

Toxicological and Mutagenic Effects of Particulate Matter from Domestic Activities



 ,
,  ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. PM10 Sampling and Chemical Characterisation
2.2. Sample Preparation for Toxicological Assays
2.3. Cell Culture
2.4. Cell Viability Measurements
2.5. Analysis of Intracellular ROS
2.6. Cell Cycle Analysis
2.7. Mutagenicity Assay
2.8. Statistical Analysis
3. Results and Discussion
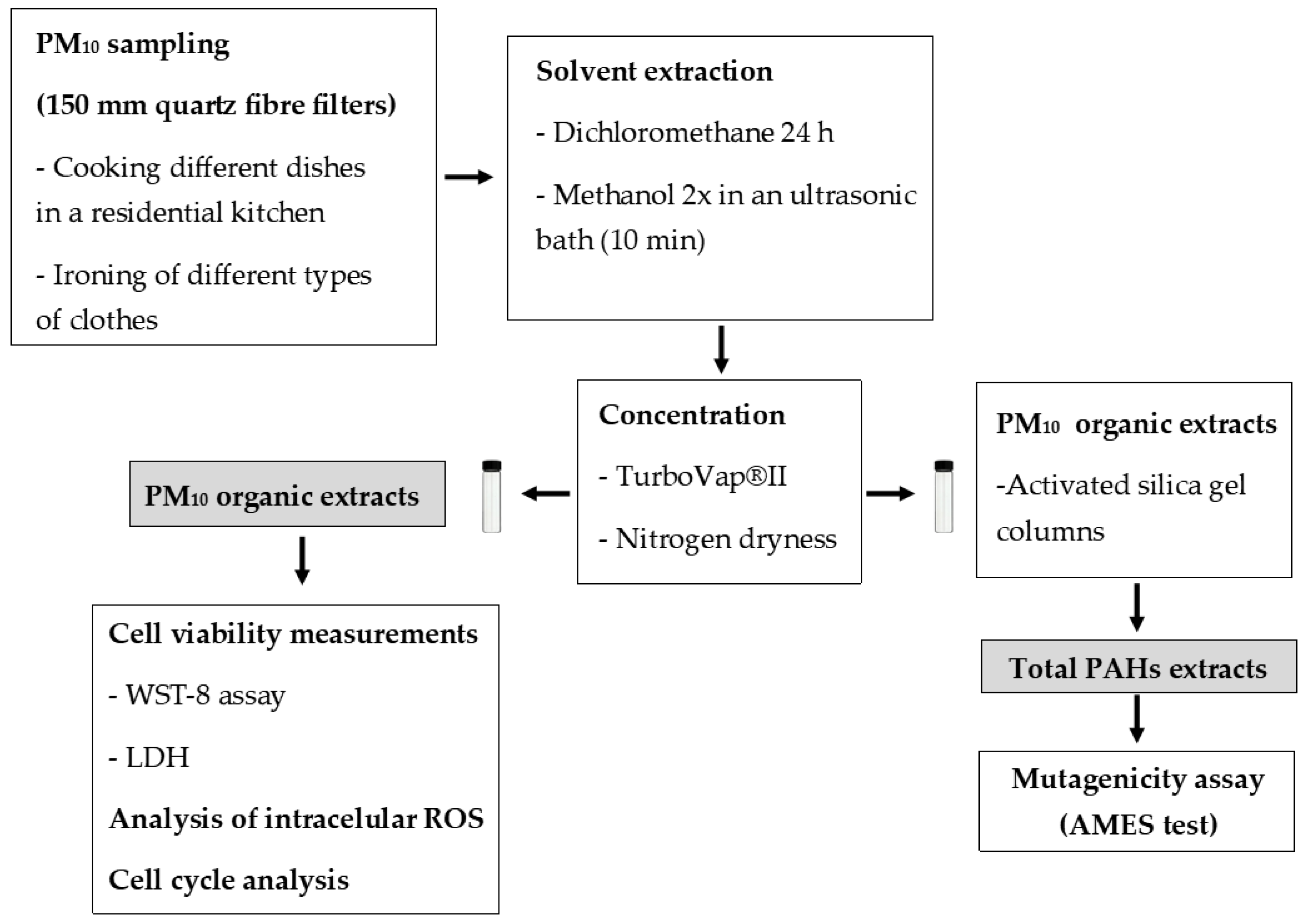
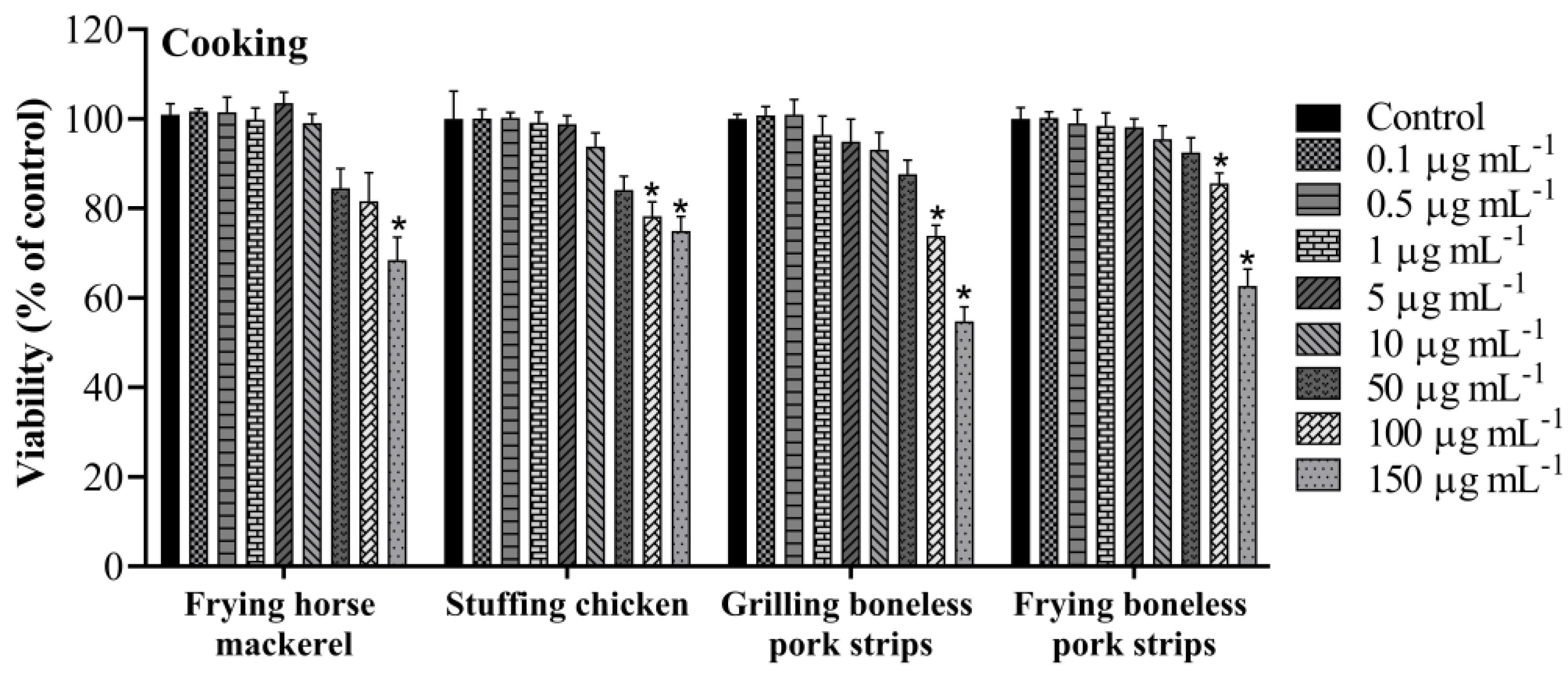
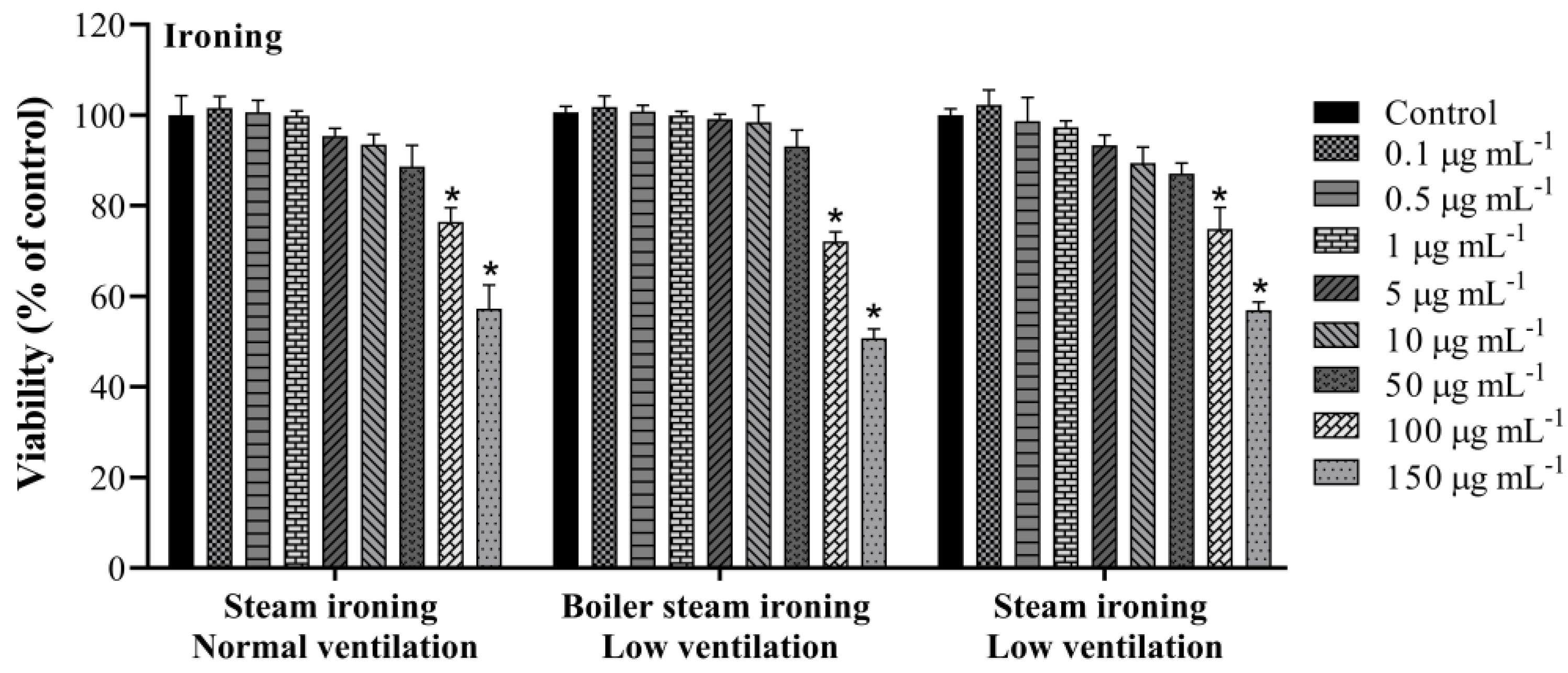
3.1. Cell Viability
3.2. Relationship between Chemical Components and Cytotoxic Effects
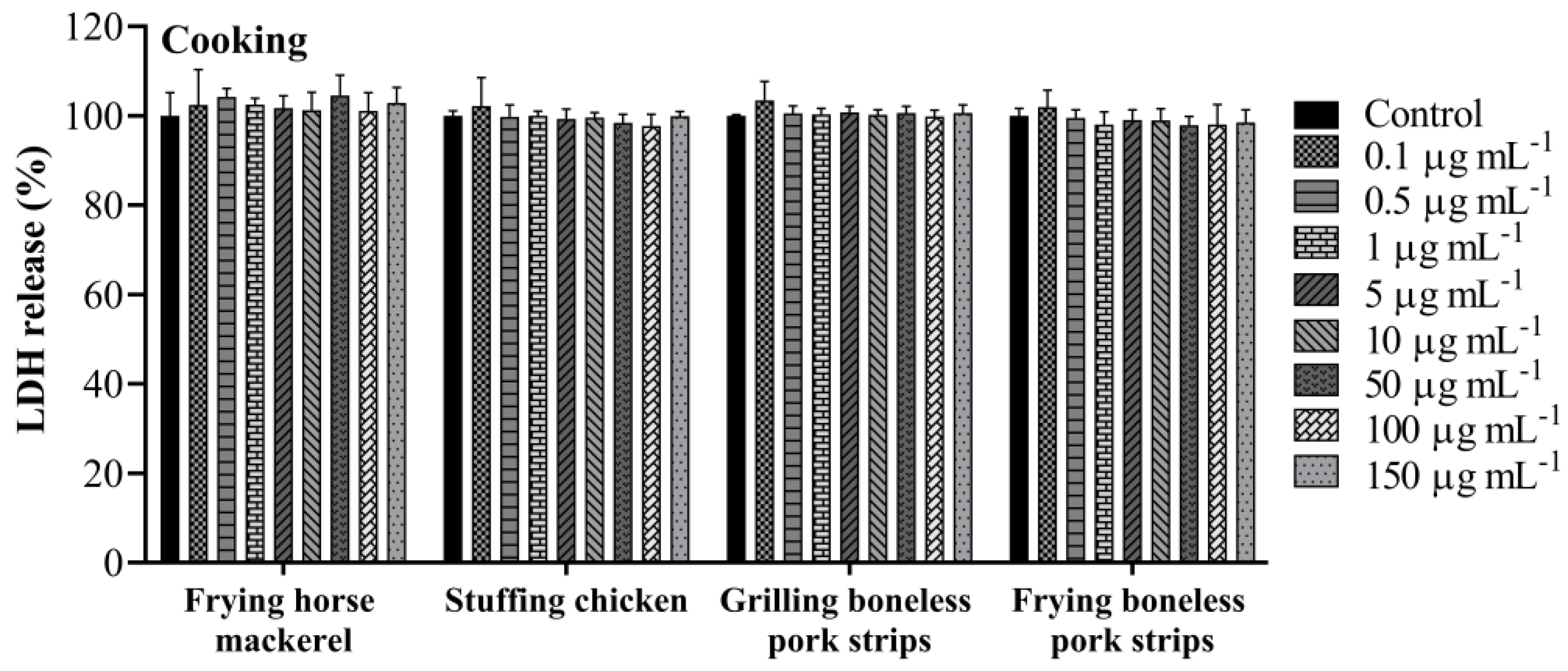
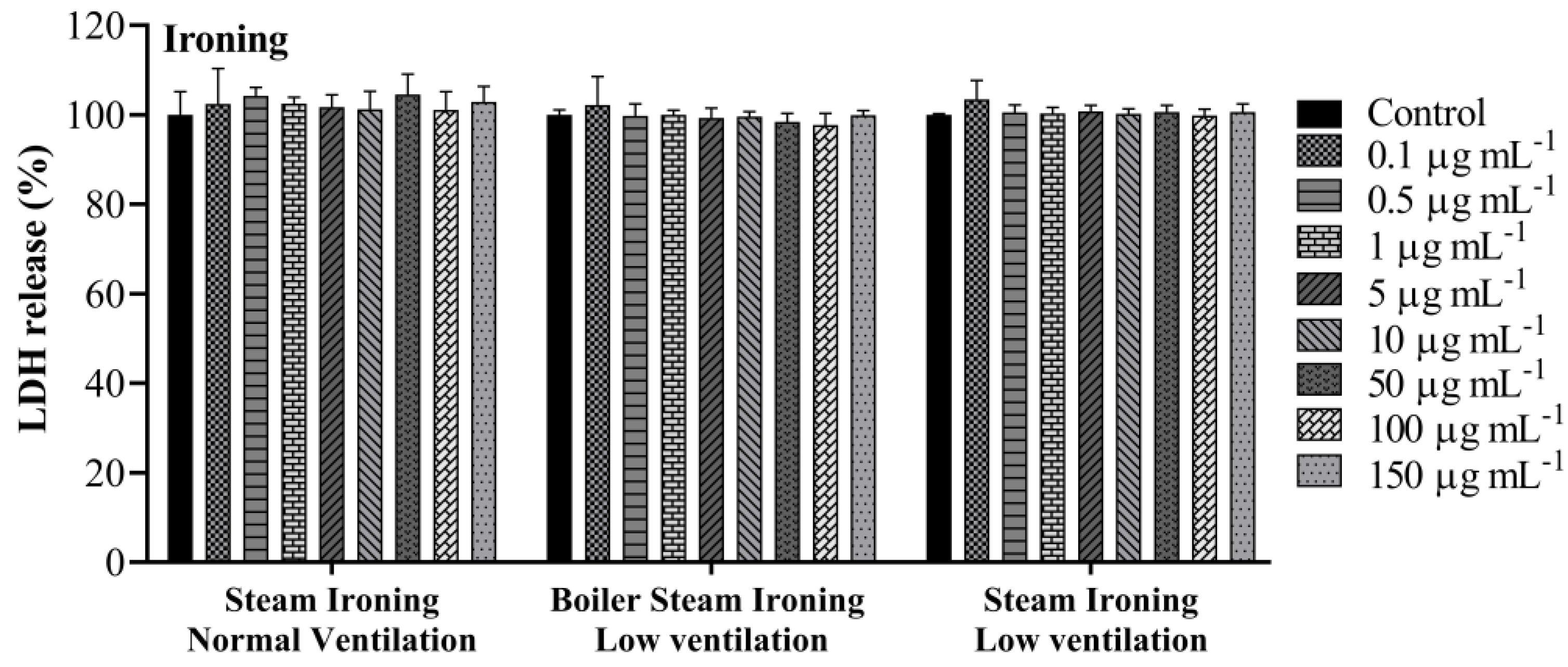
3.3. Lactate Dehydrogenase Activity
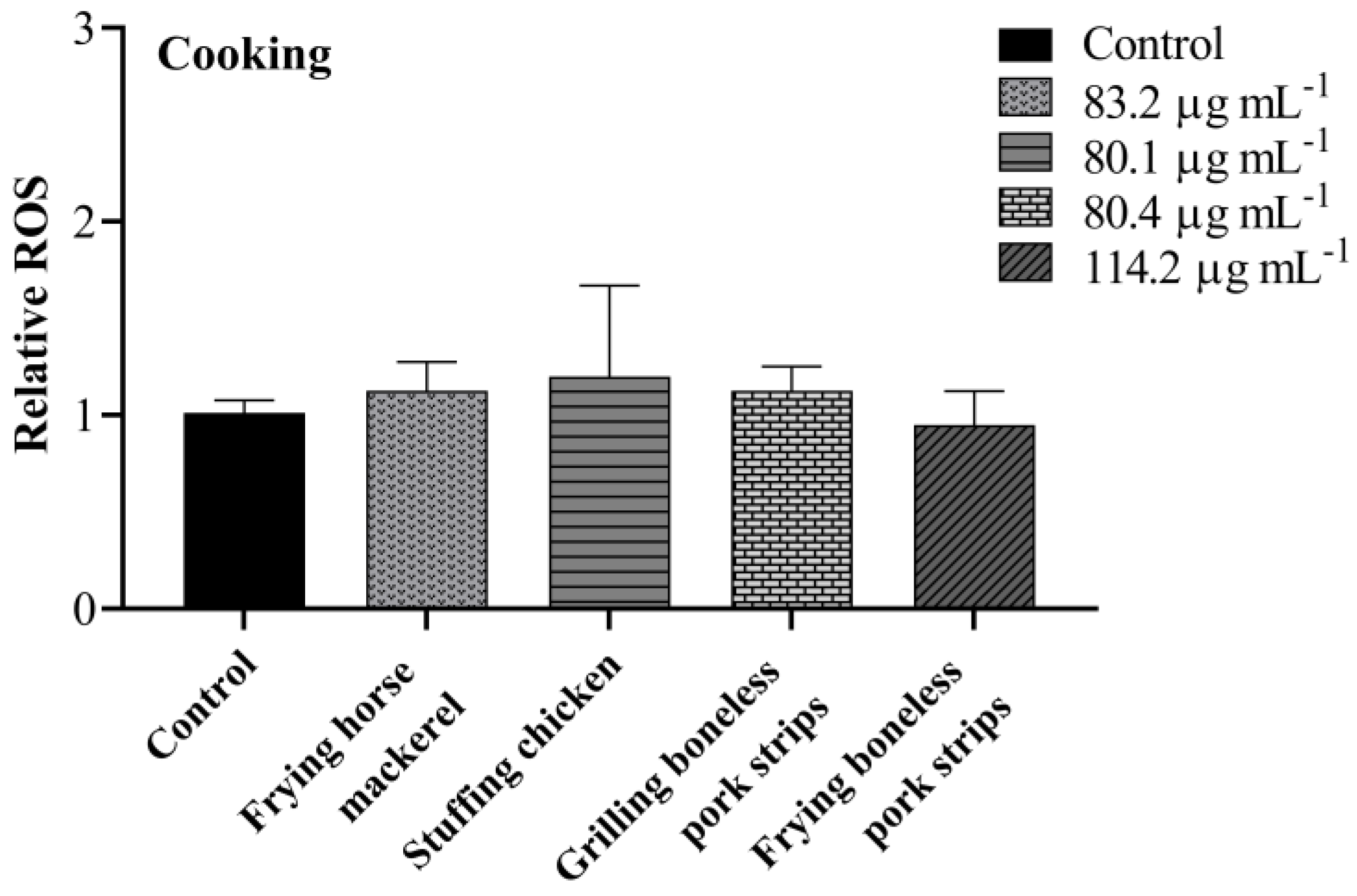
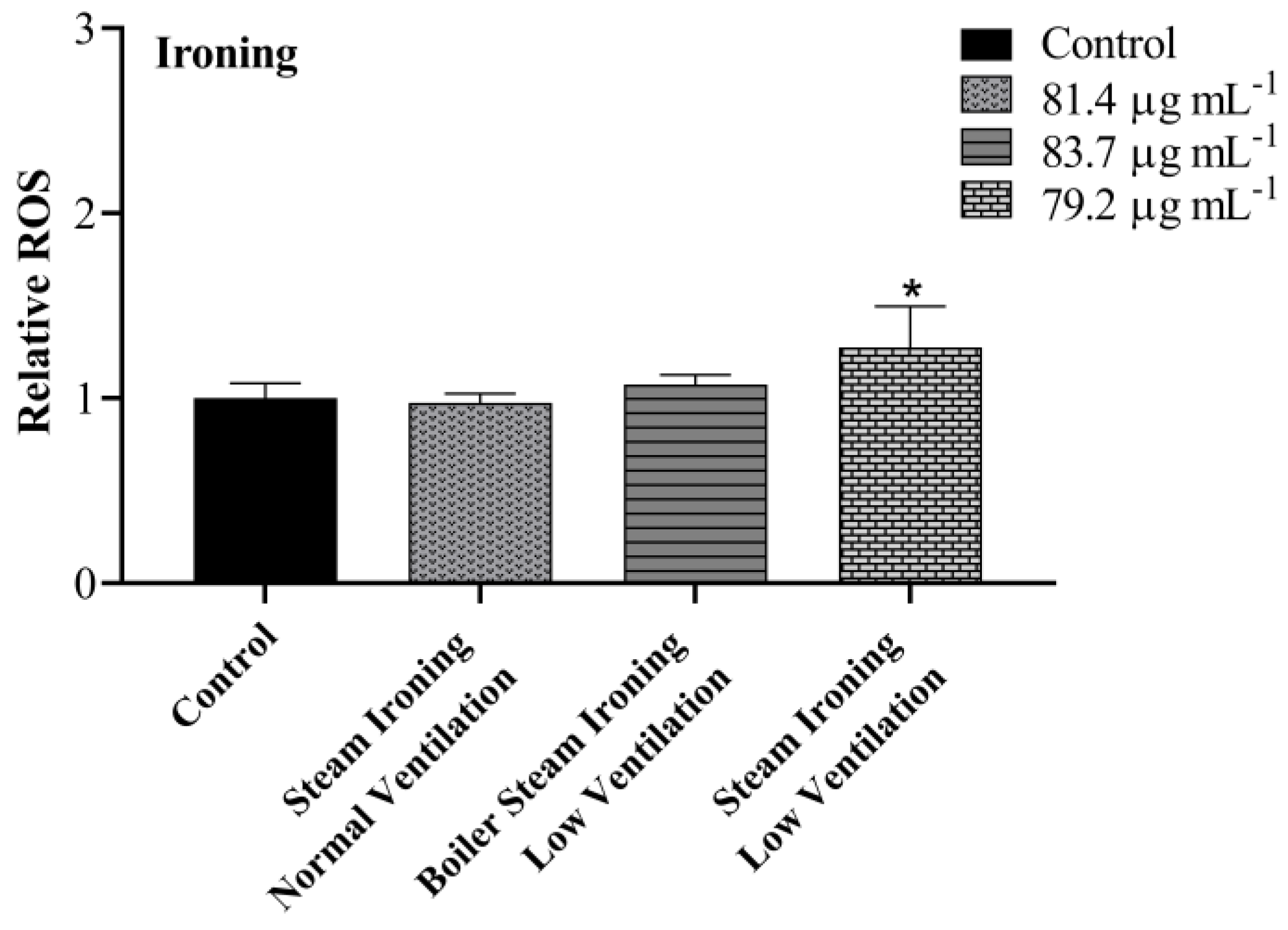
3.4. Intracellular ROS
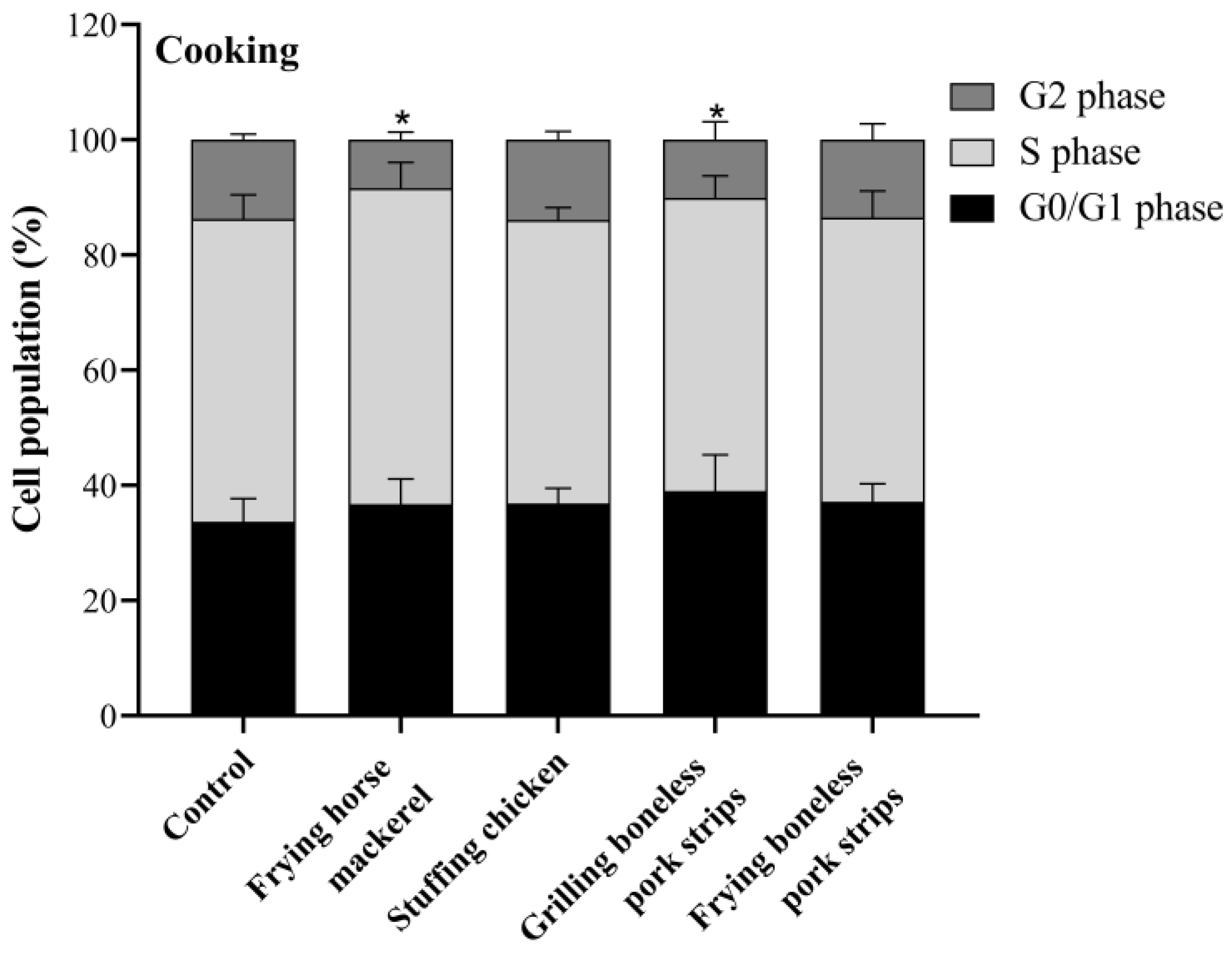
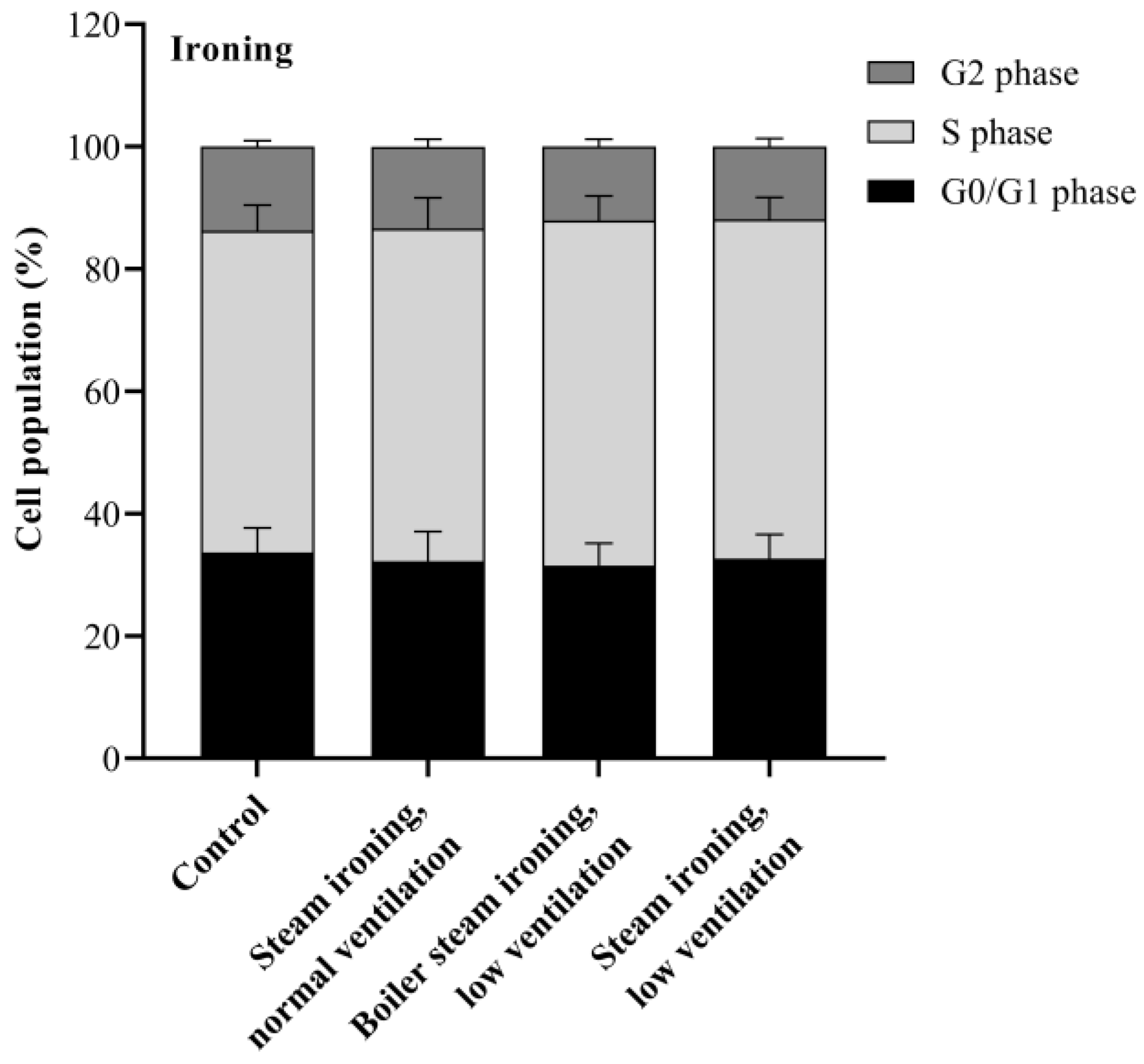
3.5. Cell Cycle
3.6. Mutagenicity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Manisalidis, I.; Stavropoulou, E.; Stavropoulos, A.; Bezirtzoglou, E. Environmental and Health Impacts of Air Pollution: A Review. Front. Public Health 2020, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Abt, E.; Suh, H.H.; Allen, G.; Koutrakis, P. Characterization of indoor particle sources: A study conducted in the metropolitan Boston area. Environ. Health Perspect. 2000, 108, 35–44. [Google Scholar] [CrossRef]
- Zhang, N.; Han, B.; He, F.; Xu, J.; Zhao, R.; Zhang, Y.; Bai, Z. Chemical characteristic of PM2.5 emission and inhalational carcinogenic risk of domestic Chinese cooking. Environ. Pollut. 2017, 227, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Morawska, L.; Afshari, A.; Bae, G.N.; Buonanno, G.; Chao, C.Y.; Hänninen, O.; Hofmann, W.; Isaxon, C.; Jayaratne, E.R.; Pasanen, P.; et al. Indoor aerosols: From personal exposure to risk assessment. Indoor Air 2013, 23, 462–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Kabir, E.; Kabir, S. A review on the human health impact of airborne particulate matter. Environ. Int. 2015, 74, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Gorjinezhad, S.; Kerimray, A.; Torkmahalleh, M.A.; Keleş, M.; Ozturk, F.; Hopke, P.K. Quantifying trace elements in the emitted particulate matter during cooking and health risk assessment. Environ. Sci. Pollut. Res. 2017, 24, 9515–9529. [Google Scholar] [CrossRef]
