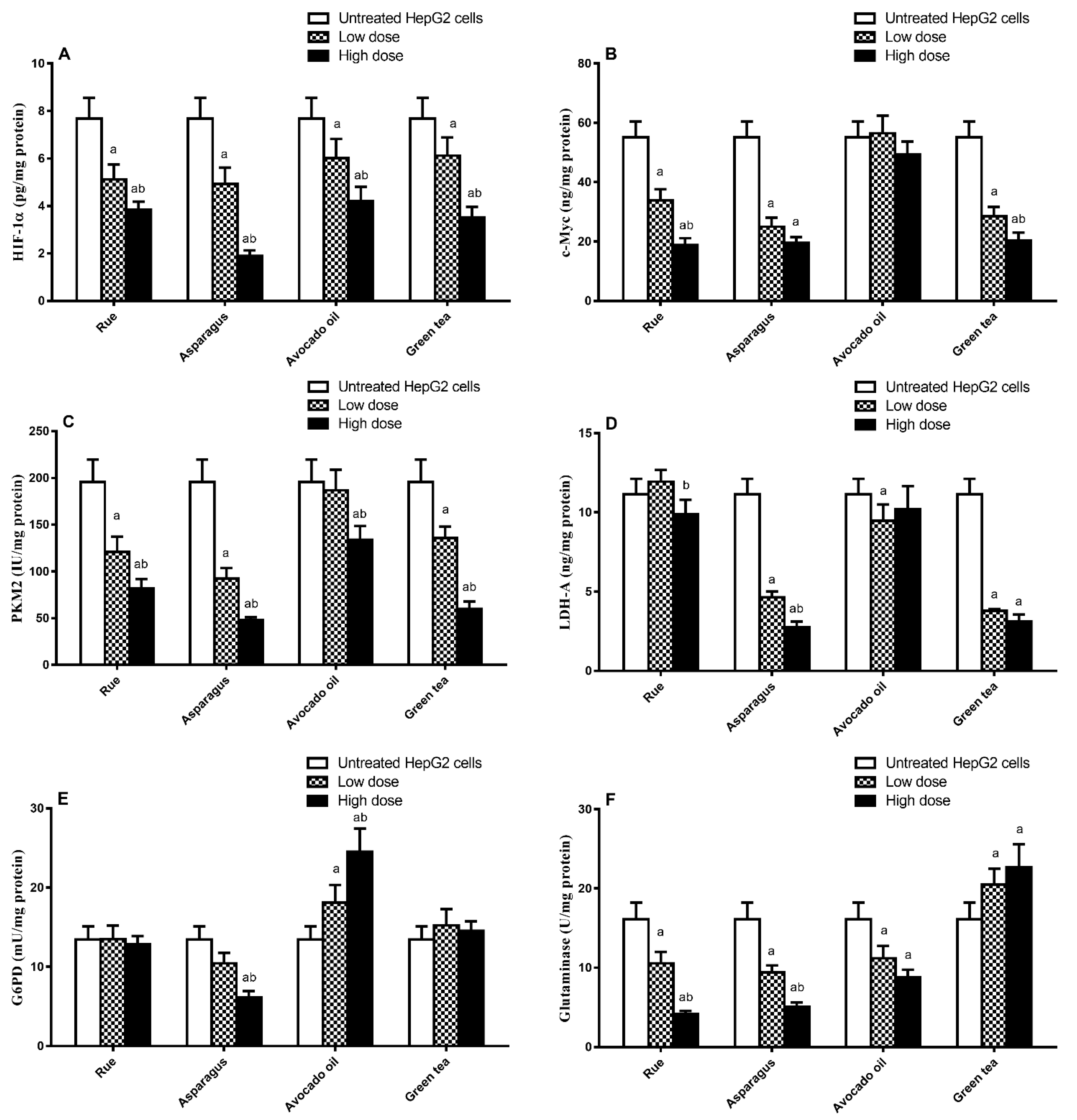
3.4. Effect of HepG2 Cell Treatment with Plant Extract on Metabolism-Regulating Enzymes/Proteins Level
3.5. Correlation between the Reduction (%) in Doxorubicin IC50 and the Change in Metabolism-Regulating Enzymes/Proteins Level
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, J.D.; Hainaut, P.; Gores, G.J.; Amadou, A.; Plymoth, A.; Roberts, L.R. A global view of hepatocellular carcinoma: Trends, risk, prevention and management. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 589–604. [Google Scholar] [CrossRef] [PubMed]
- Rashed, W.M.; Kandeil, M.A.M.; Mahmoud, M.O.; Ezzat, S. Hepatocellular carcinoma (HCC) in Egypt: A comprehensive overview. J. Egypt Natl. Cancer Inst. 2020, 32, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGowan, J.V.; Chung, R.; Maulik, A.; Piotrowska, I.; Walker, J.M.; Yellon, D.M. Anthracycline chemotherapy and cardiotoxicity. Cardiovasc. Drugs Ther. 2017, 31, 63–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Grazie, M.; Biagini, M.R.; Tarocchi, M.; Polvani, S.; Galli, A. Chemotherapy for hepatocellular carcinoma: The present and the future. World J. Hepatol. 2017, 9, 907–920. [Google Scholar] [CrossRef]
- Quinto, A.M.; Nutu, O.A.; Manso, R.S.R.; Alonso, I.J.; Pulido, J.C.; Municio, A.M.; García-Sesma, Á.; Segurola, C.L.; Caballero, J.M.; Romero, L.C.J. Complications of transarterial chemoembolization (TACE) in the treatment of liver tumors. Cirugía Española 2018, 96, 560–567. [Google Scholar] [CrossRef]
- Muntean, D.M.; Sturza, A.; Pavel, I.Z.; Duicu, O.M. Modulation of cancer metabolism by phytochemicals—A brief overview. Anticancer Agents Med. Chem. 2018, 18, 684–692. [Google Scholar] [CrossRef]
- Pistollato, F.; Giampieri, F.; Battino, M. The use of plant-derived bioactive compounds to target cancer stem cells and modulate tumor microenvironment. Food Chem. Toxicol. 2015, 75, 58070. [Google Scholar] [CrossRef]
- Zhang, Q.Y.; Wang, F.X.; Jia, K.K.; Kong, L.D. Natural product interventions for chemotherapy and radiotherapy-induced side effects. Front. Pharmacol. 2018, 9, 1253. [Google Scholar] [CrossRef] [Green Version]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxidative Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Cosme, P.; Rodríguez, A.B.; Espino, J.; Garrido, M. Plant Phenolics: Bioavailability as a Key Determinant of Their Potential Health-Promoting Applications. Antioxidants 2020, 9, 1263. [Google Scholar] [CrossRef]
- Kalyanaraman, B. Teaching the basics of cancer metabolism: Developing antitumor strategies by exploiting the differences between normal and cancer cell metabolism. Redox Biol. 2017, 12, 833–842. [Google Scholar] [CrossRef]
