
Relation among Mercury, Selenium, and Biomarkers of Oxidative Stress in Northern Pike (Esox lucius)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Measures of Gene Expression
2.3. Mercury Determination
2.4. Selenium Determination
2.5. Statistical Analysis
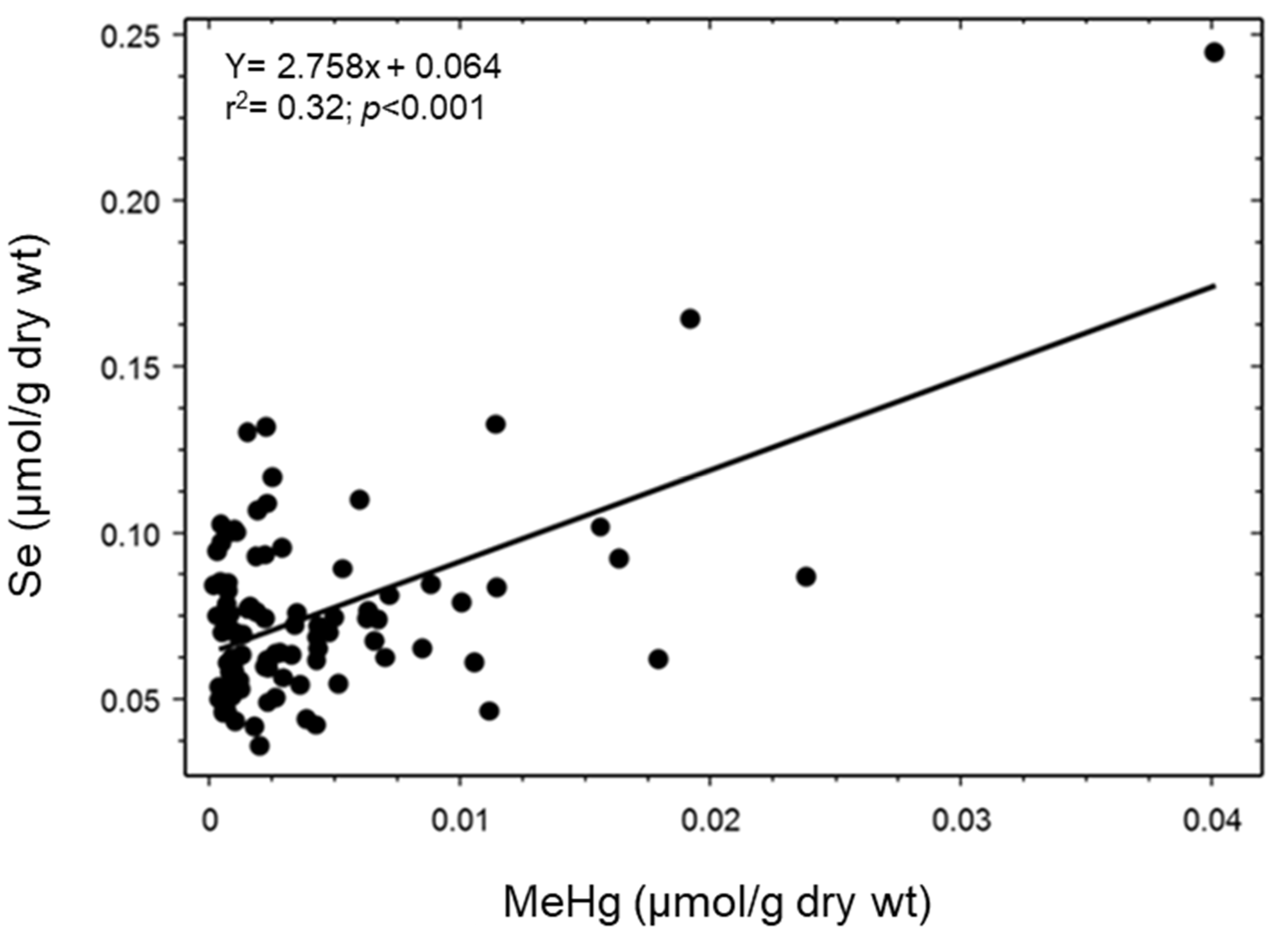
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kidd, K.; Batchelar, K. Mercury. In Homeostasis and Toxicology of Non-Essential Metals; Wood, C.M., Farrell, A.P., Brauner, C.J., Eds.; Elsevier: New York, NY, USA; Academic Press: New York, NY, USA, 2012; Volume 31, pp. 237–295. [Google Scholar] [CrossRef]
- Yang, D.-Y.; Chen, Y.-W.; Gunn, J.M.; Belzile, N. Selenium and mercury in organisms: Interactions and mechanisms. Environ. Rev. 2008, 16, 71–92. [Google Scholar] [CrossRef]
- Biester, H.; Müller, G.; Schöler, H.F. Binding and mobility of mercury in soils contaminated by emissions from chlor-alkali plants. Sci. Total Environ. 2002, 284, 191–203. [Google Scholar] [CrossRef]
- Hammerschmidt, C.R.; Fitzgerald, W.F. Methylmercury in freshwater fish linked to atmospheric mercury deposition. Environ. Sci. Technol. 2006, 40, 7764–7770. [Google Scholar] [CrossRef]
- Pacyna, E.G.; Pacyna, J.M.; Sundseth, K.; Munthe, J.; Kindbom, K.; Wilson, S.; Steenhuisen, F.; Maxson, P. Global emission of mercury to the atmosphere from anthropogenic sources in 2005 and projections to 2020. Atmos. Environ. 2013, 44, 2487–2499. [Google Scholar] [CrossRef]
- Pacyna, E.G.; Pacyna, J.M.; Steenhuisen, F.; Wilson, S. Global anthropogenic mercury emission inventory for 2000. Atmos. Environ. 2006, 40, 4048–4063. [Google Scholar] [CrossRef]
- Wiener, J.G.; Krabbenhoft, D.P.; Heinz, G.H.; Scheuhammer, A.M. Ecotoxicology of mercury. In Handbook of Ecotoxicology, 2nd ed.; Hofman, D.J., Rattner, B.A., Burton, G.A., Jr., Cairns, J.J., Eds.; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Trudel, M.; Rasmussen, J.B. Modeling the elimination of mercury by fish. Environ. Sci. Technol. 1997, 31, 1716–1722. [Google Scholar] [CrossRef]
- Oliveira Ribeiro, C.A.; Rouleau, C.; Pelletier, É.; Audet, C.; Tjälve, H. Distribution kinetics of dietary methylmercury in the Arctic charr (Salvelinus alpinus). Environ. Sci. Technol. 1999, 33, 902–907. [Google Scholar] [CrossRef]
- Southworth, G.R.; Turner, R.R.; Peterson, M.J.; Bogle, M.A. Form of mercury in stream fish exposed to high concentrations of dissolved inorganic mercury. Chemosphere 1995, 30, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Bloom, N.S. On the chemical form of mercury in edible fish and marine invertebrate tissue. Can. J. Fish. Aquat. Sci. 1992, 49, 1010–1017. [Google Scholar] [CrossRef]
- Scheuhammer, A. The chronic toxicity of aluminium, cadmium, mercury, and lead in birds: A review. Environ. Pollut. 1987, 46, 263–295. [Google Scholar] [CrossRef] [PubMed]
- Scheuhammer, A.M.; Meyer, M.W.; Sandheinrich, M.B.; Murray, M.W. Effects of environmental methylmercury on the health of wild birds, mammals, and fish. AMBIO 2007, 36, 12–18. [Google Scholar] [CrossRef]