- Amouei, M.; Gorjinezhad, S.; Sumru, H.; Hopke, P.K. Review of factors impacting emission/concentration of cooking generated particulate matter. Sci. Total Environ. 2017, 586, 1046–1056. [Google Scholar] [CrossRef]
- Abdullahi, K.L.; Delgado-Saborit, J.M.; Harrison, R.M. Emissions and indoor concentrations of particulate matter and its specific chemical components from cooking: A review. Atmos. Environ. 2013, 71, 260–294. [Google Scholar] [CrossRef]
- Schiavon, M.; Rada, E.C.; Ragazzi, M.; Antognoni, S.; Zanoni, S. Domestic activities and PM Generation: A contribution to the understanding of indoor sources of air pollution. Int. J. Sustain. Dev. 2015, 10, 347–360. [Google Scholar] [CrossRef] [Green Version]
- Bhangar, S.; Mullen, N.A.; Hering, S.V.; Kreisberg, N.M.; Nazaroff, W.W. Ultrafine particle concentrations and exposures in seven residences in northern California. Indoor Air 2011, 21, 132–144. [Google Scholar] [CrossRef]
- Afshari, A.; Matson, U.; Ekberg, L.E. Characterization of indoor sources of fine and ultrafine particles: A study conducted in a full-scale chamber. Indoor Air 2005, 15, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Ciuzas, D.; Prasauskas, T.; Krugly, E.; Sidaraviciute, R.; Jurelionis, A.; Seduikyte, L.; Kauneliene, V.; Wierzbicka, A.; Martuzevicius, D. Characterization of indoor aerosol temporal variations for the real-time management of indoor air quality. Atmos. Environ. 2015, 118, 107–117. [Google Scholar] [CrossRef]
- Montoya, L.D.; Hildemann, L.M. Size distributions and height variations of airborne particulate matter and cat allergen indoors immediately following dust-disturbing activities. J. Aerosol. Sci. 2005, 36, 735–749. [Google Scholar] [CrossRef]
- Corsi, R.L.; Siegel, J.A.; Chiang, C. Particle resuspension during the use of vacuum cleaners on residential carpet. J. Occup. Environ. Hyg. 2008, 5, 37–41. [Google Scholar] [CrossRef] [PubMed]
