
Exopolysaccharides-Mediated ZnO Nanoparticles for the Treatment of Aquatic Diseases in Freshwater Fish Oreochromis mossambicus


 ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Experimental Animal
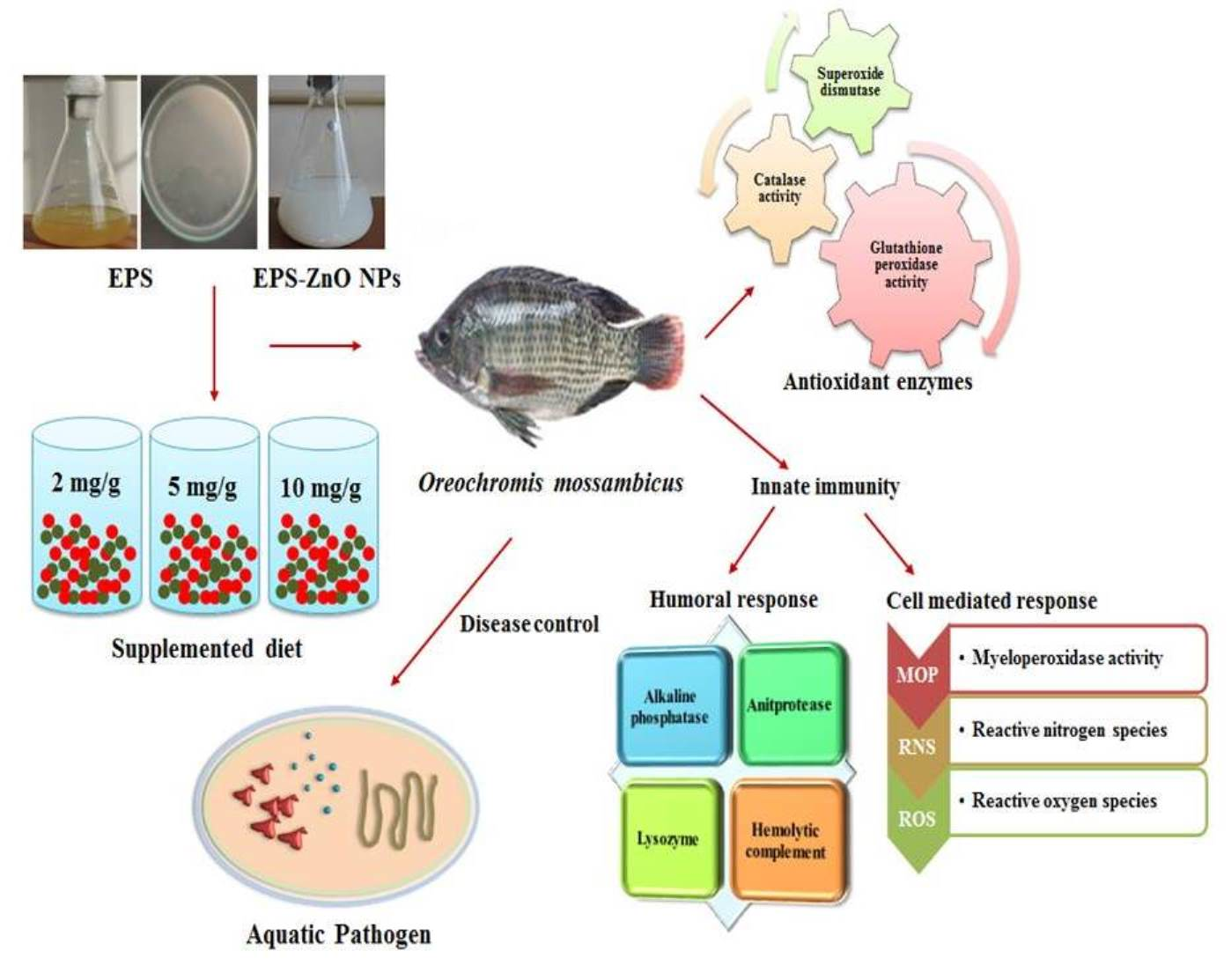
2.2. Methods for EPS Extraction and EPS-ZnO NP Synthesis
2.3. Experimental Diet Formulation
2.4. Experimental Design and Sample Collection
2.4.1. Liver Tissue Collection
2.4.2. Blood and Serum Collection
2.4.3. Skin Mucus Collection
2.5. Growth Performance
2.6. Immune Response
2.6.1. Cellular-Immunological Parameters
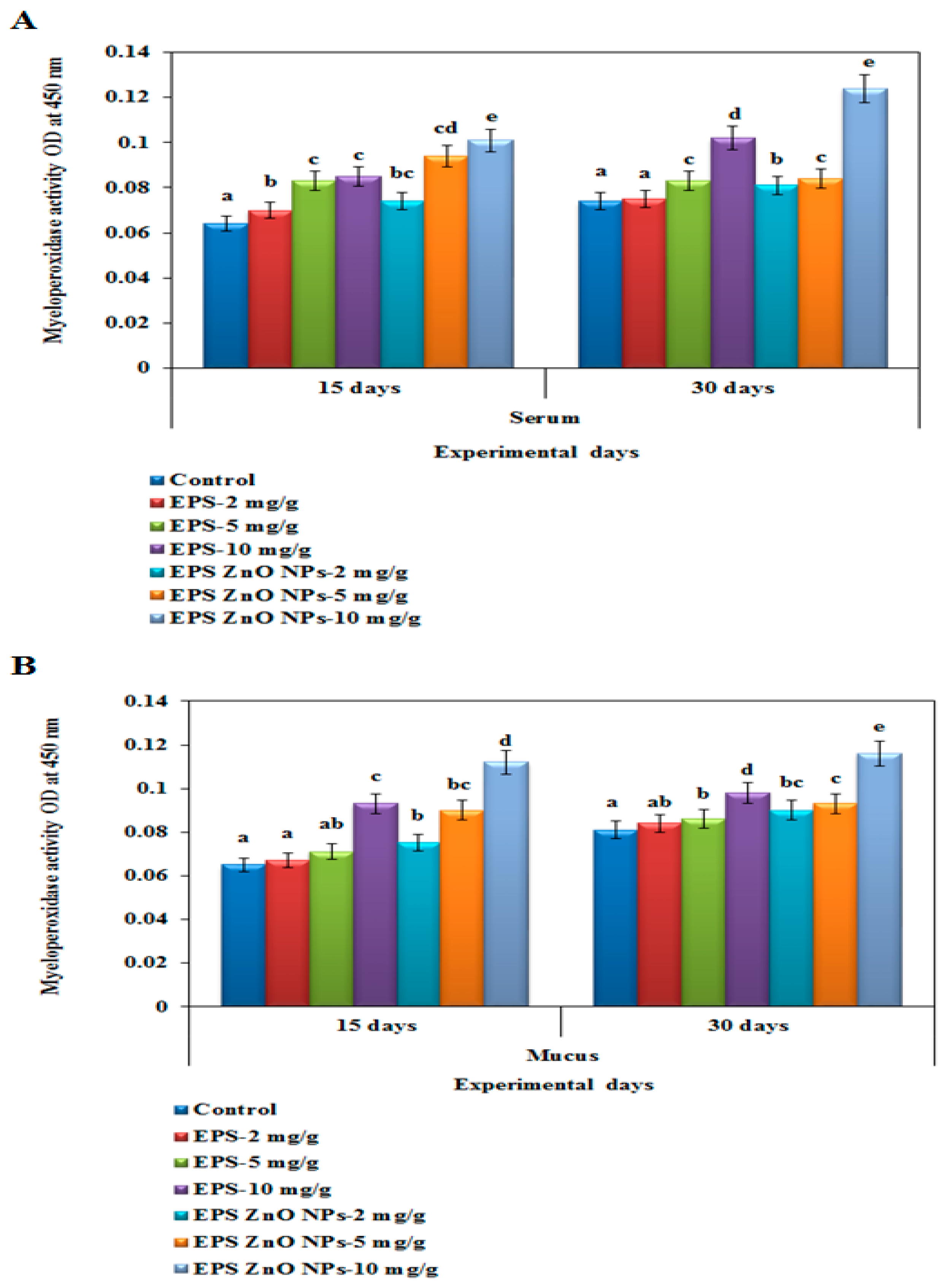
Myeloperoxidase (MPO) Activity
Reactive Oxygen Species (ROS) Activity
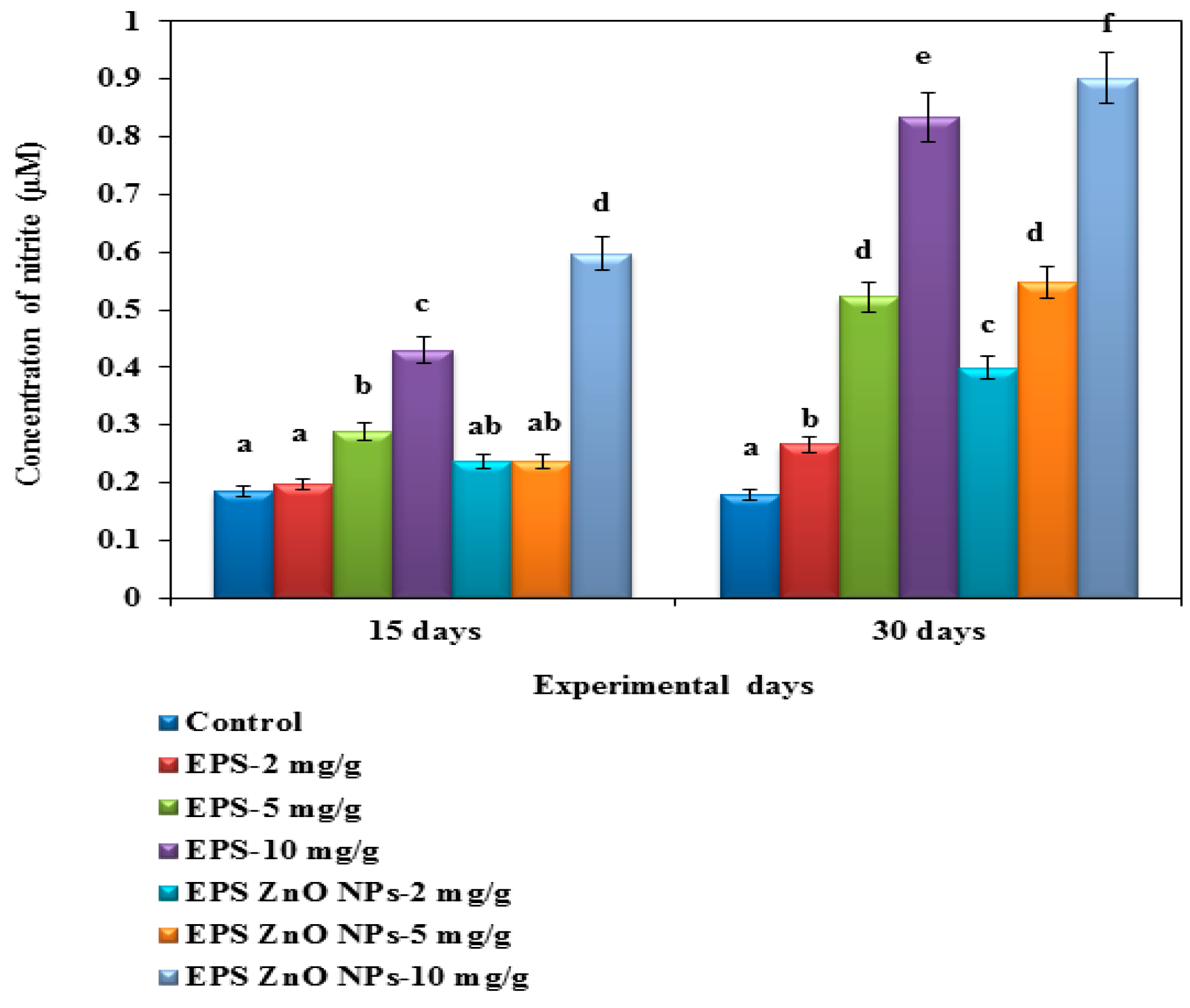
Reactive Nitrogen Species (RNS) Activity
2.6.2. Humoral-Immunological Parameters
Antiprotease Activity
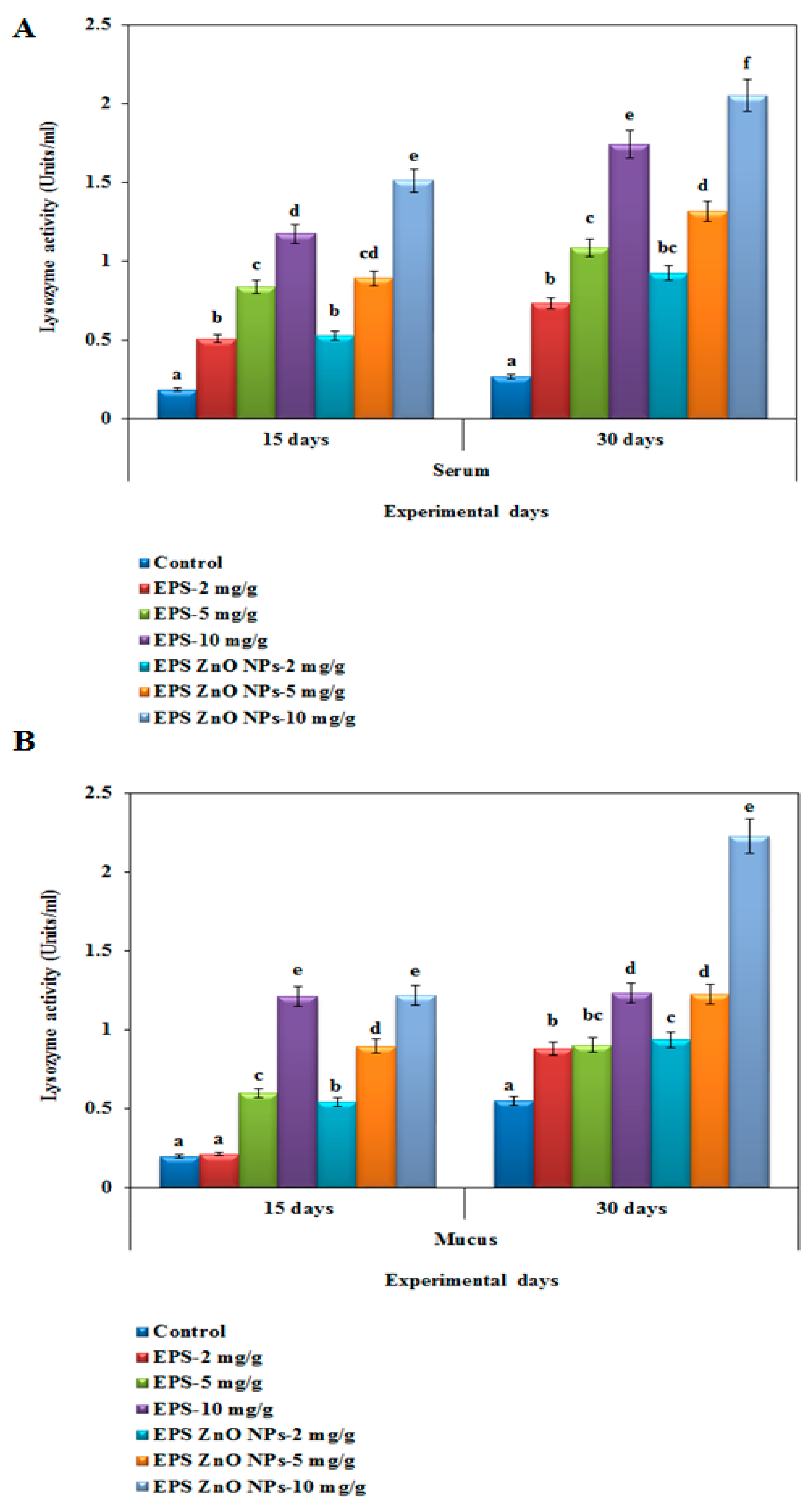
Lysozyme-LYZ Activity
Alkaline Phosphatase-ALP Activity
Hemolytic Complement Activity
2.7. Antioxidant Enzymes Activity
2.7.1. Superoxide Dismutase Activity
2.7.2. Catalase Activity
2.7.3. Glutathione Peroxidase Activity
2.8. Challenge Test with A. hydrophila and V. parahaemolyticus
2.9. Statistical Analyses
3. Results
3.1. Growth Performance
3.2. Immune Response
3.2.1. Cellular Immunological Parameters
Myeloperoxidase-MOP Activity
ROS and RNS Activity
3.2.2. Humoral Immunological Parameters
Antiprotease Activity
Lysozyme-LYZ Activity
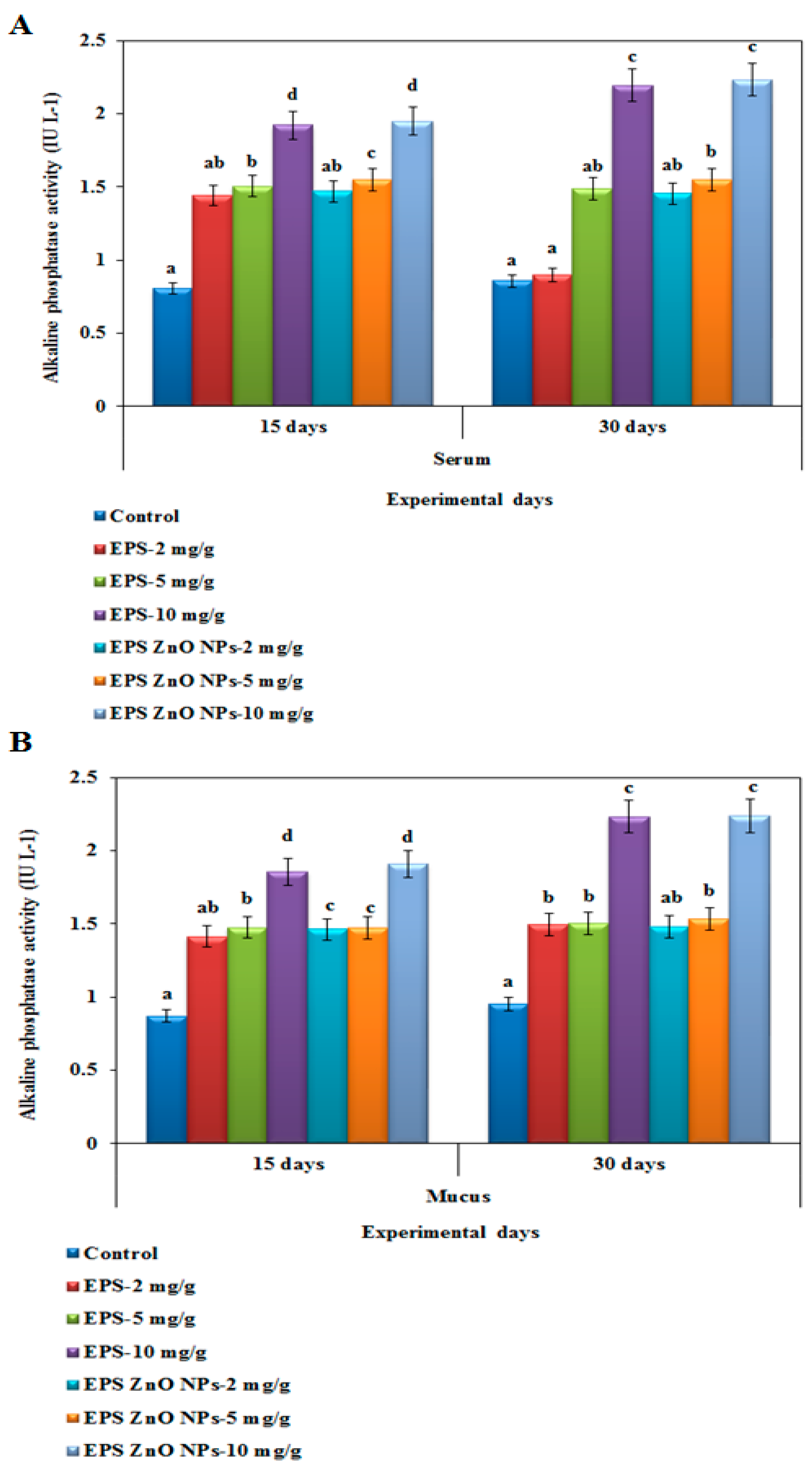
Alkaline Phosphatase-ALP Activity
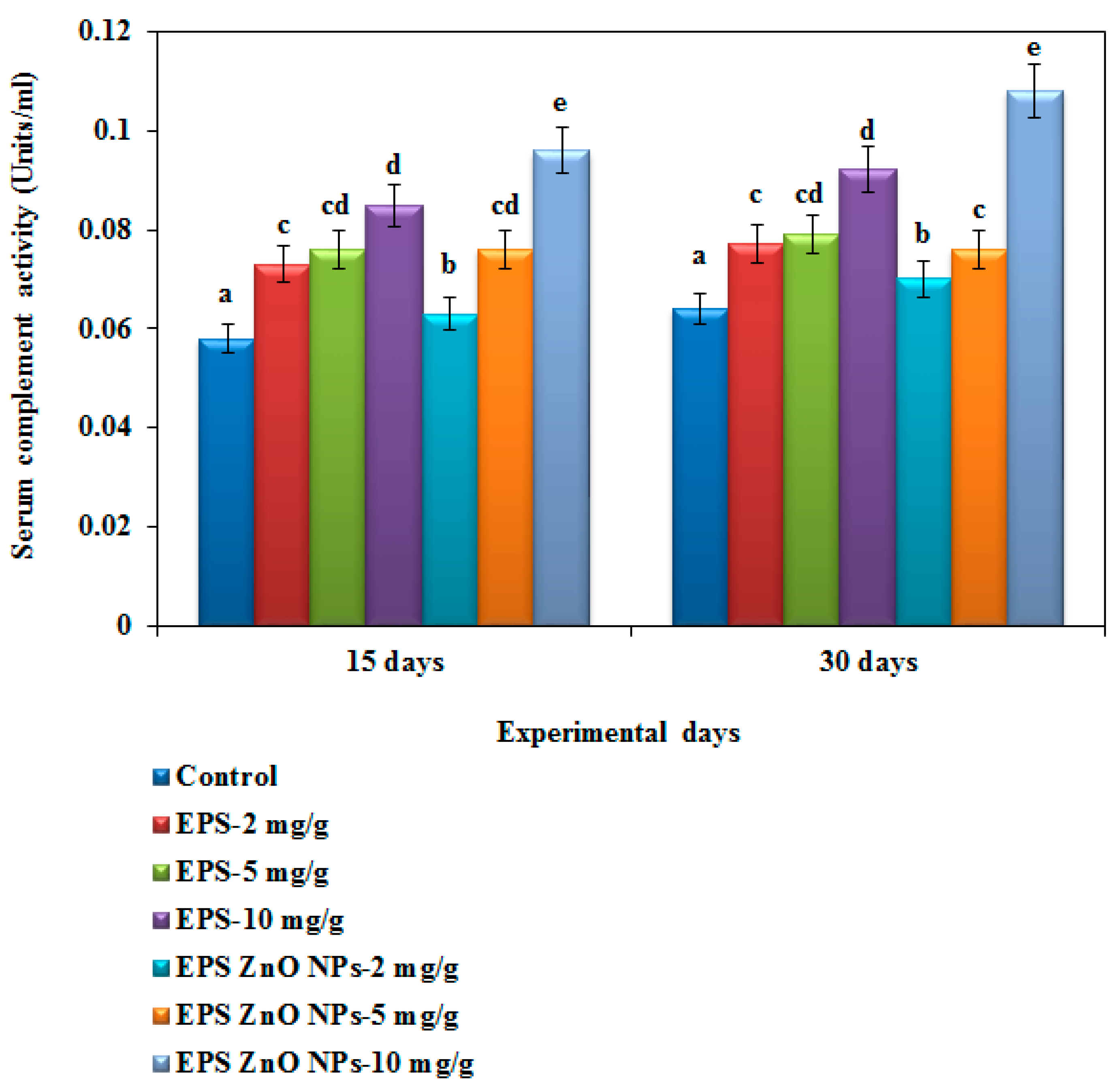
Hemolytic Complement Activity
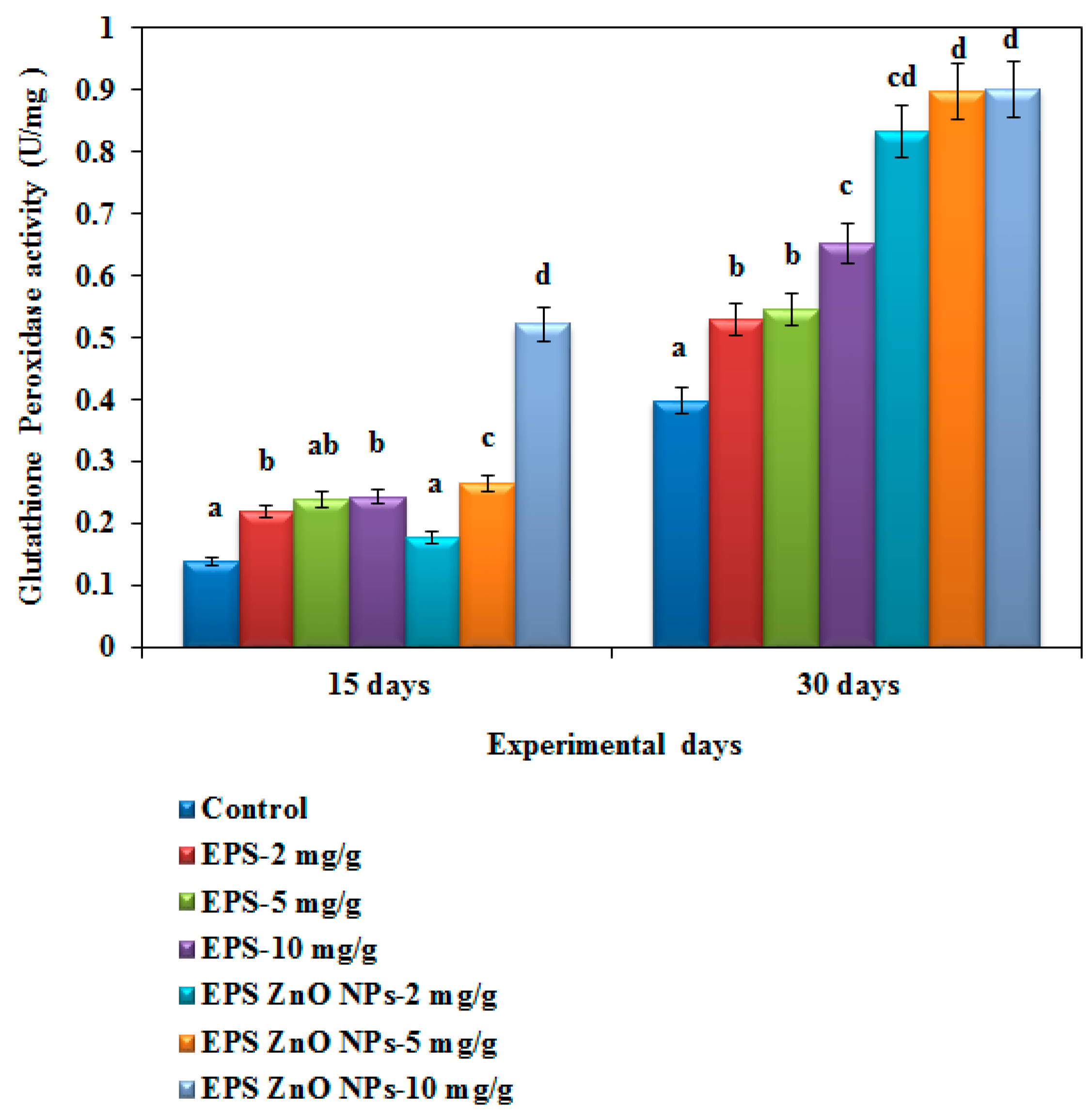
3.3. Antioxidant Enzyme Activity
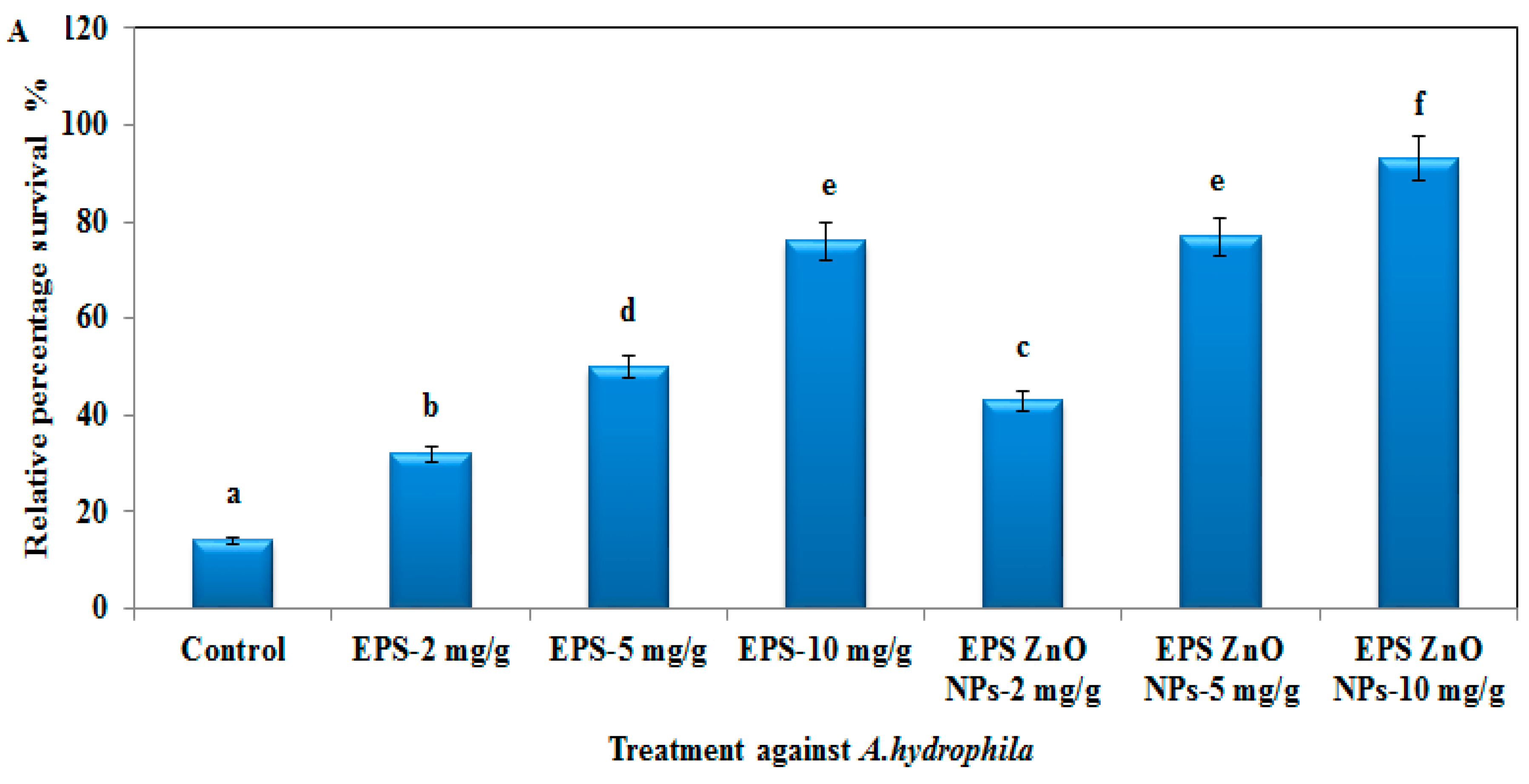
3.4. Challenge Test with A. hydrophila and V. parahaemolyticus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, J.L.; Meng, X.L.; Lu, R.H.; Wu, C.; Luo, Y.T.; Yan, X.; Li, X.J.; Kong, X.H.; Nie, G.X. Effects of Rehmannia glutinosa on growth performance, immunological parameters and disease resistance to Aeromonas hydrophila in common carp (Cyprinus carpio L.). Aquaculture 2015, 435, 293–300. [Google Scholar] [CrossRef]
- Aboyadak, I.M.; Sabry, N.M.; Ali, N.G.; El-Sayed, H.S. Isolation of Staphylococcus epidermidis, Bacillus cereus and Pseudomonas stutzeri from diseased European sea bass (Dicentrarchus labrax) for the first time in Egypt. Egypt. J. Aquat. Biol. Fish. 2016, 20, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Gallage, S.; Katagiri, T.; Endo, M.; Maita, M. Comprehensive evaluation of immunomodulation by moderate hypoxia in S. agalactiae vaccinated Nile tilapia. Fish Shellfish Immunol. 2017, 66, 445–454. [Google Scholar] [CrossRef] [PubMed]
- FAO. Global Aquaculture Production; United Nations Fisheries and Aquaculture Department: Rome, Italy, 2018. [Google Scholar]
- Austin, B.; Austin, D.A. Vibrionaceae representatives. In Bacterial Fish Pathogens; Springer: Dordrecht, The Netherlands, 2012; pp. 357–411. [Google Scholar] [CrossRef]