- Wren, C.D. A review of metal accumulation and toxicity in wild mammals. Environ. Res. 1986, 40, 210–244. [Google Scholar] [CrossRef]
- Wolfe, M.F.; Schwarzbach, S.; Sulaiman, R.A. Effects of mercury on wildlife: A comprehensive review. Environ. Toxicol. Chem. 1998, 17, 146–160. [Google Scholar] [CrossRef]
- Burger, J. Trace element levels in pine snake hatchlings: Tissue and temporal differences. Arch. Environ. Contam. Toxicol. 1992, 22, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, B.C.; Willson, J.D.; Hopkins, W.A. Mercury exposure is associated with negative effects on turtle reproduction. Environ. Sci. Technol. 2013, 47, 2416–2422. [Google Scholar] [CrossRef] [PubMed]
- Depew, D.C.; Basu, N.; Burgess, N.M.; Campbell, L.M.; Devlin, E.W.; Drevnick, P.E.; Hammerschmidt, C.R.; Murphy, C.A.; Sandheinrich, M.B.; Wiener, J.G. Toxicity of dietary methylmercury to fish: Derivation of ecologically meaningful threshold concentrations. Environ. Toxicol. Chem. 2012, 31, 1536–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandheinrich, M.B.; Wiener, J.G. Methylmercury in freshwater fish: Recent advances in assessing toxicity of environmentaly relevant exposures. In Environmental Contaminants in Biota, 2nd ed.; Beyer, W.N., Meador, J.P., Eds.; CRC Press: Boca Raton, FL, USA, 2011; ISBN 9781420084054. [Google Scholar]
- Berntssen, M.H.G.; Aatland, A.; Handy, R.D. Chronic dietary mercury exposure causes oxidative stress, brain lesions, and altered behaviour in Atlantic salmon (Salmo salar) parr. Aquat. Toxicol. 2003, 65, 55–72. [Google Scholar] [CrossRef]
- Larose, C.; Canuel, R.; Lucotte, M.; Di Giulio, R.T. Toxicological effects of methylmercury on walleye (Sander vitreus) and perch (Perca flavescens) from lakes of the boreal forest. Comp. Biochem. Physiol. C. Toxicol. Pharmacol. 2008, 147, 139–149. [Google Scholar] [CrossRef]
- Drevnick, P.E.; Roberts, A.P.; Otter, R.R.; Hammerschmidt, C.R.; Klaper, R.; Oris, J.T. Mercury toxicity in livers of northern pike (Esox lucius) from Isle Royale, USA. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2008, 147, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Ralston, N.V.C.; Raymond, L.J. Dietary selenium’s protective effects against methylmercury toxicity. Toxicology 2010, 278, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Parízek, J.; Ostádalová, I. The protective effect of small amounts of selenite in sublimate intoxication. Experientia 1967, 23, 142–143. [Google Scholar] [CrossRef]
- Bjerregaard, P.; Fjordside, S.; Hansen, M.G.; Petrova, M.B. Dietary selenium reduces retention of methyl mercury in freshwater fish. Environ. Sci. Technol. 2011, 45, 9793–9798. [Google Scholar] [CrossRef]