- De Gennaro, G.; Dambruoso, P.R.; Di Gilio, A.; Di Palma, V.; Marzocca, A.; Tutino, M. Discontinuous and continuous indoor air quality monitoring in homes with fireplaces or wood stoves as heating system. Int. J. Environ. Res. Public Health 2015, 13, 78. [Google Scholar] [CrossRef] [Green Version]
- Canha, N.; Lopes, I.; Vicente, E.D.; Vicente, A.M.; Bandowe, B.A.M.; Almeida, S.M.; Alves, C.A. Mutagenicity assessment of aerosols in emissions from domestic combustion processes. Environ. Sci. Pollut. Res. 2016, 23, 10799–10807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicente, E.D.; Figueiredo, D.; Gonçalves, C.; Lopes, I.; Oliveira, H.; Kováts, N.; Pinheiro, T.; Alves, C.A. In vitro toxicity of indoor and outdoor PM10 from residential wood combustion. Sci. Total Environ. 2021, 782, 146820. [Google Scholar] [CrossRef]
- Vicente, E.D.; Figueiredo, D.; Gonçalves, C.; Lopes, I.; Oliveira, H.; Kováts, N.; Pinheiro, T.; Alves, C.A. In vitro toxicity of particulate matter emissions from residential pellet combustion. J. Environ. Sci. 2022, 115, 215–226. [Google Scholar] [CrossRef]
- Massey, D.; Kulshrestha, A.; Masih, J.; Taneja, A. Seasonal trends of PM10, PM5.0, PM2.5 & PM1.0 in indoor and outdoor environments of residential homes located in North-Central India. Build. Environ. 2012, 47, 223–231. [Google Scholar] [CrossRef]
- Wan, M.P.; Wu, C.L.; Sze To, G.N.; Chan, T.C.; Chao, C.Y.H. Ultrafine particles, and PM2.5 generated from cooking in homes. Atmos. Environ. 2011, 45, 6141–6148. [Google Scholar] [CrossRef]
- Katiyar, R.; Rastogi, S.K. Study of indoor air quality of kitchens of rural area’s in Lucknow. Curr. World Environ. 2014, 9, 525–530. [Google Scholar] [CrossRef]
- Chen, Y.; Shen, G.; Huang, Y.; Zhang, Y.; Han, Y.; Wang, R.; Shen, H.; Su, S.; Lin, N.; Zhu, D.; et al. Household air pollution and personal exposure risk of polycyclic aromatic hydrocarbons among rural residents in Shanxi, China. Indoor Air 2016, 26, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Jimenez, J.L.; Canagaratna, M.R.; Allan, J.D.; Coe, H.; Ulbrich, I.; Alfarra, M.R.; Takami, A.; Middlebrook, A.M.; Sun, Y.L.; et al. Ubiquity and dominance of oxygenated species in organic aerosols in anthropogenically-influenced Northern Hemisphere midlatitudes. Geophys. Res. Lett. 2007, 34, L13801. [Google Scholar] [CrossRef] [Green Version]