- Shoemaker, C.; Xu, D.H.; LaFrentz, B.; LaPatra, S. Overview of fish immune system and infectious diseases. In Dietary Nutrients, Additives and Fish Health; Wiley: Toronto, ON, Canada, 2015; pp. 1–24. [Google Scholar] [CrossRef]
- Huang, L.; Qin, Y.; Yan, Q.; Lin, G.; Huang, L.; Huang, B.; Huang, W. MinD plays an important role in Aeromonas hydrophila adherence to Anguilla japonica mucus. Gene 2015, 565, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Sun, J.; Liu, C.; Xue, Z. Application of immunostimulants in aquaculture: Current knowledge and future perspectives. Aquac. Res. 2017, 48, 1–23. [Google Scholar] [CrossRef]
- Elsheshtawy, A.; Yehia, N.; Elkemary, M.; Soliman, H. Direct detection of unamplified Aeromonas hydrophila DNA in clinical fish samples using gold nanoparticle probe-based assay. Aquaculture 2019, 500, 451–457. [Google Scholar] [CrossRef]
- Li, H.; Tian, X.; Zhao, K.; Jiang, W.; Dong, S. Effect of Clostridium butyricum in different forms on growth performance, disease resistance, expression of genes involved in immune responses and mTOR signaling pathway of Litopenaeus vannamai. Fish Shellfish Immunol. 2019, 87, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Alderman, D.J.; Hastings, T.S. Antibiotic use in aquaculture: Development of antibiotic resistance–potential for consumer health risks. Int. J. Food Sci. Technol. 1998, 33, 139–155. [Google Scholar] [CrossRef]
- Teuber, M. Veterinary use and antibiotic resistance. Curr. Opin. Microbiol. 2001, 4, 493–499. [Google Scholar] [CrossRef]
- Xiong, W.; Sun, Y.; Zhang, T.; Ding, X.; Li, Y.; Wang, M.; Zeng, Z. Antibiotics, antibiotic resistance genes, and bacterial community composition in fresh water aquaculture environment in China. Microb. Ecol. 2015, 70, 425–432. [Google Scholar] [CrossRef]
- Liu, X.; Steele, J.C.; Meng, X.Z. Usage, residue, and human health risk of antibiotics in Chinese aquaculture: A review. Environ. Pollut. 2017, 223, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.C.; Takaoka, O.; Jeong, G.S.; Lee, S.W.; Ishimaru, K.; Seoka, M.; Takii, K. Dietary medicinal herbs improve growth and some non-specific immunity of red sea bream Pagrus major. Fish Sci. 2007, 73, 63–69. [Google Scholar] [CrossRef]
- Bulfon, C.; Volpatti, D.; Galeotti, M. Current research on the use of plant-derived products in farmed fish. Aquac. Res. 2015, 46, 513–551. [Google Scholar] [CrossRef]
- Gobi, N.; Ramya, C.; Vaseeharan, B.; Malaikozhundan, B.; Vijayakumar, S.; Murugan, K.; Benelli, G. Oreochromis mossambicus diet supplementation with Psidium guajava leaf extracts enhance growth, immune, antioxidant response and resistance to Aeromonas hydrophila. Fish Shellfish Immunol. 2016, 58, 572–583. [Google Scholar] [CrossRef] [PubMed]