- Ganther, A.H.E.; Goudie, C.; Sunde, M.L.; Kopecky, M.J.; Wagner, P.; Oh, S.-H.; Hoekstra, W.G. Selenium: Relation to decreased toxicity of methylmercury added to diets containing tuna. Science 1972, 175, 1122–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindh, U.; Johansson, E. Mercury effects of selenium against mercury toxicity as studied in the rat liver and kidney by nuclear analytical techniques. Biol. Trace Elem. Res. 1987, 12, 109–120. [Google Scholar] [CrossRef]
- Ralston, N.V.C.; Blackwell, J.L.; Raymond, L.J. Importance of molar ratios in selenium-dependent protection against methylmercury toxicity. Biol. Trace Elem. Res. 2007, 119, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Southworth, G.R.; Peterson, M.J.; Ryon, M.G. Long-term increased bioaccumulation of mercury in largemouth bass follows reduction of waterborne selenium. Chemosphere 2000, 41, 1101–1105. [Google Scholar] [CrossRef] [PubMed]
- Behne, D.; Weiss-Nowak, C.; Kalcklösch, M.; Westphal, C.; Gessner, H.; Kyriakopoulos, A. Studies on the distribution and characteristics of new mammalian selenium-containing proteins. Analyst 1995, 120, 823–825. [Google Scholar] [CrossRef]
- Schwarz, K.; Foltz, C.M. Selenium as an integral part of factor 3 against dietary necrotic liver degeneration. J. Biol. Chem. 1957, 233, 245–251. [Google Scholar] [CrossRef]
- Dang, F.; Wang, W.-X. Antagonistic interaction of mercury and selenium in a marine fish is dependent on their chemical species. Environ. Sci. Technol. 2011, 45, 3116–3122. [Google Scholar] [CrossRef]
- Janz, D.M. Selenium. In Homeostasis and Toxicology of Essential Metals; Fish Physiology; Wood, C.M., Farrell, A.P., Brauner, C.J., Eds.; Elsevier: New York, NY, USA; Academic Press: New York, NY, USA, 2012; Volume 31, pp. 327–374. [Google Scholar]
- Sorensen, M.; Bjerregaard, P. Interactive accumulation of mercury and selenium in the sea star Asterias rubens. Mar. Biol. 1991, 108, 269–276. [Google Scholar] [CrossRef]
- Eisler, R. Handbook of Chemical Risk Assessment: Health Hazards to Humans, Plants, and Animals; Lewis Publishers: Boca Raton, FL, USA, 2000; Volume 3. [Google Scholar]
- Gerson, J.R.; Walters, D.M.; Eagles-Smith, C.A.; Bernhardt, E.S.; Brandt, J.E. Do Two Wrongs Make a Right? Persistent Uncertainties Regarding Environmental Selenium-Mercury Interactions. Environ. Sci. Technol. 2020, 54, 9228–9234. [Google Scholar] [CrossRef]
- Wiener, J.G.; Haro, R.J.; Rolfhus, K.R.; Sandheinrich, M.B.; Bailey, S.W.; Northwick, R.M.; Gostomski, T.J. Bioaccumulative Contaminants in Aquatic Food Webs in Six National Park Units of the Western Great Lakes Region: 2008–2012; CreateSpace Independent: Fort Collins, CO, USA, 2016. [Google Scholar]
- Hammerschmidt, C.R.; Fitzgerald, W.F. Methylmercury in mosquitoes related to atmospheric mercury deposition and contamination. Environ. Sci. Technol. 2005, 39, 3034–3039. [Google Scholar] [CrossRef] [PubMed]
- Horvat, M.; Liang, L.; Bloom, N.S. Comparison of distillation with other current isolation methods for the determination of methyl mercury compounds in low level environmental samples. Anal. Chim. Acta 1993, 282, 153–168. [Google Scholar] [CrossRef]
