

Investigation of Metal Toxicity on Microalgae Phaeodactylum tricornutum, Hipersaline Zooplankter Artemia salina, and Jellyfish Aurelia aurita

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Toxicity Test with Phaeodactylum tricornutum
2.2. Toxicity Test with Artemia salina
2.3. Toxicity Test with Aurelia aurita
2.4. Statistical Analysis
3. Results
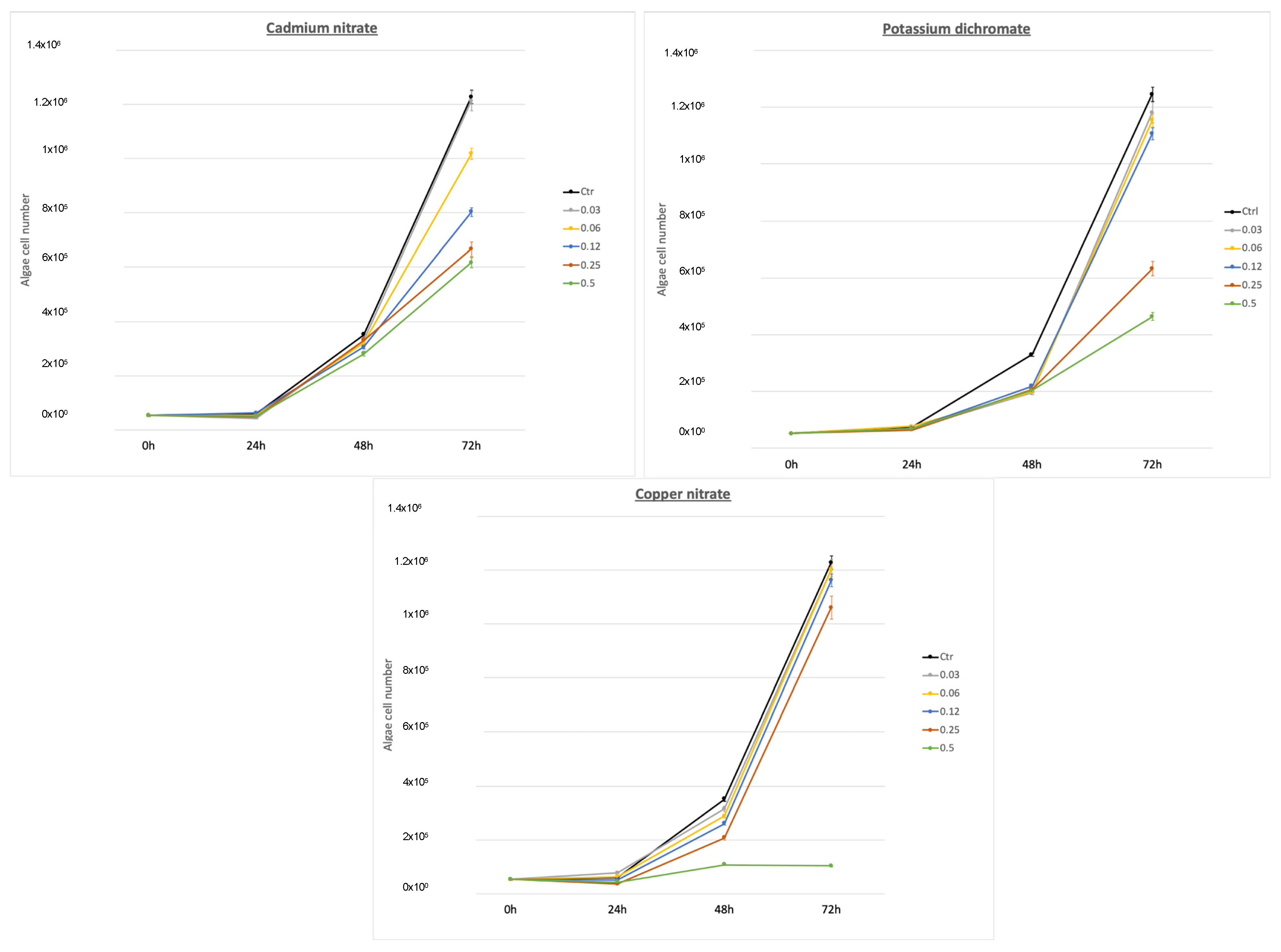
3.1. Phaeodactylum tricornutum (Primary Producer Model)
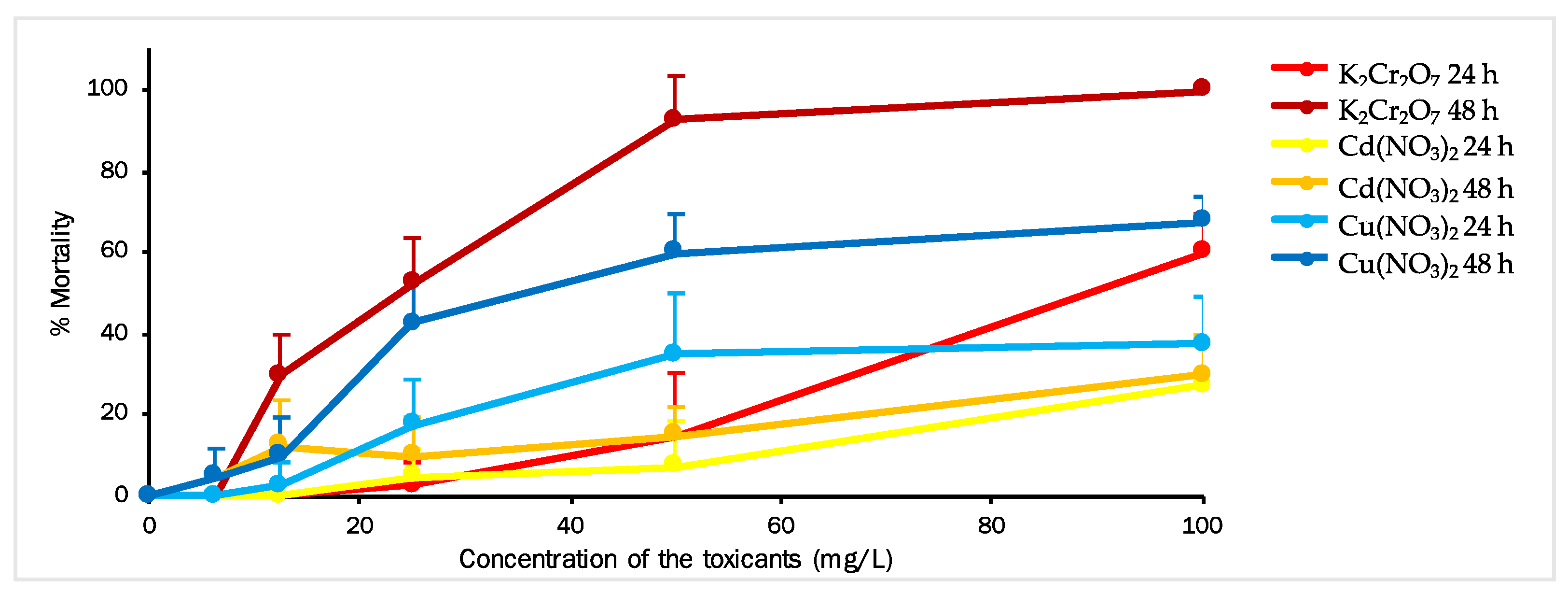
3.2. Artemia salina (Primary Consumer Model)
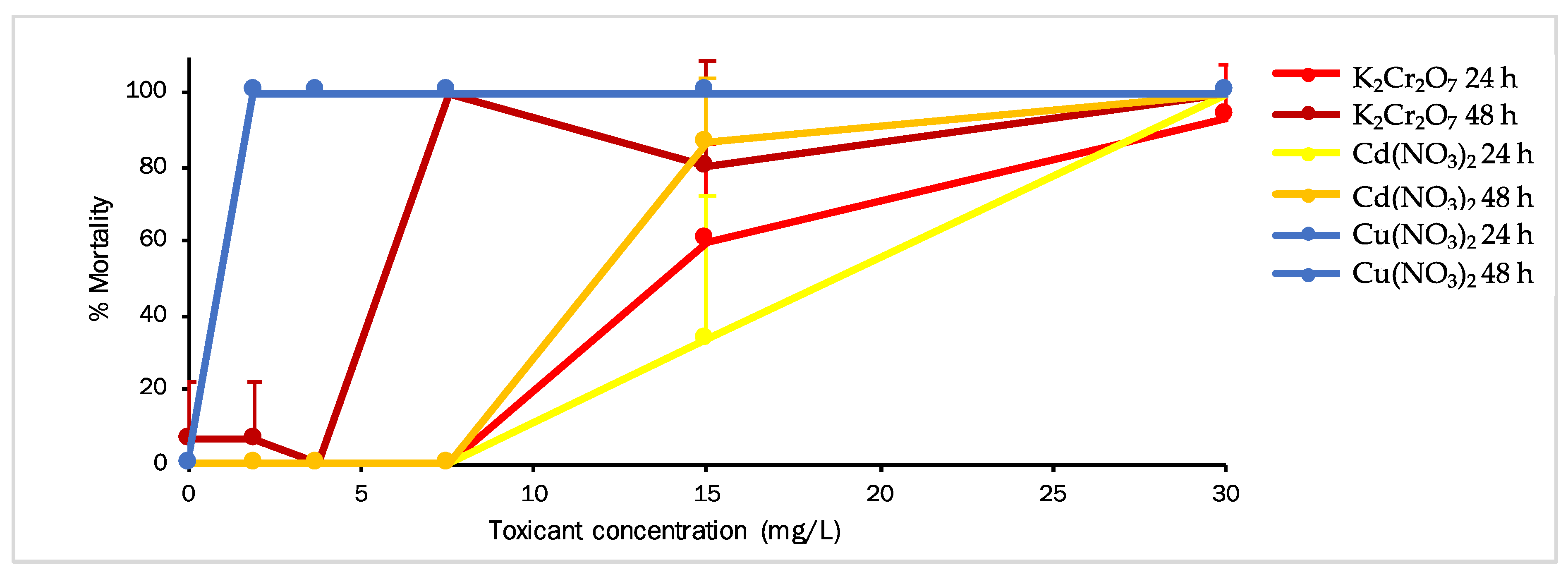
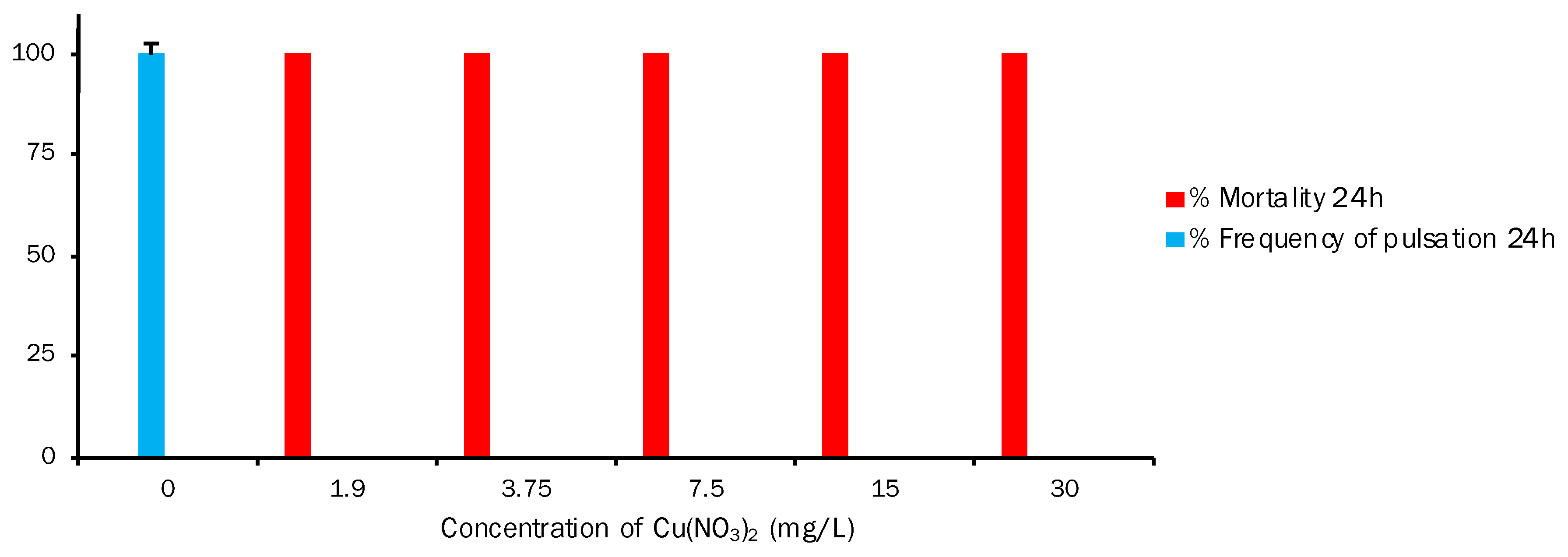
3.3. Aurelia aurita (Secondary Consumer Model)
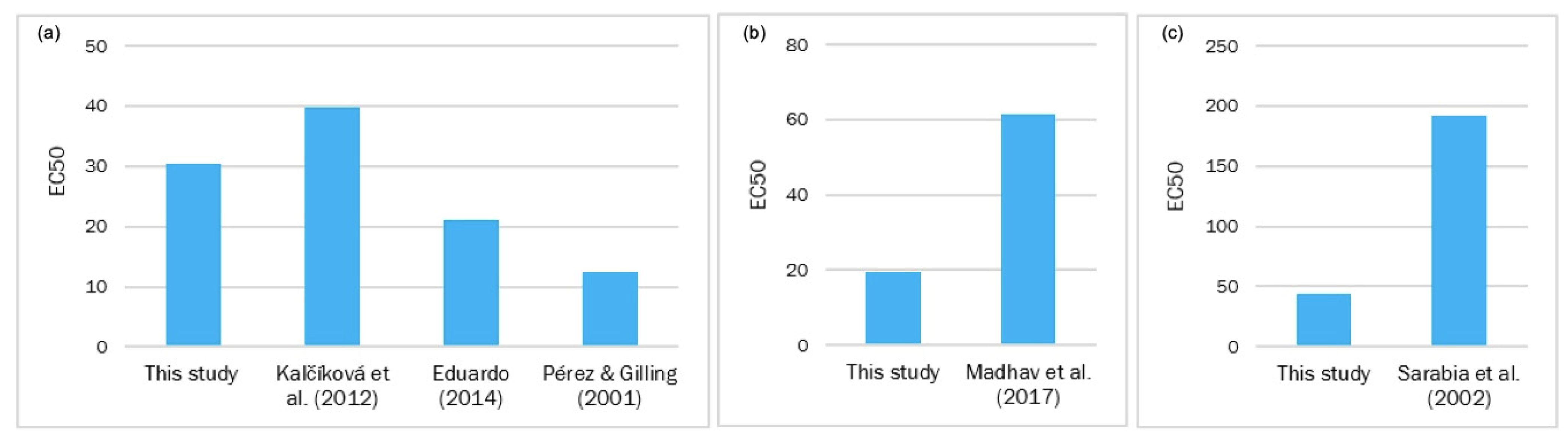
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Z.H.; He, K.; Liu, C.; Li, P.; Zlabek, V. Aquatic Environmental Health and Toxicology. Biomed Res. Int. 2016, 2016, 3514898. [Google Scholar] [CrossRef]
- Abowey, J.F.N.; Sikoki, F.D. Water Pollution Management and Control; Double Trust Publication Company: Port Harcourt, Nigeria, 2007; p. 236. [Google Scholar]
- Ekubo, A.T.; Abowi, J.F.N. Aspects of Aquatic Pollution in Nigeria. Res. J. Environ. Earth Sci. 2011, 6, 673–693. [Google Scholar]
- Hampel, M.; Blasco, J.; Segner, H. Molecular and Cellular Effects of Contamination in Aquatic Ecosystems. Environ. Sci. Pollut. Res. 2015, 22, 17261–17266. [Google Scholar] [CrossRef]
- Baby, J.; Raj, J.; Biby, E.; Sankarganesh, P.; Jeevitha, M.; Ajisha, S.; Rajan, S. Toxic Effect of Heavy Metals on Aquatic Environment. Int. J. Biol. Chem. Sci. 2011, 4, 939–952. [Google Scholar] [CrossRef]
- Farombi, E.; Adelowo, O.; Ajimoko, Y. Biomarkers of Oxidative Stress and Heavy Metal Levels as Indicators of Environmental Pollution in African Cat Fish (Clarias gariepinus) from Nigeria Ogun River. Int. J. Environ. Res. Public Health 2007, 4, 158–165. [Google Scholar] [CrossRef]
- Khayatzadeh, J.; Abbasi, E. The 1 St International Applied Geological Congress; Islamic Azad University: Mashad Branch, Iran, 2010. [Google Scholar]
- Singh, J. Effects of Heavy Metals on Soil, Plants, Human Health and Aquatic Life. Int. J. Res. Chem. Environ. 2011, 1, 15–21. [Google Scholar]
- Alloway, B.J. (Ed.) Heavy Metals in Soils; Environmental Pollution; Springer: Dordrecht, The Netherlands, 2013; Volume 22, ISBN 978-94-007-4469-1. [Google Scholar]
- Launay, M.A.; Dittmer, U.; Steinmetz, H. Organic Micropollutants Discharged by Combined Sewer Overflows—Characterisation of Pollutant Sources and Stormwater-Related Processes. Water Res. 2016, 104, 82–92. [Google Scholar] [CrossRef]