- Gobi, N.; Vaseeharan, B.; Chen, J.C.; Rekha, R.; Vijayakumar, S.; Anjugam, M.; Iswarya, A. Dietary supplementation of probiotic Bacillus licheniformis Dahb1 improves growth performance, mucus and serum immune parameters, antioxidant enzyme activity as well as resistance against Aeromonas hydrophila in tilapia Oreochromis mossambicus. Fish Shellfish Immunol. 2018, 74, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Talpur, A.D.; Ikhwanuddin, M.H.D. Dietary effects of garlic (Allium sativum) on haemato-immunological parameters, survival, growth, and disease resistance against Vibrio harveyi infection in Asian sea bass, Lates calcarifer (Bloch). Aquaculture 2012, 364, 6–12. [Google Scholar] [CrossRef]
- Dimapilis, E.A.S.; Hsu, C.S.; Mendoza, R.M.O.; Lu, M.C. Zinc oxide nanoparticles for water disinfection. Sustain. Environ. Res. 2018, 28, 47–56. [Google Scholar] [CrossRef]
- Heijerick, D.G.; de Schamphelaere, K.A.; Janssen, C.R. Predicting acute zinc toxicity for Daphnia magna as a function of key water chemistry characteristics: Development and validation of a biotic ligand model. Environ. Toxicol. Chem. 2002, 21, 1309–1315. [Google Scholar] [CrossRef]
- Wekell, J.C.; Shearer, K.D.; Gauglitz, E.J., Jr. Zinc supplementation of trout diets: Tissue indicators of body zinc status. Prog. Fish. Cult. 1986, 48, 205–212. [Google Scholar] [CrossRef]
- Swain, P.S.; Rao, S.B.; Rajendran, D.; Dominic, G.; Selvaraju, S. Nano zinc, an alternative to conventional zinc as animal feed supplement: A review. Anim. Nutr. 2016, 2, 134–141. [Google Scholar] [CrossRef]
- Nasr-Eldahan, S.; Nabil-Adam, A.; Shreadah, M.A.; Maher, A.M.; El-Sayed Ali, T. A review article on nanotechnology in aquaculture sustainability as a novel tool in fish disease control. Aquac. Int. 2021, 29, 1459–1480. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, M.A.; Evans, C.W.; Raston, C.L. Green chemistry and the health implications of nanoparticles. Green Chem. 2006, 8, 417–432. [Google Scholar] [CrossRef]
- Rajendran, D. Application of nano minerals in animal production system. Res. J. Biotechnol. 2013, 8, 1–3. [Google Scholar]
- Welman, A.D.; Maddox, I.S. Exopolysaccharides from lactic acid bacteria: Perspectives and challenges. Trends Biotechnol. 2003, 21, 269–274. [Google Scholar] [CrossRef]
- Marudhupandi, T.; Inbakandan, D. Polysaccharides in aquatic disease management. Fish. Aquac. J. 2015, 6, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Gobi, N.; Malaikozhundan, B.; Sekar, V.; Shanthi, S.; Vaseeharan, B.; Jayakumar, R.; Nazar, A.K. GFP tagged Vibrio parahaemolyticus Dahv2 infection and the protective effects of the probiotic Bacillus licheniformis Dahb1 on the growth, immune and antioxidant responses in Pangasius hypophthalmus. Fish Shellfish Immunol. 2016, 52, 230–238. [Google Scholar] [CrossRef]
- Abinaya, M.; Vaseeharan, B.; Divya, M.; Vijayakumar, S.; Govindarajan, M.; Alharbi, N.S.; Khaled, J.M.; Al-Anbr, M.N.; Benelli, G. Structural characterization of Bacillus licheniformis Dahb1 exopolysaccharide—Antimicrobial potential and larvicidal activity on malaria and Zika virus mosquito vectors. Environ. Sci. Pollut. Res. 2018, 25, 18604–18619. [Google Scholar] [CrossRef]