- Olson, M.L.; Cleckner, L.B.; Hurley, J.P.; Heelan, T.W.; Krabbenhoft, D.P. Resolution of matrix effects on analysis of total and methyl mercury in aqueous samples from the Florida Everglades. Fresenius. J. Anal. Chem. 1997, 358, 392–396. [Google Scholar] [CrossRef]
- Cade, B.S.; Noon, B.R. A gentle introduction to quantile regression for ecologists. Front. Ecol. Env. 2003, 1, 412–420. [Google Scholar] [CrossRef]
- Gonzalez, P.; Dominique, Y.; Massabuau, J.C.; Boudou, A.; Bourdineaud, J.P. Comparative effects of dietary methylmercury on gene expression in liver, skeletal muscle, and brain of the zebrafish (Danio rerio). Environ. Sci. Technol. 2005, 39, 3972–3980. [Google Scholar] [CrossRef] [PubMed]
- Klaper, R.; Carter, B.J.; Richter, C.A.; Drevnick, P.E.; Sandheinrich, M.B.; Tillitt, D.E. Use of a 15 k gene microarray to determine gene expression changes in response to acute and chronic methylmercury exposure in the fathead minnow Pimephales promelas Rafinesque. J. Fish Biol. 2008, 72, 2207–2280. [Google Scholar] [CrossRef]
- Klaper, R.; Rees, C.B.; Drevnick, P.; Weber, D.; Sandheinrich, M.; Carvan, M.J. Gene expression changes related to endocrine function and decline in reproduction in fathead minnow (Pimephales promelas) after dietary methylmercury exposure. Environ. Health Perspect. 2006, 114, 1337–1343. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, D.A.; Rantin, F.T.; Kalinin, A.L. Inorganic mercury exposure: Toxicological effects, oxidative stress biomarkers and bioaccumulation in the tropical freshwater fish matrinxã, Brycon amazonicus (Spix and Agassiz, 1829). Ecotoxicology 2010, 19, 105–123. [Google Scholar] [CrossRef]
- Monteiro, D.A.; Rantin, F.T.; Kalinin, A.L. Dietary intake of inorganic mercury: Bioaccumulation and oxidative stress parameters in the neotropical fish Hoplias malabaricus. Ecotoxicology 2013, 22, 446–456. [Google Scholar] [CrossRef]
- Huang, W.; Cao, L.; Ye, Z.; Yin, X.; Dou, S. Antioxidative responses and bioaccumulation in Japanese flounder larvae and juveniles under chronic mercury exposure. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2010, 152, 99–106. [Google Scholar] [CrossRef]
- Hinck, J.E.; Blazer, V.S.; Denslow, N.D.; Myers, M.S.; Gross, T.S.; Tillitt, D.E. Biomarkers of contaminant exposure in northern pike (Esox lucius) from the Yukon River Basin, Alaska. Arch. Environ. Contam. Toxicol. 2007, 52, 549–562. [Google Scholar] [CrossRef]
- Graves, S.D.; Kidd, K.A.; Batchelar, K.L.; Cowie, A.M.; O’Driscoll, N.J.; Martyniuk, C.J. Response of oxidative stress transcripts in the brain of wild yellow perch (Perca flavescens) exposed to an environmental gradient of methylmercury. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 192, 50–58. [Google Scholar] [CrossRef]
- Sørmo, E.G.; Ciesielski, T.M.; Øverjordet, I.B.; Lierhagen, S.; Eggen, G.S.; Berg, T.; Jenssen, B.M. Selenium moderates mercury toxicity in free-ranging freshwater fish. Environ. Sci. Technol. 2011, 45, 6561–6566. [Google Scholar] [CrossRef]