- Fifield, F.W.; Haines, P.J. Environmental Analytical Chemistry, 2nd ed.; Blackwell Science: London, UK, 2000. [Google Scholar]
- Loska, K.; Wiechuła, D. Application of Principal Component Analysis for the Estimation of Source of Heavy Metal Contamination in Surface Sediments from the Rybnik Reservoir. Chemosphere 2003, 51, 723–733. [Google Scholar] [CrossRef]
- Harguinteguy, C.A.; Cirelli, A.F.; Pignata, M.L. Heavy Metal Accumulation in Leaves of Aquatic Plant Stuckenia Filiformis and Its Relationship with Sediment and Water in the Suquía River (Argentina). Microchem. J. 2014, 114, 111–118. [Google Scholar] [CrossRef]
- Bashir, I.; Lone, F.A.; Bhat, R.A.; Mir, S.A.; Dar, Z.A.; Dar, S.A. Concerns and Threats of Contamination on Aquatic Ecosystems. In Bioremediation and Biotechnology; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–26. [Google Scholar]
- Zhou, Q.; Zhang, J.; Fu, J.; Shi, J.; Jiang, G. Biomonitoring: An Appealing Tool for Assessment of Metal Pollution in the Aquatic Ecosystem. Anal. Chim. Acta 2008, 606, 135–150. [Google Scholar] [CrossRef]
- Phillips, D.J.H. The Use of Biological Indicator Organisms to Monitor Trace Metal Pollution in Marine and Estuarine Environments—A Review; Applied Science Publishers Ltd.: London, UK, 1977. [Google Scholar]
- Calzoni, G.L.; Antognoni, F.; Pari, E.; Fonti, P.; Gnes, A.; Speranza, A. Active Biomonitoring of Heavy Metal Pollution Using Rosa Rugosa Plants. Environ. Pollut. 2007, 149, 239–245. [Google Scholar] [CrossRef]
- Grant, A.; Trompf, K.; Seung, D.; Nivison-Smith, L.; Bowcock, H.; Kresse, H.; Morrow, P. Sub-cellular damage by copper in the cnidarian Zoanthus robustus. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2010, 152, 256–262. [Google Scholar] [CrossRef]
- Burger, J. Bioindicators: A Review of Their Use in the Environmental Literature 1970–2005. Environ. Bioindic. 2006, 1, 136–144. [Google Scholar] [CrossRef]
- Nordberg, G.F. Historical Perspectives on Cadmium Toxicology. Toxicol. Appl. Pharmacol. 2009, 238, 192–200. [Google Scholar] [CrossRef]
- Pavlaki, M.D.; Araújo, M.J.; Cardoso, D.N.; Silva, A.R.R.; Cruz, A.; Mendo, S.; Soares, A.M.V.M.; Calado, R.; Loureiro, S. Ecotoxicity and Genotoxicity of Cadmium in Different Marine Trophic Levels. Environ. Pollut. 2016, 215, 203–212. [Google Scholar] [CrossRef]