- Abinaya, M.; Vaseeharan, B.; Divya, M.; Sharmili, A.; Govindarajan, M.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Benelli, G. Bacterial exopolysaccharide (EPS)-coated ZnO nanoparticles showed high antibiofilm activity and larvicidal toxicity against malaria and Zika virus vectors. J. Trace Elem. Med. Biol. 2018, 45, 93–103. [Google Scholar] [CrossRef]
- Pakravan, S.; Hajimoradloo, A.; Ghorbani, R. Effect of dietary willow herb, Epilobium hirsutum extract on growth performance, body composition, haematological parameters and Aeromonas hydrophila challenge on common carp, Cyprinus carpio. Aquac. Res. 2012, 43, 861–869. [Google Scholar] [CrossRef]
- Jenkins, J.A.; Bart, H.L., Jr.; Bowker, J.D.; Bowser, P.R.; MacMillan, J.R.; Nickum, J.G. Guidelines for the Use of Fishes in Research; American Fisheries Society: Bethesda, MA, USA, 2014. [Google Scholar]
- Kumari, J.; Sahoo, P.K. Dietary β-1, 3 glucan potentiates innate immunity and disease resistance of Asian catfish, Clarias batrachus (L.). J. Fish Dis. 2006, 29, 95–101. [Google Scholar] [CrossRef]
- Secombes, C. Isolation of salmonid macrophages and analysis of their killing activity. Techn. Fish Immunol. 1990, 1, 137–154. [Google Scholar]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15N] nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Bowden, T.J.; Butler, R.; Bricknell, I.R.; Ellis, A.E. Serum trypsin-inhibitory activity in five species of farmed fish. Fish Shellfish Immunol. 1997, 7, 377–385. [Google Scholar] [CrossRef]
- Ellis, A.I. Lysozyme assays. Techn. Fish Immunol. 1990, 1, 101–103. [Google Scholar]
- Sanchooli, O.; Hajimoradloo, A.; Ghorbani, R. Measurement of alkaline phosphatase and lysozyme enzymes in epidermal mucus of different weights of Cyprinus carpio. World J. Fish Mar. Sci. 2012, 4, 521–524. [Google Scholar] [CrossRef]
- Cuesta, A.; Rodríguez, A.; Esteban, M.A.; Meseguer, J. In vivo effects of propolis, a honeybee product, on gilthead seabream innate immune responses. Fish Shellfish Immunol. 2005, 18, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K. Measurement of Mn-SOD and Cu, Zn-SOD. Exp. Protoc. React. Oxyg. Nitrogen Species 2000, 1, 91–95. [Google Scholar]
- Takahara, S.; Hamilton, H.B.; Neel, J.V.; Kobara, T.Y.; Ogura, Y.; Nishimura, E.T. Hypocatalasemia: A new genetic carrier state. J. Clin. Investig. 1960, 39, 610–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotruck, J.T.; Pope, A.L.; Ganther, H.E.; Swanson, A.B.; Hafeman, D.G.; Hoekstra, W. Selenium: Biochemical role as a component of glutathione peroxidase. Science 1973, 179, 588–590. [Google Scholar] [CrossRef]
- Misra, C.K.; Das, B.K.; Mukherjee, S.C.; Meher, P.K. The immunomodulatory effects of tuftsin on the non-specific immune system of Indian Major carp, Labeo rohita. Fish Shellfish Immunol. 2006, 20, 728–738. [Google Scholar] [CrossRef]