- Warburg, O.; Wind, F.; Negelein, E. The metabolism of tumors in the body. J. Gen. Physiol. 1927, 8, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Koppenol, W.H.; Bounds, P.L.; Dang, C.V. Otto Warburg’s contributions to current concepts of cancer metabolism. Nat. Rev. Cancer 2011, 11, 325–337. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Outschoorn, U.E.; Peiris-Pagés, M.; Pestell, R.G.; Sotgia, F.; Lisanti, M.P. Cancer metabolism: A therapeutic perspective. Nat. Rev. Clin. Oncol. 2017, 14, 11–31, Erratum in 14(12):113. [Google Scholar] [CrossRef]
- Jiménez-Sánchez, C.; Lozano-Sánchez, J.; Rodríguez-Pérez, C.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Comprehensive, untargeted, and qualitative RP-HPLC-ESI-QTOF/MS2 metabolite profiling of green asparagus (Asparagus officinalis). J. Food Compos. Anal. 2016, 46, 78–87. [Google Scholar] [CrossRef]
- Abdel-Wahab, A.H.A.; Effat, H.; Mahrous, E.A.; Ali, M.A.; Al-Shafie, T.A. A licorice roots extract induces apoptosis and cell cycle arrest and improves metabolism via regulating MiRNAs in liver cancer cells. Nutr. Cancer 2021, 73, 1047–1058. [Google Scholar] [CrossRef]
- Guerra, A.R.; Duarte, M.F.; Duarte, I.F. Targeting tumor metabolism with plant-derived natural products: Emerging trends in cancer therapy. J.Agric. Food Chem. 2018, 66, 10663–10685. [Google Scholar] [CrossRef]
- Lin, S.R.; Chang, C.H.; Hsu, C.F.; Tsai, M.J.; Cheng, H.; Leong, M.K.; Sung, P.J.; Chen, J.C.; Weng, C.F. Natural compounds as potential adjuvants to cancer therapy: Preclinical evidence. Br. J. Pharmacol. 2020, 177, 1409–1432. [Google Scholar] [CrossRef] [Green Version]
- Nalini, D.; Selvaraj, J.; Kumar, G.S. Herbal nutraceuticals: Safe and potent therapeutics to battle tumor hypoxia. J. Cancer Res. Clin. Oncol. 2020, 146, 1–18. [Google Scholar] [CrossRef]
- Golding, J.P.; Wardhaugh, T.; Patrick, L.; Turner, M.; Phillips, J.B.; Bruce, J.I.; Kimani, S.G. Targeting tumour energy metabolism potentiates the cytotoxicity of 5-aminolevulinic acid photodynamic therapy. Br. J. Cancer 2013, 109, 976–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Zong, X. Metabolic symbiosis in chemoresistance: Refocusing the role of aerobic glycolysis. Front. Oncol. 2020, 10, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capelôa, T.; Benyahia, Z.; Zampieri, L.X.; Blackman, M.C.N.M.; Sonveaux, P. Metabolic and non-metabolic pathways that control cancer resistance to anthracyclines. Semin. Cell Dev. Biol. 2020, 98, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, Y.S.; Lee, H.A.; Lim, Y.; Kim, Y. Mulberry leaf extract inhibits invasive potential and downregulates hypoxia-inducible factor-1α (HIF-1α) in SK-N-BE2C neuroblastoma cells. Biosci. Biotechnol. Biochem. 2013, 77, 722–728. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.J.; Cui, S.; Lu, D.X.; Yang, Y.Z.; Luo, Y.; Ma, L.; Ma, Y.; Wuren, T.; Chang, R.; Qi, L.; et al. Effects of the aqueous extract of a Tibetan herb, Rhodiola algida var. tangutica on proliferation and HIF-1α, HIF-2α expression in MCF-7 cells under hypoxic condition in vitro. Cancer Cell Int. 2015, 15, 81. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Ke, X.; Ma, N.; Wang, W.; Fu, W.; Zhang, H.; Zhao, M.; Gao, X.; Hao, X.; Zhang, Z. Formononetin, an active compound of Astragalus membranaceus (Fisch) Bunge, inhibits hypoxia-induced retinal neovascularization via the HIF-1α/VEGF signaling pathway. Drug Des. Dev. Ther. 2016, 10, 3071–3081. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Yao, S.; Cao, S.; Mo, G.; Li, J.; Cao, Y.; Huang, F. Triterpenoid Saponins from Anemone flaccida Suppress Tumor Cell Proliferation by Regulating MAPK, PD1/PDL1, and STAT3 Signaling Pathways and Altering Cancer Metabolism. OncoTargets Ther. 2019, 12, 10917–10930. [Google Scholar] [CrossRef] [Green Version]
- Jie, Y.; Yang, X.; Chen, W. Pulsatilla Decoction Combined with 5-Fluorouracil Triggers Immunogenic Cell Death in the Colorectal Cancer Cells. Cancer Biother. Radiopharm. 2021, 37, 945–954. [Google Scholar] [CrossRef]
- Zhou, P.; Wang, Y.; Li, B.; Liu, J.; Han, W.; He, X. Inhibitory Effect of Total Saponins of Pulsatilla on Inflammatory Microenvironment of Lung Cancer Mice and Its Mechanism. Rev. Científica-Fac. De Cienc. Vet. 2020, 30, 646–654. [Google Scholar]
- Slattery, E.; Cline, T.; Housley, L. Effects of Sulforaphane in the Tumor Microenvironment of Triple-Negative Breast Cancer. FASEB J. 2017, 31, 646–647. [Google Scholar] [CrossRef]
- Deep, G.; Kumar, R.; Jain, A.K.; Dhar, D.; Panigrahi, G.K.; Hussain, A.; Agarwal, C.; El-Elimat, T.; Sica, V.P.; Oberlies, N.H.; et al. Graviola inhibits hypoxia-induced NADPH oxidase activity in prostate cancer cells reducing their proliferation and clonogenicity. Sci. Rep. 2016, 6, 23135. [Google Scholar] [CrossRef] [Green Version]
- Awad, M.G.; Ali, R.A.; Abd El-Monem, D.D.; El-Magd, M.A. Graviola leaves extract enhances the anticancer effect of cisplatin on various cancer cell lines. Mol. Cell. Toxicol. 2020, 16, 385–399. [Google Scholar] [CrossRef]
- El-Sayed, A.; Fayed, R.H.; Castañeda Vázquez, H.; Rüegge, K. Future Trend to Replace Chemical Products with Nutraceutical Food/Feed Additive: A Mini Review. J. Appl. Vet. Sci. 2022, 7, 20–29. [Google Scholar] [CrossRef]
- Colucci-D’Amato, L.; Cimaglia, G. Ruta graveolens as a potential source of neuroactive compounds to promote and restore neural functions. J. Tradit. Complement. Med. 2020, 10, 309–314. [Google Scholar] [CrossRef]
- Ali, S.K.; Hamed, A.R.; Soltan, M.M.; Hegazy, U.M.; Elgorashi, E.E.; El-Garf, I.A.; Hussein, A.A. In-vitro evaluation of selected Egyptian traditional herbal medicines for treatment of Alzheimer disease. BMC Complement Altern. Med. 2013, 13, 121. [Google Scholar] [CrossRef] [Green Version]
- Hichri, F.; Omri Hichri, A.; Maha, M.; Saad Mana Hossan, A.; Flamini, G.; Ben Jannet, H. Chemical Composition, Antibacterial, Antioxidant and in Vitro Antidiabetic Activities of Essential Oils from Eruca vesicaria. Chem. Biodivers. 2019, 16, e1900183. [Google Scholar] [CrossRef]