- Chandurvelan, R.; Marsden, I.D.; Gaw, S.; Glover, C.N. Waterborne Cadmium Impacts Immunocytotoxic and Cytogenotoxic Endpoints in Green-Lipped Mussel, Perna Canaliculus. Aquat. Toxicol. 2013, 142–143, 283–293. [Google Scholar] [CrossRef]
- Engel, D.W.; Fowler, B.A. Factors Influencing Cadmium Accumulation and Its Toxicity to Marine Organisms. Environ. Health Perspect. 1979, 28, 81–88. [Google Scholar] [CrossRef]
- Hadjispyrou, S.; Kungolos, A.; Anagnostopoulos, A. Toxicity, Bioaccumulation, and Interactive Effects of Organotin, Cadmium, and Chromium on Artemia Franciscana. Ecotoxicol. Environ. Saf. 2001, 49, 179–186. [Google Scholar] [CrossRef]
- Yuleidis González Pérez, L.; Patricia, L.; Gilling, A. Determinación de la Toxicidad Aguda del Dicromato de Potasio en Larvas de Artemia salina; Centro de Toxicología y Biomedicina/Cuba: Santiago de Cuba, Cuba, 2001; Volume 1. [Google Scholar]
- Lopez, J.S.; Lee, L.; Mackey, K.R.M. The Toxicity of Copper to Crocosphaera Watsonii and Other Marine Phytoplankton: A Systematic Review. Front. Mar. Sci. 2019, 5, 00511. [Google Scholar] [CrossRef]
- Nriagu, J.O. Global Inventory of Natural and Anthropogenic Emissions of Trace Metals to the Atmosphere. Nature 1979, 279, 409–411. [Google Scholar] [CrossRef]
- Duce, R.A.; Liss, P.S.; Merrill, J.T.; Atlas, E.L.; Buat-Menard, P.; Hicks, B.B.; Miller, J.M.; Prospero, J.M.; Arimoto, R.; Church, T.M.; et al. The Atmospheric Input of Trace Species to the World Ocean. Glob. Biogeochem. Cycles 1991, 5, 193–259. [Google Scholar] [CrossRef]
- Prospero, J.M. Long-Term Measurements of the Transport of African Mineral Dust to the Southeastern United States: Implications for Regional Air Quality. J. Geophys. Res. Atmos. 1999, 104, 15917–15927. [Google Scholar] [CrossRef]
- Maenhaut, W.; Salma, I.; Cafmeyer, J.; Annegarn, H.J.; Andreae, M.O. Regional Atmospheric Aerosol Composition and Sources in the Eastern Transvaal, South Africa, and Impact of Biomass Burning. J. Geophys. Res. 1996, 101, 23631–23650. [Google Scholar] [CrossRef]
- Casado-Martínez, M. Interlaboratory Assessment of Marine Bioassays to Evaluate the Environmental Quality of Coastal Sediments in Spain. III. Bioassay using embryos sea urchin Paracentrotus lividus. Cienc. Mar. 2006, 32, 139–147. [Google Scholar] [CrossRef]
- Gambardella, C.; Costa, E.; Piazza, V.; Fabbrocini, A.; Magi, E.; Faimali, M.; Garaventa, F. Effect of Silver Nanoparticles on Marine Organisms Belonging to Different Trophic Levels. Mar. Environ. Res. 2015, 111, 41–49. [Google Scholar] [CrossRef]