- Dawood, M.A.; Koshio, S.; Esteban, M.Á. Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev Aquac. 2018, 10, 950–974. [Google Scholar] [CrossRef]
- Reverter, M.; Bontemps, N.; Lecchini, D.; Banaigs, B.; Sasal, P. Use of plant extracts in fish aquaculture as an alternative to chemotherapy: Current status and future perspectives. Aquaculture 2014, 433, 50–61. [Google Scholar] [CrossRef]
- Shen, B. A new golden age of natural products drug discovery. Cell 2015, 163, 1297–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilen, S.; Ünal, S.; Güvensoy, H. Effects of oyster mushroom (Pleurotus ostreatus) and nettle (Urtica dioica) methanolic extracts on immune responses and resistance to Aeromonas hydrophila in rainbow trout (Oncorhynchus mykiss). Aquaculture 2016, 454, 90–94. [Google Scholar] [CrossRef]
- Yousefi, M.; Hoseini, S.M.; Vatnikov, Y.A.; Kulikov, E.V.; Drukovsky, S.G. Rosemary leaf powder improved growth performance, immune and antioxidant parameters, and crowding stress responses in common carp (Cyprinus carpio) fingerlings. Aquaculture 2019, 505, 473–480. [Google Scholar] [CrossRef]
- Zoral, M.A.; Futami, K.; Endo, M.; Maita, M.; Katagiri, T. Anthelmintic activity of Rosmarinus officinalis against Dactylogyrus minutus (Monogenea) infections in Cyprinus carpio. Vet. Parasitol. 2017, 247, 1–6. [Google Scholar] [CrossRef]
- Das, A.; Nakhro, K.; Chowdhury, S.; Kamilya, D. Effects of potential probiotic Bacillus amyloliquifaciens FPTB16 on systemic and cutaneous mucosal immune responses and disease resistance of catla (Catla catla). Fish Shellfish Immunol. 2013, 35, 1547–1553. [Google Scholar] [CrossRef]
- Salinas, I.; Abelli, L.; Bertoni, F.; Picchietti, S.; Roque, A.; Furones, D.; Cuesta, A.; Meseguer, J.; Esteban, M.Á. Monospecies and multispecies probiotic formulations produce different systemic and local immunostimulatory effects in the gilthead seabream (Sparus aurata L.). Fish Shellfish Immunol. 2008, 25, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Bairagi, A.; Ghosh, K.S.; Sen, S.K.; Ray, A.K. Enzyme producing bacterial flora isolated from fish digestive tracts. Aquac. Int. 2002, 10, 109–121. [Google Scholar] [CrossRef]
- Banerjee, G.; Ray, A.K.; Askarian, F.; Ringø, E. Characterisation and identification of enzyme-producing autochthonous bacteria from the gastrointestinal tract of two Indian air-breathing fish. Benef. Microbes 2013, 4, 277–284. [Google Scholar] [CrossRef]
- Mondal, S.; Roy, T.; Ray, A.K. Characterization and identification of enzyme-producing bacteria isolated from the digestive tract of bata, Labeo bata. J. World Aquac. Soc. 2010, 41, 369–377. [Google Scholar] [CrossRef]
- Ray, A.K.; Roy, T.; Mondal, S.; Ringø, E. Identification of gut-associated amylase, cellulase and protease-producing bacteria in three species of Indian major carps. Aquac. Res. 2010, 41, 1462–1469. [Google Scholar] [CrossRef]