- Alves, C.A.; Vicente, E.D.; Evtyugina, M.; Vicente, A.M.P.; Sainnokhoi, T.; Kováts, N. Cooking activities in a domestic kitchen: Chemical and toxicological profiling of emissions. Sci. Total Environ. 2021, 772, 145412. [Google Scholar] [CrossRef]
- Kamal, A.; Cincinelli, A.; Martellini, T.; Malik, R.N. A review of PAH exposure from the combustion of biomass fuel and their less surveyed effect on the blood parameters. Environ. Sci. Pollut. Res. 2015, 22, 4076–4098. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, S.; Paul, D.; Hsieh, H.; Yang, H.; Lee, H. Carcinogenic potencies of polycyclic aromatic hydrocarbons for back-door neighbors of restaurants with cooking emissions. Sci. Total Environ. 2012, 417–418, 68–75. [Google Scholar] [CrossRef]
- Dai, W.; Zhong, H.; Li, L.; Cao, J.; Huang, Y.; Shen, M.; Wang, L.; Dong, J.; Tie, X.; Ho, S.S.H.; et al. Characterization and health risk assessment of airborne pollutants in commercial restaurants in northwestern China: Under a low ventilation condition in wintertime. Sci. Total Environ. 2018, 633, 308–316. [Google Scholar] [CrossRef]
- Taner, S.; Pekey, B.; Pekey, H. Fine particulate matter in the indoor air of barbeque restaurants: Elemental compositions, sources and health risks. Sci. Total Environ. 2013, 454–455, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Alves, C.A.; Evtyugina, M.; Cerqueira, M.; Nunes, T.; Duarte, M.; Vicente, E.D. Volatile organic compounds emitted by the stacks of restaurants. Air Qual. Atmos. Health 2015, 8, 401–412. [Google Scholar] [CrossRef]
- Jørgensen, R.B.; Strandberg, B.; Sjaastad, A.K.; Johansen, A.; Svendsen, K. Simulated restaurant cook exposure to emissions of PAHs, mutagenic aldehydes, and particles from frying bacon. J. Occup. Environ Hyg. 2013, 10, 122–131. [Google Scholar] [CrossRef]
- Sjaastad, A.K.; Jørgensen, R.B.; Svendsen, K. Exposure to polycyclic aromatic hydrocarbons (PAHs), mutagenic aldehydes and particulate matter during pan frying of beefsteak. Occup. Environ. Med. 2010, 67, 228–232. [Google Scholar] [CrossRef]