- Chen, Y.-W.; Belzile, N.; Gunn, J.M. Antagonistic effect of selenium on mercury assimilation by fish populations near Sudbury metal smelters? Am. Soc. Limnol. Oceanogr. 2001, 46, 1814–1818. [Google Scholar] [CrossRef]
- Deng, D.-F.; Teh, F.-C.; Teh, S.J. Effect of dietary methylmercury and seleno-methionine on Sacramento splittail larvae. Sci. Total Environ. 2008, 407, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Fung, L.A.H.; Antoine, J.M.R.; Grant, C.N.; Buddo, D.S.A. Evaluation of dietary exposure to minerals, trace elements and heavy metals from the muscle tissue of the lionfish Pterois volitans (Linnaeus 1758). Food Chem. Toxicol. 2013, 60, 205–212. [Google Scholar] [CrossRef]
- Kaneko, J.J.; Ralston, N.V.C. Selenium and mercury in pelagic fish in the central North Pacific near Hawaii. Biol. Trace Elem. Res. 2007, 119, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.A.; Ralston, N.V.C.; Peck, D.V.; Van Sickle, J.; Robertson, J.D.; Spate, V.L.; Morris, J.S. How might selenium moderate the toxic effects of mercury in stream fish of the western U.S.? Environ. Sci. Technol. 2009, 43, 3919–3925. [Google Scholar] [CrossRef]
- Ralston, N.V.C.; Ralston, C.R.; Blackwell, J.L.; Raymond, L.J. Dietary and tissue selenium in relation to methylmercury toxicity. Neurotoxicology 2008, 29, 802–811. [Google Scholar] [CrossRef] [Green Version]
- Raymond, L.J.; Ralston, N.V.C. Selenium’s importance in regulatory issues regarding mercury. Fuel Process. Technol. 2009, 90, 1333–1338. [Google Scholar] [CrossRef]
- Barone, G.; Storelli, A.; Mallamaci, R.; Storelli, M.M. Comparative Study on Trace Metal Accumulation in Liver of Mediterranean Deep-Sea Fish and Their Selenium/Mercury Molar Ratios. Water Air Soil Pollut. 2017, 228, 211. [Google Scholar] [CrossRef]
- Gochfeld, M.; Burger, J. Mercury interactions with selenium and sulfur and the relevance of the Se:Hg molar ratio to fish consumption advice. Environ. Sci. Pollut. Res. 2021, 28, 18407–18420. [Google Scholar] [CrossRef] [PubMed]
- Scheuhammer, A.; Wong, A.H.K.; Bond, D. Mercury and selenium accumulation in common loons (Gavia immer) and common mergansers (Mergus merganser) from eastern Canada. Environ. Toxicol. Chem. 1998, 17, 197–201. [Google Scholar] [CrossRef]
- Yamashita, Y.; Yamashita, M.; Iida, H. Selenium content in seafood in Japan. Nutrients 2013, 5, 388–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, X.; Wang, W.; Cheng, J.; Yuan, T.; Zhao, X.; Zhuang, H.; Qu, L. Free radicals and antioxidant status in rat liver after dietary exposure of environmental mercury. Environ. Toxicol. Pharmacol. 2006, 22, 309–314. [Google Scholar] [CrossRef]