- Topçu, N.E.; Martell, L.F.; Yilmaz, I.N.; Isinibilir, M. Benthic Hydrozoans as Potential Indicators of Water Masses and Anthropogenic Impact in the Sea of Marmara. Mediterr. Mar. Sci. 2018, 273, 15117. [Google Scholar] [CrossRef]
- Daly, M.; Brugler, M.R.; Cartwright, P.; Collins, A.G.; Dawson, M.N.; Fautin, D.G.; France, S.C.; Mcfadden, C.S.; Opresko, D.M.; Rodriguez, E.; et al. The Phylum Cnidaria: A Review of Phylogenetic Patterns and Diversity 300 Years after Linnaeus*. Zootaxa 2007, 1668, 127–182. [Google Scholar] [CrossRef]
- Boero, F.; Bouillon, J.; Gravili, C.; Miglietta, M.; Parsons, T.; Piraino, S. Gelatinous Plankton: Irregularities Rule the World (Sometimes). Mar. Ecol. Prog. Ser. 2008, 356, 299–310. [Google Scholar] [CrossRef]
- Epstein, H.E.; Templeman, M.A.; Kingsford, M.J. Fine-Scale Detection of Pollutants by a Benthic Marine Jellyfish. Mar. Pollut. Bull. 2016, 107, 340–346. [Google Scholar] [CrossRef]
- Richardson, A.J.; Bakun, A.; Hays, G.C.; Gibbons, M.J. The Jellyfish Joyride: Causes, Consequences and Management Responses to a More Gelatinous Future. Trends Ecol. Evol. 2009, 24, 312–322. [Google Scholar] [CrossRef]
- Sweetman, A.K.; Smith, C.R.; Dale, T.; Jones, D.O.B. Rapid Scavenging of Jellyfish Carcasses Reveals the Importance of Gelatinous Material to Deep-Sea Food Webs. Proc. R. Soc. B Biol. Sci. 2014, 281, 20142210. [Google Scholar] [CrossRef]
- Hays, G.C.; Doyle, T.K.; Houghton, J.D.R. A Paradigm Shift in the Trophic Importance of Jellyfish? Trends Ecol. Evol. 2018, 33, 874–884. [Google Scholar] [CrossRef]
- Sullivan, B.K.; Garcia, J.R.; Klein-Macphee, G. Prey Selection by the Scyphomedusan Predator Aurelia Aurita; Springer-Verlag: Berlin/Heidelberg, Germany, 1994; Volume 121. [Google Scholar]
- Purcell, J. Predation on Zooplankton by Large Jellyfish, Aurelia, Cyanea and Aequorea, in Prince William Sound, Alaska. Mar. Ecol. Prog. Ser. 2003, 246, 137–152. [Google Scholar] [CrossRef]
- Colin, S.P.; Costello, J.H.; Graham, W.M.; Higgins, J.I. Omnivory by the Small Cosmopolitan Hydromedusa Aglaura Hemistoma. Limnol. Oceanogr. 2005, 50, 1264–1268. [Google Scholar] [CrossRef]
- Pitt, K.A.; Clement, A.-L.; Connolly, R.M.; Thibault-Botha, D. Predation by Jellyfish on Large and Emergent Zooplankton: Implications for Benthic–Pelagic Coupling. Estuar. Coast. Shelf Sci. 2008, 76, 827–833. [Google Scholar] [CrossRef]
- Macali, A.; Bergami, E. Jellyfish as Innovative Bioindicator for Plastic Pollution. Ecol. Indic. 2020, 115, 106375. [Google Scholar] [CrossRef]