- Nayak, S.K.; Swain, P.; Mukherjee, S.C. Effect of dietary supplementation of probiotic and vitamin C on the immune response of Indian major carp, Labeo rohita (Ham.). Fish Shellfish Immunol. 2007, 23, 892–896. [Google Scholar] [CrossRef]
- Giri, S.S.; Sukumaran, V.; Dangi, N.K. Characteristics of bacterial isolates from the gut of freshwater fish, Labeo rohita that may be useful as potential probiotic bacteria. Probiotics Antimicrob. Proteins 2012, 4, 238–242. [Google Scholar] [CrossRef]
- Aly, S.M.; Ahmed, Y.A.G.; Abdel-Aziz, A. Studies on Bacillus subtilis and Lactobacillus acidophilus, as potential probiotics, on the immune response and resistance of Tilapia nilotica (Oreochromis niloticus) to challenge infections. Fish Shellfish Immunol. 2008, 25, 128–136. [Google Scholar] [CrossRef]
- Harikrishnan, R.; Balasundaram, C.; Heo, M.S. Lactobacillus sakei BK19 enriched diet enhances the immunity status and disease resistance to streptococcosis infection in kelp grouper, Epinephelus bruneus. Fish Shellfish Immunol. 2010, 29, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Merrifield, D.L.; Bradley, G.; Baker, R.T.M.; Davies, S.J. Probiotic applications for rainbow trout (Oncorhynchus mykiss Walbaum) II. Effects on growth performance, feed utilization, intestinal microbiota and related health criteria postantibiotic treatment. Aquac. Nutr. 2010, 16, 496–503. [Google Scholar] [CrossRef]
- Azarin, H.; Aramli, M.S.; Imanpour, M.R.; Rajabpour, M. Effect of a probiotic containing Bacillus licheniformis and Bacillus subtilis and ferroin solution on growth performance, body composition and haematological parameters in Kutum (Rutilus frisii kutum) fry. Probiotics Antimicrob. Proteins 2015, 7, 31–37. [Google Scholar] [CrossRef]
- Ellis, A.E. Innate host defense mechanisms of fish against viruses and bacteria. Dev. Comp. Immunol. 2001, 25, 827–839. [Google Scholar] [CrossRef]
- Newaj-Fyzul, A.; Adesiyun, A.A.; Mutani, A.; Ramsubhag, A.; Brunt, J.; Austin, B. Bacillus subtilis AB1 controls Aeromonas infection in rainbow trout (Oncorhynchus mykiss, Walbaum). J. Appl. Microbiol. 2007, 103, 1699–1706. [Google Scholar] [CrossRef]
- Cerezuela, R.; Guardiola, F.A.; Meseguer, J.; Esteban, M.A. Enrichment of gilthead seabream (Sparus aurata L.) diet with microalgae: Effects on the immune system. Fish Physiol. Biochem. 2012, 38, 1729–1739. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.R.; Gong, Q.F.; Fang, H.; Liang, W.W.; Chen, M.; He, R.J. Effect of Sophora flavescens on non-specific immune response of tilapia (GIFT Oreochromis niloticus) and disease resistance against Streptococcus agalactiae. Fish Shellfish Immunol. 2013, 34, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Alexander, C.P.; Kirubakaran, C.J.W.; Michael, R.D. Water soluble fraction of Tinospora cordifolia leaves enhanced the non-specific immune mechanisms and disease resistance in Oreochromis mossambicus. Fish Shellfish Immunol. 2010, 29, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Christybapita, D.; Divyagnaneswari, M.; Michael, R.D. Oral administration of Eclipta alba leaf aqueous extract enhances the non-specific immune responses and disease resistance of Oreochromis mossambicus. Fish Shellfish Immunol. 2007, 23, 840–852. [Google Scholar] [CrossRef] [PubMed]