- Grotto, D.; Valentini, J.; Serpeloni, J.M.; Monteiro, P.A.P.; Latorraca, E.F.; de Oliveira, R.S.; Antunes, L.M.G.; Garcia, S.C.; Barbosa, F.J. Evaluation of toxic effects of a diet containing fish contaminated with methylmercury in rats mimicking the exposure in the Amazon riverside population. Environ. Res. 2011, 111, 1074–1082. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.K.; Wang, F. Mercury-selenium compounds and their toxicological significance: Toward a molecular understanding of the mercury-selenium antagonism. Environ. Toxicol. Chem. 2009, 28, 1567–1577. [Google Scholar] [CrossRef] [PubMed]
- Olsvik, P.A.; Amlund, H.; Sæle, Ø.; Ellingsen, S.; Skjaerven, K.H. Impact of dietary selenium on methylmercury toxicity in juvenile Atlantic cod: A transcriptional survey. Chemosphere 2015, 120, 199–205. [Google Scholar] [CrossRef]
- Armstrong, R.N. Structure, catalytic mechanism, and evolution of the glutathione transferases. Chem. Res. Toxicol. 1997, 10, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Elia, A.C.; Galarini, R.; Taticchi, M.I.; Dörr, A.J.M.; Mantilacci, L. Antioxidant responses and bioaccumulation in Ictalurus melas under mercury exposure. Ecotoxicol. Environ. Saf. 2003, 55, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Amiard, J.-C.; Amiard-Triquet, C.; Barka, S.; Pellerin, J.; Rainbow, P.S. Metallothioneins in aquatic invertebrates: Their role in metal detoxification and their use as biomarkers. Aquat. Toxicol. 2006, 76, 160–202. [Google Scholar] [CrossRef] [PubMed]
- do Nascimento, J.L.M.; Oliveira, K.R.M.; Crespo-Lopez, M.E.; Macchi, B.M.; Maués, L.A.L.; Pinheiro, M.D.C.N.; Silveira, L.C.L.; Herculano, A.M. Methylmercury neurotoxicity and antioxidant defenses. Indian J. Med. Res. 2008, 128, 373–382. [Google Scholar]
- Hamilton, P.B.; Rolshausen, G.; Uren Webster, T.M.; Tyler, C.R. Adaptive capabilities and fitness consequences associated with pollution exposure in fish. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoliar, O.B.; Lushchak, V.I. Environmental Pollution and Oxidative Stress in Fish. In Oxidative Stress—Environmental Induction and Dietary Antioxidants; Lushchak, V.I., Ed.; IntechOpen: Rijeka, Croatia, 2012; pp. 131–166. [Google Scholar]
- Birnie-Gauvin, K.; Costantini, D.; Cooke, S.J.; Willmore, W.G. A comparative and evolutionary approach to oxidative stress in fish: A review. Fish Fish. 2017, 18, 928–942. [Google Scholar] [CrossRef]
- McFarland, V.A.; Inouye, L.S.; Lutz, C.H.; Jarvis, A.S.; Clarke, J.U.; McCant, D.D. Biomarkers of oxidative stress and genotoxicity in livers of field- collected brown bullhead, Ameiurus nebulosus. Arch. Environ. Contam. Toxicol. 1999, 37, 236–241. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Direction | Primer (5′–3′) | Accession Number |
|---|---|---|---|
| Glutathione s-transferase | Forward | GACTTCCCAGAATGGATGAAGG | BT07989.1 |
| Reverse | TGACTGAAACAGGACCAAATCA | ||
| Metallothionein | Forward | CTGGATCTTGCAACTGTGGT | X59392.1 |
| Reverse | CTTGCTGCAACCAGAAGGA | ||
| Superoxide dismutase | Forward | CGCAGAGGACAAGTACAAAGA | BT079033.1 |
| Reverse | GATGTGGCCTCCTCCATTAAA | ||
| Catalase | Forward | GTGGGAAAGACCACACCTATC | BT045615.1 |
| Reverse | GTTTCCCTCGTCAGTGTAGAAC | ||
| Ubiquitin | Forward | GCCTTTCCTACCTGACAGTATTC | BT079424.1 |