- Costa, E.; Gambardella, C.; Piazza, V.; Vassalli, M.; Sbrana, F.; Lavorano, S.; Garaventa, F.; Faimali, M. Microplastics Ingestion in the Ephyra Stage of Aurelia Sp. triggers acute and behavioral responses. Ecotoxicol. Environ. Saf. 2020, 189, 109983. [Google Scholar] [CrossRef]
- Almeda, R.; Wambaugh, Z.; Chai, C.; Wang, Z.; Liu, Z.; Buskey, E.J. Effects of Crude Oil Exposure on Bioaccumulation of Polycyclic Aromatic Hydrocarbons and Survival of Adult and Larval Stages of Gelatinous Zooplankton. PLoS ONE 2013, 8, e74476. [Google Scholar] [CrossRef]
- Echols, B.S.; Smith, A.J.; Gardinali, P.R.; Rand, G.M. The Use of Ephyrae of a Scyphozoan Jellyfish, Aurelia Aurita, in the Aquatic Toxicological Assessment of Macondo Oils from the Deepwater Horizon Incident. Chemosphere 2016, 144, 1893–1900. [Google Scholar] [CrossRef]
- Faimali, M.; Garaventa, F.; Piazza, V.; Costa, E.; Greco, G.; Mazzola, V.; Beltrandi, M.; Bongiovanni, E.; Lavorano, S.; Gnone, G. Ephyra Jellyfish as a New Model for Ecotoxicological Bioassays. Mar. Environ. Res. 2014, 93, 93–101. [Google Scholar] [CrossRef]
- Lechable, M.; Jan, A.; Duchene, A.; Uveira, J.; Weissbourd, B.; Gissat, L.; Collet, S.; Gilletta, L.; Chevalier, S.; Leclère, L.; et al. An Improved Whole Life Cycle Culture Protocol for the Hydrozoan Genetic Model Clytia Hemisphaerica. Biol. Open 2020, 9, bio051268. [Google Scholar] [CrossRef] [PubMed]
- Lucas, C.H. Reproduction and Life History Strategies of the Common Jellyfish, Aurelia Aurita. Hydrobiologia 2001, 451, 229–246. [Google Scholar] [CrossRef]
- van der Veer, H.W.; Oorthuysen, W. Abundance, Growth and Food Demand of the Scyphomedusa Aurelia Aurita in the Western Wadden Sea. Neth. J. Sea Res. 1985, 19, 38–44. [Google Scholar] [CrossRef]
- Carré, D.; Carré, C. Origin of Germ Cells, Sex Determination, and Sex Inversion in Medusae of the Genus Clytia (Hydrozoa, Leptomedusae): The Influence of Temperature. J. Exp. Zool. 2000, 287, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Larsen, G.D. Unraveling the Mysteries of the Medusa. Lab. Anim. 2016, 45, 163. [Google Scholar] [CrossRef]
- Houliston, E.; Momose, T.; Manuel, M. Clytia Hemisphaerica: A Jellyfish Cousin Joins the Laboratory. Trends Genet. 2010, 26, 159–167. [Google Scholar] [CrossRef]
- Bae, M.-J.; Park, Y.-S. Biological Early Warning System Based on the Responses of Aquatic Organisms to Disturbances: A Review. Sci. Total Environ. 2014, 466–467, 635–649. [Google Scholar] [CrossRef]