- Zhang, L.; Bai, Z.; You, Y.; Wu, J.; Feng, Y.; Zhu, T. Chemical and stable carbon isotopic characterization for PAHs in aerosol emitted from two indoor sources. Chemosphere 2009, 75, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, Y.Y.; Cao, J.Y.; Tao, F.B.; Zhu, X.X.; Yao, C.J.; Chen, D.J.; Che, Z.; Zhao, Q.H.; Wen, L.P. Oxidative stress, apoptosis, and cell cycle arrest are induced in primary fetal alveolar type II epithelial cells exposed to fine particulate matter from cooking oil fumes. Environ. Sci. Pollut. Res. 2015, 22, 9728–9741. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, T.; Antunes, L.; Bastos, J.; Mayer, A.; Forjaz, G.; Araujo, A.; Nunes, C. Lung cancer: A nationwide study to characterize sex differences, incidence, and spatial patterns in Portugal. In Vivo 2020, 34, 2711–2719. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, T.; Forjaz, G.; Antunes, L.; Bastos, J.; Mayer, A.; Aguiar, P.; Ara, A. Lung cancer survival and sex-specific patterns in Portugal: A population-based analysis. Pulmonology, 2021; in press. [Google Scholar] [CrossRef] [PubMed]
- Osornio-Vargas, A.R.; Bonner, J.C.; Alfaro-Moreno, E.; Martínez, L.; García-Cuellar, C.; Ponce-de-León Rosales, S.; Miranda, J.; Rosas, I. Proinflammatory and Cytotoxic Effects of Mexico City Air Pollution Particulate Matter in Vitro Are Dependent on Particle Size and Composition. Environ. Health Perspect. 2003, 111, 1289–1293. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, H.; Tanaka, K.; Tazuya-murayama, K.; Yamashita, T. Cytotoxic Effects of Water-Soluble Extracts of Coarse and Fine Atmospheric Particulate Matter on Mast Cell Lines. Biol. Pharm. Bull. 2021, 44, 57–62. [Google Scholar] [CrossRef]
- Schins, R.P.F.; Knaapen, A.D.M.; Weishaupt, C.; Winzer, A.; Borm, P.J.A. Cytotoxic and Inflammatory Effects of Coarse and Fine Particulate Matter in Macrophages and Epithelial. Ann. Occup. Hyg. 2002, 46, 203–206. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, I.W.H.; Enlo-Scott, Z.; Nagy, E.; Mudway, I.S.; Tetley, T.D.; Arlt, V.M.; Phillips, D.H. Genotoxicity of fine and coarse fraction ambient particulate matter in immortalised normal (TT1) and cancer-derived (A549) alveolar epithelial cells. Environ. Mol. Mutagen. 