- Guo, Q.; Wang, N.; Liu, H.; Li, Z.; Lu, L.; Wang, C. The bioactive compounds and biological functions of Asparagus officinalis L.—A review. J. Funct. Foods 2020, 65, 103727. [Google Scholar] [CrossRef]
- Wang, M.; Yu, P.; Chittiboyina, A.G.; Chen, D.; Zhao, J.; Avula, B.; Wang, Y.H.; Khan, I.A. Characterization, Quantification and Quality Assessment of Avocado (Persea americana Mill.) Oils. Molecules 2020, 25, 1453. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar]
- Kwee, J.K. A paradoxical chemoresistance and tumor suppressive role of antioxidant in solid cancer cells: A strange case of Dr. Jekyll and Mr. Hyde. Biomed. Res. Int. 2014, 2014, 209845. [Google Scholar] [CrossRef] [Green Version]
- Wierenga, A.T.J.; Cunningham, A.; Erdem, A.; Lopera, N.V.; Brouwers-Vos, A.Z.; Pruis, M.; Mulder, A.B.; Günther, U.L.; Martens, J.H.A.; Vellenga, E.; et al. HIF1/2-exerted control over glycolytic gene expression is not functionally relevant for glycolysis in human leukemic stem/progenitor cells. Cancer Metab. 2019, 7, 11. [Google Scholar] [CrossRef]
- Milane, L.; Duan, Z.; Amiji, M. Role of hypoxia and glycolysis in the development of multi-drug resistance in human tumor cells and the establishment of an orthotopic multi-drug resistant tumor model in nude mice using hypoxic pre-conditioning. Cancer Cell Int. 2011, 11, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masson, N.; Ratcliffe, P.J. Hypoxia signaling pathways in cancer metabolism: The importance of co-selecting interconnected physiological pathways. Cancer Metab. 2014, 2, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doe, M.R.; Ascano, J.M.; Kaur, M.; Cole, M.D. Myc posttranscriptionally induces HIF1 protein and target gene expression in normal and cancer cells. Cancer Res. 2012, 72, 949–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Venneti, S.; Nagrath, D. Glutaminolysis: A hallmark of cancer metabolism. Annu. Rev. Biomed. Eng. 2017, 19, 163–194. [Google Scholar] [CrossRef] [PubMed]
- Acheampong, D.O.; Baffour, I.K.; Atsu Barku, V.Y.; Addo, J.K.; Essuman, M.A.; Boye, A. Zanthoxylum zanthoxyloides Alkaloidal Extract Improves CCl(4)-Induced Hepatocellular Carcinoma-Like Phenotypes in Rats. Evid.-Based Complement. Altern. Med. Ecam 2021, 2021, 3804379. [Google Scholar] [CrossRef]
- Guo, W.; Tan, H.Y.; Chen, F.; Wang, N.; Feng, Y. Targeting cancer metabolism to resensitize chemotherapy: Potential development of cancer chemosensitizers from traditional chinese medicines. Cancers 2020, 12, 404. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.C.; Chand, R.; Bhatti, B.S.; Sati, O.P. New oligospirostanosides and oligofurostanosides from Asparagus adscendens roots. Planta Med. 1982, 46, 48–51. [Google Scholar] [CrossRef]
- Zhang, H.J.; Sydara, K.; Tan, G.T.; Ma, C.; Southavong, B.; Soejarto, D.D.; Pezzuto, J.M.; Fong, H.H. Bioactive constituents from Asparagus cochinchinensis. J. Nat. Prod. 2004, 67, 194–200. [Google Scholar] [CrossRef]