- Uba, B.O. Effects of Aromatic Hydrocarbons and Marine Sediments from Niger Delta on the Growth of Microalga Phaeodactylum tricornutum. Biotechnol. J. Int. 2019, 22, 1–18. [Google Scholar] [CrossRef]
- Pastorino, P.; Broccoli, A.; Anselmi, S.; Bagolin, E.; Prearo, M.; Barceló, D.; Renzi, M. The Microalgae Chaetoceros tenuissimus Exposed to Contaminants of Emerging Concern: A Potential Alternative to Standardized Species for Marine Quality Assessment. Ecol. Indic. 2022, 141, 109075. [Google Scholar] [CrossRef]
- Piccardo, M.; Provenza, F.; Grazioli, E.; Anselmi, S.; Terlizzi, A.; Renzi, M. Impacts of Plastic-Made Packaging on Marine Key Species: Effects Following Water Acidification and Ecological Implications. J. Mar. Sci. Eng. 2021, 9, 432. [Google Scholar] [CrossRef]
- Clément, L.; Hurel, C.; Marmier, N. Toxicity of TiO2 Nanoparticles to Cladocerans, Algae, Rotifers and Plants—Effects of Size and Crystalline Structure. Chemosphere 2013, 90, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Si, D.; Yang, L.; Yan, H.; Wang, Q. Bioaccumulation and Transformation of Cadmium by Phaeodactylum tricornutum. Sci. China B Chem. 2009, 52, 2373–2380. [Google Scholar] [CrossRef]
- Okamoto, O.K.; Asano, C.S.; Aidar, E.; Colepicolo, P. Effects of Cadmium on Growth and Superoxide Dismutase Activity of the Marine Migroalga Tetraselmis gracilis. J. Phycol. 1996, 32, 74–79. [Google Scholar] [CrossRef]
- Wang, L.; Zheng, B. Toxic Effects of Fluoranthene and Copper on Marine Diatom Phaeodactylum tricornutum. J. Environ. Sci. 2008, 20, 1363–1372. [Google Scholar] [CrossRef]
- Jung, S.M.; Bae, J.S.; Kang, S.G.; Son, J.S.; Jeon, J.H.; Lee, H.J.; Jeon, J.Y.; Sidharthan, M.; Ryu, S.H.; Shin, H.W. Acute Toxicity of Organic Antifouling Biocides to Phytoplankton Nitzschia Pungens and Zooplankton Artemia Larvae. Mar. Pollut. Bull. 2017, 124, 811–818. [Google Scholar] [CrossRef]
- Franklin, N.M.; Stauber, J.L.; Apte, S.C.; Lim, R.P. Effect of Initial Cell Density on the Bioavailability and Toxicity of Copper in Microalgal Bioassays. Environ. Toxicol. Chem. 2002, 21, 742–751. [Google Scholar] [CrossRef]
- Rico, M.; López, A.; Santana-Casiano, J.M.; Gonzàlez, A.G.; Gonzàlez-Dàvila, M. Variability of the Phenolic Profile in the Diatom Phaeodactylum tricornutum Growing under Copper and Iron Stress. Limnol. Oceanogr. 2013, 58, 144–152. [Google Scholar] [CrossRef]