2018, 301, 290–301. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.J.; Cao, Z.M.; Guo, X.W.; Chen, H.J.; Lian, Y.; Zheng, W.J.; Chen, Y.J.; Lian, H.Z.; Hu, X. Cytotoxicity and toxicoproteomic analyses of human lung epithelial cells exposed to extracts of atmospheric particulate matters on PTFE filters using acetone and water. Ecotoxicol. Environ. Saf. 2020, 191, 110223. [Google Scholar] [CrossRef]
- Gerlofs-Nijland, M.E.; Dormans, J.A.; Bloemen, H.J.; Leseman, D.L.; John, A.; Boere, F.; Kelly, F.J.; Mudway, I.S.; Jimenez, A.A.; Donaldson, K.; et al. Toxicity of coarse and fine particulate matter from sites with contrasting traffic profiles. Inhal. Toxicol. 2007, 19, 1055–1069. [Google Scholar] [CrossRef] [PubMed]
- Spagnolo, A.M.; Ottria, G.; Perdelli, F.; Cristina, M.L. Chemical Characterisation of the Coarse and Fine Particulate Matter in the Environment of an Underground Railway System: Cytotoxic Effects and Oxidative Stress—A Preliminary Study. Int. J. Environ. Res. Public Health 2015, 12, 4031–4046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, S.; Rivas-Santiago, C.E.; Ibironke, O.A.; Carranza, C.; Meng, Q.; Osornio-Vargas, Á.; Zhang, J.; Torres, M.; Chow, J.C.; Watson, J.G.; et al. Season and size of urban particulate matter differentially affect cytotoxicity and human immune responses to Mycobacterium tuberculosis. PLoS ONE 2019, 14, e0219122. [Google Scholar] [CrossRef] [Green Version]
- Adar, S.D.; Filigrana, P.A.; Clements, N.; Peel, J.L. Ambient Coarse Particulate Matter and Human Health: A Systematic Review and Meta-Analysis. Curr. Environ. Health Rep. 2014, 1, 258–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunekreef, B.; Forsberg, B. Epidemiological evidence of effects of coarse airborne particles on health. Eur. Respir. J. 2005, 26, 309–318. [Google Scholar] [CrossRef]
- Vicente, E.D.; Evtyugina, M.; Vicente, A.M.; Calvo, A.I.; Oduber, F.; Blanco-alegre, C.; Castro, A.; Fraile, R.; Nunes, T.; Lucarelli, F.; et al. Impact of ironing on indoor particle levels and composition. Build. Environ. 2021, 192, 107636. [Google Scholar] [CrossRef]