- Lu, J.; Chen, H.; He, F.; You, Y.; Feng, Z.; Chen, W.; Li, X.; Zhao, L. Ginsenoside 20(S)-Rg3 upregulates HIF-1α-targeting miR-519a-5p to inhibit the Warburg effect in ovarian cancer cells. Clin. Exp. Pharmacol. Physiol. 2020, 47, 1455–1463. [Google Scholar] [CrossRef]
- Li, J.; Liu, T.; Zhao, L.; Chen, W.; Hou, H.; Ye, Z.; Li, X. Ginsenoside 20(S)-Rg3 inhibits the Warburg effect through STAT3 pathways in ovarian cancer cells. Int. J. Oncol. 2015, 46, 775–781. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Tu, R.; Liu, H.; Qing, G. Regulation of cancer cell metabolism: Oncogenic MYC in the driver’s seat. Signal Transduct. Target Ther. 2020, 5, 124. [Google Scholar] [CrossRef]
- Cluntun, A.A.; Lukey, M.J.; Cerione, R.A.; Locasale, J.W. Glutamine metabolism in vcancer: Understanding the heterogeneity. Trends Cancer 2017, 3, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Malik, S.; Moraes, D.F.C.; do Amaral, F.M.M.; Ribeiro, M.N.S. Ruta graveolens: Phytochemistry, pharmacology, and biotechnology. In Transgenesis and Secondary Metabolism.; Jha, S., Ed.; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Kawaii, S.; Tomono, Y.; Katase, E.; Ogawa, K.; Yano, M.; Takemura, Y.; Ju-ichi, M.; Ito, C.; Furukawa, H. The antiproliferative effect of acridone alkaloids on several cancer cell lines. J. Nat. Prod. 1999, 62, 587–589. [Google Scholar] [CrossRef]
- Réthy, B.; Zupkó, I.; Minorics, R.; Hohmann, J.; Ocsovszki, I.; Falkay, G. Investigation of cytotoxic activity on human cancer cell lines of arborinine and furanoacridones isolated from Ruta graveolens. Planta Med. 2007, 73, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Richardson, J.S.M.; Sethi, G.; Lee, G.S.; Abdul Malek, S.N. Chalepin: Isolated from Ruta angustifolia L. Pers induces mitochondrial mediated apoptosis in lung carcinoma cells. BMC Complement Altern. Med. 2016, 16, 389. [Google Scholar] [CrossRef] [Green Version]
- Rietveld, A.; Wiseman, S. Antioxidant effects of tea: Evidence from human clinical trials. J Nutr 2003, 133, 3285S–3292S. [Google Scholar] [CrossRef] [Green Version]
- Bhuyan, D.J.; Alsherbiny, M.A.; Perera, S.; Low, M.; Basu, A.; Devi, O.A.; Barooah, M.S.; Li, C.G.; Papoutsis, K. The Odyssey of Bioactive Compounds in Avocado (Persea americana) and Their Health Benefits. Antioxidants 2019, 8, 426. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira Júnior, R.G.; Christiane Adrielly, A.F.; da Silva Almeida, J.R.G.; Grougnet, R.; Thiéry, V.; Picot, L. Sensitization of tumor cells to chemotherapy by natural products: A systematic review of preclinical data and molecular mechanisms. Fitoterapia 2018, 129, 383–400. [Google Scholar] [CrossRef]
- Almatroodi, S.A.; Almatroudi, A.; Khan, A.A.; Alhumaydhi, F.A.; Alsahli, M.A.; Rahmani, A.H. Potential therapeutic targets of Epigallocatechin Gallate (EGCG), the most abundant catechin in green tea, and its role in the therapy of various types of cancer. Molecules 2020, 25, 3146. [Google Scholar] [CrossRef]