- Divyagnaneswari, M.; Christybapita, D.; Michael, R.D. Enhancement of nonspecific immunity and disease resistance in Oreochromis mossambicus by Solanum trilobatum leaf fractions. Fish Shellfish Immunol. 2007, 23, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Lie, Ø.; Evensen, Ø.; Sorensen, A.; Froysadal, E. Study on lysozyme activity in some fish species. Dis. Aquat. Org. 1989, 6, 1–5. [Google Scholar] [CrossRef]
- Panigrahi, A.; Kiron, V.; Kobayashi, T.; Puangkaew, J.; Satoh, S.; Sugita, H. Immune responses in rainbow trout Oncorhynchus mykiss induced by a potential probiotic bacteria Lactobacillus rhamnosus JCM 1136. Vet. Immunol. Immunopathol. 2004, 102, 379–388. [Google Scholar] [CrossRef]
- Taoka, Y.; Maeda, H.; Jo, J.Y.; Kim, S.M.; Park, S.I.; Yoshikawa, T.; Sakata, T. Use of live and dead probiotic cells in tilapia Oreochromis niloticus. Fish Sci. 2006, 72, 755–766. [Google Scholar] [CrossRef]
- Pratheepa, V.; Sukumaran, N. Effect of Euphorbia hirta plant leaf extract on immunostimulant response of Aeromonas hydrophila infected Cyprinus carpio. PeerJ 2014, 2, 671. [Google Scholar] [CrossRef] [Green Version]
- Rao, Y.V.; Das, B.K.; Jyotyrmayee, P.; Chakrabarti, R. Effect of Achyranthes aspera on the immunity and survival of Labeo rohita infected with Aeromonas hydrophila. Fish Shellfish Immunol. 2006, 20, 263–273. [Google Scholar] [CrossRef]
- Sharifuzzaman, S.M.; Austin, B. Influence of probiotic feeding duration on disease resistance and immune parameters in rainbow trout. Fish Shellfish Immunol. 2009, 27, 440–445. [Google Scholar] [CrossRef] [PubMed]
- Cordero, H.; Morcillo, P.; Meseguer, J.; Cuesta, A.; Esteban, M.Á. Effects of Shewanella putrefaciens on innate immunity and cytokine expression profile upon high stocking density of gilthead seabream specimens. Fish Shellfish Immunol. 2016, 51, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Harikrishnan, R.; Balasundaram, C.; Heo, M.S. Diet enriched with mushroom Phellinus linteus extract enhances the growth, innate immune response, and disease resistance of kelp grouper, Epinephelus bruneus against vibriosis. Fish Shellfish Immunol. 2011, 30, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Van Doan, H.; Hoseinifar, S.H.; Khanongnuch, C.; Kanpiengjai, A.; Unban, K.; Srichaiyo, S. Host-associated probiotics boosted mucosal and serum immunity, disease resistance and growth performance of Nile tilapia (Oreochromis niloticus). Aquaculture 2018, 491, 94–100. [Google Scholar] [CrossRef]
- Jinendiran, S.; Nathan, A.A.; Ramesh, D.; Vaseeharan, B.; Sivakumar, N. Modulation of innate immunity, expression of cytokine genes and disease resistance against Aeromonas hydrophila infection in goldfish (Carassius auratus) by dietary supplementation with Exiguobacterium acetylicum S01. Fish Shellfish Immunol. 2019, 84, 458–469. [Google Scholar] [CrossRef]