- Oliveira, H.; Monteiro, C.; Pinho, F.; Pinho, S.; Ferreira de Oliveira, J.M.; Santos, C. Cadmium-induced genotoxicity in human osteoblast-like cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2014, 775–776, 38–47. [Google Scholar] [CrossRef]
- Mortelmans, K.; Zeiger, E. The Ames Salmonella/microsome mutagenicity assay. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2000, 455, 29–60. [Google Scholar] [CrossRef]
- Cao, J.; Ding, R.; Wang, Y.; Chen, D.; Guo, D.; Liang, C. Toxic effect of cooking oil fumes in primary fetal pulmonary type II-like epithelial cells. Environ. Toxicol. Pharmacol. 2013, 36, 320–331. [Google Scholar] [CrossRef]
- Dou, C.; Zhang, J.; Qi, C. Cooking oil fume-derived PM2.5 induces apoptosis in A549 cells and MAPK/NF-кB/STAT1 pathway activation. Environ. Sci. Pollut. Res. 2018, 25, 9940–9948. [Google Scholar] [CrossRef]
- Bandowe, B.A.M.; Lui, K.H.; Jones, T.; BéruBé, K.; Adams, R.; Niu, X.; Wei, C.; Cao, J.J.; Lee, S.C.; Chuang, H.C.; et al. The chemical composition and toxicological effects of fine particulate matter (PM2.5) emitted from different cooking styles. Environ. Pollut. 2021, 288, 117754. [Google Scholar] [CrossRef] [PubMed]
- Maddika, S.; Ande, S.R.; Panigrahi, S.; Paranjothy, T.; Weglarczyk, K.; Zuse, A.; Eshraghi, M.; Manda, K.D.; Wiechec, E.; Los, M. Cell survival, cell death and cell cycle pathways are interconnected: Implications for cancer therapy. Drug Resist. Updates 2007, 10, 13–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oanh, N.T.K.; Nghiem, L.H.; Phyu, Y.L. Emission of Polycyclic Aromatic Hydrocarbons, Toxicity, and Mutagenicity from Domestic Cooking Using Sawdust Briquettes, Wood, and Kerosene. Environ. Sci. Technol. 2002, 36, 833–839. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC20 | |
|---|---|
| Cooking | |
| Frying horse mackerel | 83.2 ± 8.6 |
| stuffing chicken | 80.1 ± 7.0 |
| Grilling boneless pork strips | 80.4 ± 4.0 |
| Frying boneless pork strips | 114.2 ± 3.1 |
| Ironing | |
| Steam ironing normal ventilation | 81.4 ± 5.5 |
| Boiler steam ironing low ventilation | 83.7 ± 2.2 |
| Steam ironing low ventilation | 79.2 ± 5.6 |
| TA100 −S9 | TA100 +S9 | TA98 −S9 | TA98 +S9 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| ng PAHs/Plate | Revertants/Plate | MR | Revertants/Plate | MR | Revertants/Plate | MR | Revertants/Plate | MR | |