| Reverse | AAAGTCAACGCTCCATCTCC |
| Park | n | Wet Weight (kg) | Total Length (mm) | Se (µg/g Dry Weight) | MeHg (µg/g Dry Weight) | THg (µg/g Dry Weight) | THg:Se Molar Ratio |
|---|---|---|---|---|---|---|---|
| ISRO | 25 | 1.24 ± 0.49 | 572 ± 87 | 6.345 ± 2.299 | 0.311 ± 0.473 | 1.034 ± 1.414 | 0.064 ± 0.0496 |
| PIRO | 26 | 1.66 ± 1.27 | 586 ± 147 | 4.935 ± 1.176 | 0.490 ± 0.366 | 0.852 ± 0.614 | 0.068 ± 0.050 |
| SLBE | 8 | 0.77 ± 0.27 | 493 ± 53 | 6.156 ± 1.715 | 0.196 ± 0.110 | 0.360 ± 0.113 | 0.023 ± 0.022 |
| VOYA | 35 | 0.86 ± 0.42 | 527 ± 94 | 6.371 ± 2.787 | 0.177 ± 1.692 | 2.922 ± 3.146 | 0.181 ± 0.117 |
| sod Expression | cat Expression | mt Expression | gst Expression | Se (µmol/g dry wt) | MeHg (µmol/g dry wt) | THg (µmol/g dry wt) | |
|---|---|---|---|---|---|---|---|
| sod expression | 1 | ||||||
| cat expression | 0.698 (0.000) | 1 | |||||
| mt expression | 0.311 (0.002) | 0.377 (0.000) | 1 | ||||
| gst expression | 0.702 (0.000) | 0.503 (0.000) | 0.229 (0.026) | 1 | |||
| Se (µmol/g dry wt) | −0.157 (0.130) | −0.092 (0.385) | 0.197 (0.057) | −0.122 (0.240) | 1 | ||
| MeHg (µmol/g dry wt) | 0.021 (0.839) | 0.114 (0.279) | −0.095 (0.364) | −0.102 (0.329) | 0.178 (0.085) | 1 | |
| THg (µmol/g dry wt) | −0.057 (0.586) | 0.040 (0.702) | −0.065 (0.534) | −0.141 (0.175) | 0.347 (0.001) | 0.919 (0.000) | 1 |
| Park | ||||
|---|---|---|---|---|
| Gene Expressed | ISRO | PIRO | SLBE | VOYA |
| sod | 0.0003–0.786 n = 25 (0.281 ± 0.205) | 0.057–0.950 n = 26 (0.450 ± 0.259) | 0.074–2.189 n = 8 (0.671–0.716) | 0.004–1.168 n = 35 (0.352 ± 0.279) |
| cat | 0.0006–4.015 n = 24 (1.480 ± 0.925) | 0.006–8.598 n = 26 (2.896 ± 2.226) | 0.354–7.616 n = 8 (2.430 ± 2.208) | 0.002–9.109 n = 34 (2.898 ± 2.535) |
| mt | 0.142–61.958 n = 25 (24.463 ± 17.326) | 0.133–74.851 n = 26 (22.279 ± 18.595) | 23.095–85.840 n = 8 (47.120 ± 20.385) | 0.036–87.455 n = 35 (24.111 ± 21.974) |
| gst | 0.003–1.637 n = 25 (0.315 ± 0.348) | 0.048–0.897 n = 26 (0.261 ± 0.222) | 0.169–0.973 n = 8 (0.367 ± 0.266) | 0.004–0.899 n = 35 (0.233 ± 0.235) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magnuson, J.T.; Sandheinrich, M.B. Relation among Mercury, Selenium, and Biomarkers of Oxidative Stress in Northern Pike (Esox lucius). Toxics 2023, 11, 244. https://doi.org/10.3390/toxics11030244
Magnuson JT, Sandheinrich MB. Relation among Mercury, Selenium, and Biomarkers of Oxidative Stress in Northern Pike (Esox lucius). Toxics. 2023; 11(3):244. https://doi.org/10.3390/toxics11030244
Chicago/Turabian StyleMagnuson, Jason T., and Mark B. Sandheinrich. 2023. "Relation among Mercury, Selenium, and Biomarkers of Oxidative Stress in Northern Pike (Esox lucius)" Toxics 11, no. 3: 244. https://doi.org/10.3390/toxics11030244
APA StyleMagnuson, J. T., & Sandheinrich, M. B. (2023). Relation among Mercury, Selenium, and Biomarkers of Oxidative Stress in Northern Pike (Esox lucius). Toxics, 11(3), 244. https://doi.org/10.3390/toxics11030244