- Lara-Marquez, M.; Baez-Magana, M.; Raymundo-Ramos, C.; Spagnuolo, P.A.; Macias-Rodriguez, L.; Salgado-Garciglia, R.; Ochoa-Zarzosa, A.; Lopez-Meza, J.E. Lipid-rich extract from Mexican avocado (Persea americana var. drymifolia) induces apoptosis and modulates the inflammatory response in Caco-2 human colon cancer cells. J. Funct. Foods 2020, 64, 103658. [Google Scholar] [CrossRef]
- Savić, I.M.; Nikolić, V.D.; Savić, I.M.; Nikolić, L.B.; Jović, M.D.; Jović, M. The qualitative analysis of the green tea extract using ESI-MS method. Savrem. Tehnol. 2014, 3, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Kim, M.K. Effects of Different Harvesting Times and Oxidative Fermentation Methods on Phytochemicals, Flavors, and Sensory Properties of Korean Teas. In Chemistry of Korean Foods and Beverages; ACS Symposium Series: Washington, DC, USA, 2019; pp. 77–95. [Google Scholar]
- Stöggl, W.M.; Huck, C.W.; Bonn, G.K. Structural elucidation of catechin and epicatechin in sorrel leaf extracts using liquid-chromatography coupled to diode array-, fluorescence-, and mass spectrometric detection. J. Sep. Sci. 2004, 27, 524–528. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Bishayee, K.; Khuda-Bukhsh, A.R. Graveoline isolated from ethanolic extract of Ruta graveolens triggers apoptosis and autophagy in skin melanoma cells: A novel apoptosis-independent autophagic signaling pathway. Phytother. Res. 2014, 8, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Zobel, A.M.; Brown, S.A. Determination of furanocoumarins on the leaf surface of Ruta graveolens with an improved extraction technique. J. Nat. Prod. 1988, 51, 941–946. [Google Scholar] [CrossRef]
- Mancuso, G.; Borgonovo, G.; Scaglioni, L.; Bassoli, A. Phytochemicals from Ruta graveolens activate TAS2R bitter taste receptors and TRP channels involved in gustation and nociception. Molecules 2015, 20, 18907–18922. [Google Scholar] [CrossRef] [Green Version]
- Kostova, I.; Ivanova, A.; Mikhova, B.; Klaiber, I. Alkaloids and coumarins from Ruta graveolens. Mon. Fur Chem. 1999, 130, 703–707. [Google Scholar] [CrossRef]
- França Orlanda, J.F.; Nascimento, A.R. Chemical composition and antibacterial activity of Ruta graveolens L. (Rutaceae) volatile oils, from São Luís, Maranhão, Brazil. S. Afr. J. Bot 2015, 99, 103–106. [Google Scholar] [CrossRef]
- SSampaio, O.M.; Vieira, L.C.C.; Bellete, B.S.; King-Diaz, B.; Lotina-Hennsen, B.; Da Silva, M.F.d.G.F.; Veiga, T.A.M. Evaluation of alkaloids isolated from Ruta graveolens as photosynthesis inhibitors. Molecules 2018, 23, 2693. [Google Scholar] [CrossRef] [Green Version]
- Oh, E.T.; Lee, J.H.; Kim, C.S.; Keum, Y.S. Alkylquinolone alkaloid profiles in Ruta graveolens. Biochem. Syst. Ecol. 2014, 57, 384–387. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Extract | IC50 (µg/mL) |
|---|---|
| Asparagus Officinalis | 37.46 |
| Ruta graveolens | 63.99 |
| Green tea | 98.48 |