| Cooking Fried horse mackerel | 7.5 | 154 ± 30 | 0.7 | 149 ± 10 | 0.89 | 47 ± 33 | 1.4 | 23 ± 3 | 0.99 |
| Stuffed chicken | 4.5 | 153 ± 13 | 0.96 | 142 ± 21 | 0.85 | 43 ± 28 | 1.3 | 22 ± 4 | 0.96 |
| Grilled boneless pork strips | 7.5 | 156 ± 12 | 0.98 | 158 ± 22 | 0.94 | 39 ± 16 | 1.1 | 24 ± 3 | 1.0 |
| Fried boneless pork strips | 5 | 142 ± 19 | 0.89 | 184 ± 10 | 1.1 | 51 ± 16 | 1.5 | 21 ± 4 | 0.90 |
| PC | 3663 ± 741 * | 23 | 606 ± 80 * | 3.6 | 121 ± 16 * | 3.5 | 172 ± 28 * | 7.4 | |
| DMSO | 159 ± 14 | 167 ± 5 | 34 ± 11 | 23 ± 7 | |||||
| Ironing Steam ironing, low ventilation | 11 | 130 ± 3 | 0.83 | 126 ± 8 | 0.85 | 13 ± 4 | 0.74 | 16 ± 3 | 0.66 |
| Steam ironing, normal ventilation | 10 | 142 ± 30 | 0.91 | 137 ± 10 | 0.92 | 14 ± 4 | 0.82 | 24 ± 9 | 1.0 |
| Boiler steam ironing, low ventilation | 29 | 134 ± 22 | 0.86 | 143 ± 9 | 0.96 | 14 ± 1 | 0.80 | 28 ± 2 | 1.2 |
| PC | 2598 ± 329 * | 17 | 489 ± 72 * | 3.3 | 129 ± 9 * | 7.4 | 152 ± 14 * | 6.5 | |
| DMSO | 155 ± 19 | 149 ± 7 | 18 ± 2 | 24 ± 7 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figueiredo, D.; Vicente, E.D.; Vicente, A.; Gonçalves, C.; Lopes, I.; Alves, C.A.; Oliveira, H. Toxicological and Mutagenic Effects of Particulate Matter from Domestic Activities. Toxics 2023, 11, 505. https://doi.org/10.3390/toxics11060505
Figueiredo D, Vicente ED, Vicente A, Gonçalves C, Lopes I, Alves CA, Oliveira H. Toxicological and Mutagenic Effects of Particulate Matter from Domestic Activities. Toxics. 2023; 11(6):505. https://doi.org/10.3390/toxics11060505
Chicago/Turabian StyleFigueiredo, Daniela, Estela D. Vicente, Ana Vicente, Cátia Gonçalves, Isabel Lopes, Célia A. Alves, and Helena Oliveira. 2023. "Toxicological and Mutagenic Effects of Particulate Matter from Domestic Activities" Toxics 11, no. 6: 505. https://doi.org/10.3390/toxics11060505
APA StyleFigueiredo, D., Vicente, E. D., Vicente, A., Gonçalves, C., Lopes, I., Alves, C. A., & Oliveira, H. (2023). Toxicological and Mutagenic Effects of Particulate Matter from Domestic Activities. Toxics, 11(6), 505. https://doi.org/10.3390/toxics11060505