- Zulkifli, S.Z.; Aziz, F.Z.A.; Ajis, S.Z.M.; Ismail, A. Nauplii of Brine Shrimp (Artemia salina) as a Potential Toxicity Testing Organism for Heavy Metals Contamination. In From Sources to Solution; Springer: Singapore, 2014; pp. 233–237. [Google Scholar]
- Fichet, D.; Miramand, P. Vanadium Toxicity to Three Marine Invertebrates Larvae: Crassostrea Gigas, Paracentrotus Lividus and Artemia Salina. Chemosphere 1998, 37, 1363–1368. [Google Scholar] [CrossRef]
- Laughlin, R.B.; Ng, J.; Guard, H.E. Hormesis: A Response to Low Environmental Concentrations of Petroleum Hydrocarbons. Science (1979) 1981, 211, 705–707. [Google Scholar] [CrossRef]
- Karbassi, A.; Bidhendi, G.N.; Pejman, A.; Bidhendi, M.E. Environmental Impacts of Desalination on the Ecology of Lake Urmia. J. Great Lakes Res. 2010, 36, 419–424. [Google Scholar] [CrossRef]
- Sarabia, R.; Del, R.J.; Varo, I.; Díaz-Mayans, J.; Torreblanca, A. Comparing the Acute Response to Cadmium Toxicity of Nauplii from Different Populations of Artemia. Environ. Toxicol. Chem. 2002, 21, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Kalčíková, G.; Zagorc-Končan, J.; Žgajnar Gotvajn, A. Artemia salina acute immobilization test: A possible tool for aquatic ecotoxicity assessment. Water Sci. Technol. 2012, 66, 903–908. [Google Scholar] [CrossRef]
- Umarani, R.; Kumaraguru, A.K.; Nagarani, N. Investigation of Acute Toxicity of Heavy Metals in Artemia Salina Acclimated to Different Salinity. Toxicol. Environ. Chem. 2012, 94, 1547–1556. [Google Scholar] [CrossRef]
- Eduardo Ruiz González, L.; Vega-Villasante, F. Evaluación de La Toxicidad de Algunos Basidiomycetes Del Estado de Jalisco Sobre Artemia franciscana. Tesis Maest. Cienc. 2014, 34, 220–224. [Google Scholar] [CrossRef]
- Eisler, R. Cadmium Poisoning in Fundulus Heteroclitus (Pisces: Cyprinodontidae) and Other Marine Organisms. J. Fish. Res. Board. Can. 1971, 28, 1225–1234. [Google Scholar] [CrossRef]
- Madhav, M.R.; David, S.E.M.; Kumar, R.S.S.; Swathy, J.S.; Bhuvaneshwari, M.; Mukherjee, A.; Chandrasekaran, N. Toxicity and Accumulation of Copper Oxide (CuO) Nanoparticles in Different Life Stages of Artemia Salina. Environ. Toxicol. Pharmacol. 2017, 52, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Lucas, C.H.; Horton, A.A. Short-Term Effects of the Heavy Metals, Silver and Copper, on Polyps of the Common Jellyfish, Aurelia Aurita. J. Exp. Mar. Biol. Ecol. 2014, 461, 154–161. [Google Scholar] [CrossRef]
- Karntanut, W.; Pascoe, D. The Toxicity of Copper, Cadmium and Zinc to Four Different Hydra (Cnidaria: Hydrozoa). Chemosphere 2002, 47, 1059–1064. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Salt Concentration 1 | Cu | Cr | Cd |