- Giri, S.S.; Sukumaran, V.; Oviya, M. Potential probiotic Lactobacillus plantarum VSG3 improves the growth, immunity, and disease resistance of tropical freshwater fish, Labeo rohita. Fish Shellfish Immunol. 2013, 34, 660–666. [Google Scholar] [CrossRef]
- Giri, S.S.; Sen, S.S.; Sukumaran, V. Effects of dietary supplementation of potential probiotic Pseudomonas aeruginosa VSG-2 on the innate immunity and disease resistance of tropical freshwater fish, Labeo rohita. Fish Shellfish Immunol. 2012, 32, 1135–1140. [Google Scholar] [CrossRef]
- Rodriguez-Estrada, U.; Satoh, S.; Haga, Y.; Fushimi, H.; Sweetman, J. Effects of inactivated Enterococcus faecalis and mannan oligosaccharide and their combination on growth, immunity, and disease protection in rainbow trout. N. Am. J. Aquac. 2013, 75, 416–428. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | 30 Days | ||||||
|---|---|---|---|---|---|---|---|
| Control | EPS-2 mg/g | EPS-5 mg/g | EPS-10 mg/g | EPS-ZnO NPs-2 mg/g | EPS-ZnO NPs-5 mg/g | EPS-ZnO NPs-10 mg/g | |
| Initial weight (g) | 7.69 ± 1.2 a | 7.72 ± 1.4 a | 7.70 ± 1.1 a | 7.75 ± 1.8 a | 7.68 ± 1.4 a | 7.73 ± 1.3 a | 7.67 ± 1.9 a |
| Final weight (g) | 17.50 ± 0.9 b | 22.73 ± 0.2 bc | 25.41 ± 0.4 bc | 27.25 ± 0.1 cd | 24.61 ± 0.3 cd | 26.80 ± 0.5 cd | 34.42 ± 1.8 d |
| Specific growth rate (SGR) | 1.56 ± 0.3 a | 2.12 ± 0.6 bc | 2.30 ± 0.2 bc | 2.40 ± 0.6 cd | 2.23 ± 0.5 bc | 2.36 ± 0.4 bc | 2.57 ± 0.8 d |
| Feed conversion ratio (FCR) | 1.32 ± 0.2 a | 1.22 ± 0.1 ab | 1.18 ± 0.3 ab | 1.03 ± 0.4 bc | 1.08 ± 0.3 bc | 0.93 ± 0.2 cd | 0.76 ± 0.1 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abinaya, M.; Shanthi, S.; Palmy, J.; Al-Ghanim, K.A.; Govindarajan, M.; Vaseeharan, B. Exopolysaccharides-Mediated ZnO Nanoparticles for the Treatment of Aquatic Diseases in Freshwater Fish Oreochromis mossambicus. Toxics 2023, 11, 313. https://doi.org/10.3390/toxics11040313
Abinaya M, Shanthi S, Palmy J, Al-Ghanim KA, Govindarajan M, Vaseeharan B. Exopolysaccharides-Mediated ZnO Nanoparticles for the Treatment of Aquatic Diseases in Freshwater Fish Oreochromis mossambicus. Toxics. 2023; 11(4):313. https://doi.org/10.3390/toxics11040313
Chicago/Turabian StyleAbinaya, Muthukumar, Sathappan Shanthi, Jesudasan Palmy, Khalid A. Al-Ghanim, Marimuthu Govindarajan, and Baskaralingam Vaseeharan. 2023. "Exopolysaccharides-Mediated ZnO Nanoparticles for the Treatment of Aquatic Diseases in Freshwater Fish Oreochromis mossambicus" Toxics 11, no. 4: 313. https://doi.org/10.3390/toxics11040313
APA StyleAbinaya, M., Shanthi, S., Palmy, J., Al-Ghanim, K. A., Govindarajan, M., & Vaseeharan, B. (2023). Exopolysaccharides-Mediated ZnO Nanoparticles for the Treatment of Aquatic Diseases in Freshwater Fish Oreochromis mossambicus. Toxics, 11(4), 313. https://doi.org/10.3390/toxics11040313