| Avocado fruit | 126.27 |
| Nigella Sativa | 145.48 |
| Mustard | 148.97 |
| Rhubarb | 149.34 |
| Arugula | 155.34 |
| Grapefruit | 156.99 |
| Red grapes | 170.58 |
| Plant Extract Treating Dose (µg/mL) | Doxorubicin IC50 (µg/mL) | p-Value | |
|---|---|---|---|
| CTRL (Untreated cells) | - | 7.99 ± 0.41 | |
| Rue-pretreated cells | Lo: 8.62 | 5.57 ± 0.35 | 0.012 * |
| Hi: 12.08 | 4.32 ± 0.24 | 0.002 ** | |
| Asparagus-pretreated cells | Lo: 11.62 | 3.38 ± 0.19 | 0.001 ** |
| Hi: 18.08 | 1.73 ± 0.12 | 0.001 ** | |
| Avocado-pretreated cells | Lo: 44.56 | 6.82 ± 0.42 | 0.118 |
| Hi: 71.52 | 5.34 ± 0.31 | 0.007 ** | |
| Green tea-pretreated cells | Lo: 55.57 | 6.05 ± 0.32 | 0.021 * |
| Hi: 90.92 | 4.80 ± 0.26 | 0.003 ** |
| HepG2 Treatment-Induced Reduction in Doxorubicin IC50 (%) | HIF1α (ng/mL) | C-Myc (μg/mL) | PKM2 (ng/mL) | LDH-A (μmol/mL) | G6PD (mU/mL) | Glutaminase (μg/mL) | |
|---|---|---|---|---|---|---|---|
| Rue-treated cells | r | 0.916 | 0.883 | 0.934 | 0.269 | 0.192 | 0.884 |
| p-value | 0.010 * | 0.020 * | 0.006 ** | 0.606 | 0.716 | 0.019 * | |
| Asparagus-treated cells | r | 0.957 | 0.836 | 0.909 | 0.903 | 0.977 | 0.963 |
| p-value | 0.003 ** | 0.038 * | 0.012 * | 0.014 * | 0.001 ** | 0.002 ** | |
| Avocado-treated cells | r | 0.939 | −0.891 | 0.117 | 0.097 | −0.859 | −0.374 |
| p-value | 0.005 ** | 0.017 * | 0.825 | 0.855 | 0.028 * | 0.465 | |
| Green tea-treated cells | r | 0.854 | 0.950 | 0.928 | 0.857 | −0.888 | −0.850 |
| p-value | 0.031 * | 0.004 ** | 0.008 ** | 0.029 * | 0.018 * | 0.32 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Shafie, T.A.; Mahrous, E.A.; Shukry, M.; Alshahrani, M.Y.; Ibrahim, S.F.; Fericean, L.; Abdelkader, A.; Ali, M.A. A Proposed Association between Improving Energy Metabolism of HepG2 Cells by Plant Extracts and Increasing Their Sensitivity to Doxorubicin. Toxics 2023, 11, 182. https://doi.org/10.3390/toxics11020182
Al-Shafie TA, Mahrous EA, Shukry M, Alshahrani MY, Ibrahim SF, Fericean L, Abdelkader A, Ali MA. A Proposed Association between Improving Energy Metabolism of HepG2 Cells by Plant Extracts and Increasing Their Sensitivity to Doxorubicin. Toxics. 2023; 11(2):182. https://doi.org/10.3390/toxics11020182
Chicago/Turabian StyleAl-Shafie, Tamer A., Engy A. Mahrous, Mustafa Shukry, Mohammad Y. Alshahrani, Samah F. Ibrahim, Liana Fericean, Afaf Abdelkader, and Mennatallah A. Ali. 2023. "A Proposed Association between Improving Energy Metabolism of HepG2 Cells by Plant Extracts and Increasing Their Sensitivity to Doxorubicin" Toxics 11, no. 2: 182. https://doi.org/10.3390/toxics11020182
APA StyleAl-Shafie, T. A., Mahrous, E. A., Shukry, M., Alshahrani, M. Y., Ibrahim, S. F., Fericean, L., Abdelkader, A., & Ali, M. A. (2023). A Proposed Association between Improving Energy Metabolism of HepG2 Cells by Plant Extracts and Increasing Their Sensitivity to Doxorubicin. Toxics, 11(2), 182. https://doi.org/10.3390/toxics11020182