|---|---|---|---|---|
| P. tricornutum | 0.03 | 0.062 | 0.109 | 0.001 |
| 0.06 | 0.068 | 0.119 | 0.009 | |
| 0.12 | 0.08 | 0.139 | 0.027 | |
| 0.25 | 0.107 | 0.182 | 0.065 | |
| A. salina | 6.25 | 1.322 | 2.174 | 1.81 |
| 12.5 | 2.587 | 4.249 | 3.627 | |
| 25 | 5.119 | 8.399 | 7.262 | |
| 50 | 10.181 | 16.699 | 14.532 | |
| 100 | 20.306 | 33.299 | 29.072 | |
| A. aurita | 1.9 | 0.441 | 0.73 | 0.545 |
| 3.75 | 0.813 | 1.344 | 1.083 | |
| 7.5 | 1.575 | 2.589 | 2.173 | |
| 2.173 | 3.094 | 5.079 | 4.354 | |
| 30 | 6.131 | 10.059 | 8.716 |
| Organism | Toxicant | Exposure Time (h) | EC50 (mg/L) (Mean ± SD) |
|---|---|---|---|
| Phaeodactylum tricornutum | K2Cr2O7 | 72 | 15.378 ± 7.081 |
| Cd(NO3)2 | 72 | 2.494 ± 2.494 | |
| Cu(NO3)2 | 72 | 1.205 ± 0.322 | |
| Artemia salina | K2Cr2O7 | 24 | 91.359 ± 6.746 |
| 48 | 23.554 ± 2.383 | ||
| Cd(NO3)2 | 24 | 150.167 ± 27.496 | |
| 48 | 153.840 ± 65.674 | ||
| Cu(NO3)2 | 24 | 95.773 ± 28.284 | |
| 48 | 37.201 ± 1.872 | ||
| Aurelia aurita | K2Cr2O7 | 24 | 16.571 ± 4.246 |
| 48 | 6.726 ± 2.004 | ||
| Cd(NO3)2 | 24 | 19.880 ± 5.519 | |
| 48 | 12.343 ± 2.588 | ||
| Cu(NO3)2 | 24 | 0.283 ± 0 | |
| 48 | 0.283 ± 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mercado, B.; Valero, N.; Roca-Pérez, L.; Bernabeu-Berni, E.; Andreu-Sánchez, O. Investigation of Metal Toxicity on Microalgae Phaeodactylum tricornutum, Hipersaline Zooplankter Artemia salina, and Jellyfish Aurelia aurita. Toxics 2023, 11, 716. https://doi.org/10.3390/toxics11080716
Mercado B, Valero N, Roca-Pérez L, Bernabeu-Berni E, Andreu-Sánchez O. Investigation of Metal Toxicity on Microalgae Phaeodactylum tricornutum, Hipersaline Zooplankter Artemia salina, and Jellyfish Aurelia aurita. Toxics. 2023; 11(8):716. https://doi.org/10.3390/toxics11080716
Chicago/Turabian StyleMercado, Borja, Nuria Valero, Luis Roca-Pérez, Elena Bernabeu-Berni, and Oscar Andreu-Sánchez. 2023. "Investigation of Metal Toxicity on Microalgae Phaeodactylum tricornutum, Hipersaline Zooplankter Artemia salina, and Jellyfish Aurelia aurita" Toxics 11, no. 8: 716. https://doi.org/10.3390/toxics11080716
APA StyleMercado, B., Valero, N., Roca-Pérez, L., Bernabeu-Berni, E., & Andreu-Sánchez, O. (2023). Investigation of Metal Toxicity on Microalgae Phaeodactylum tricornutum, Hipersaline Zooplankter Artemia salina, and Jellyfish Aurelia aurita. Toxics, 11(8), 716. https://doi.org/10.3390/toxics11080716