


Emerging Contaminants: An Emerging Risk Factor for Diabetes Mellitus

 , and
, and 
Abstract
:1. Introduction
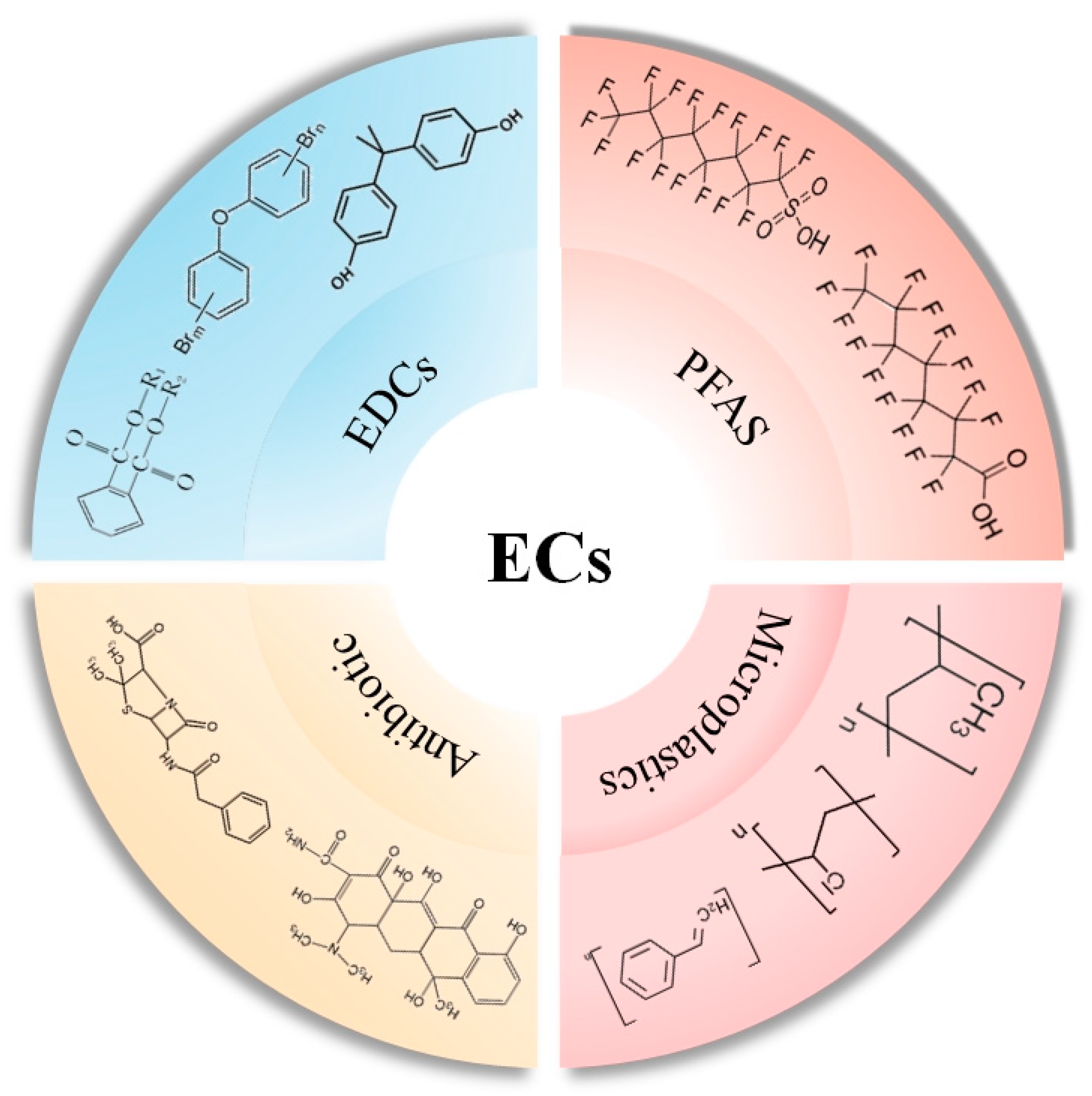
2. Emerging Contaminants (ECs)
3. Endocrine-Disrupting Chemicals (EDCs)
3.1. Persistent Organic Contaminants (POPs)
3.2. Bisphenol A (BPA) and Its Structural Analogs

{kind=link}
{kind=link}
| Class | Study Subject | Research Models | Chemical | Toxicity | Reference |
|---|---|---|---|---|---|
| Population Studies | General population (250 T2DM and 250 controls) | Case–control study | PAEs |
| [44] |
| General population (60 T2DM and 60 controls) | Case–control study | mPAEs, bisphenols |
| [45] | |
| First trimester (T1) pregnant women (60 GDM and 90 IGT) | Case–control study | PAEs |
| [46] | |
| First trimester (T1) pregnant women (169 GDM and 169 controls) | Case–control study | PAEs |
| [47] | |
| First trimester (T1) pregnant women (439 pregnant women) | Nested case–control study | PBDEs |
| [49] | |
| General population (71,415 women) | Prospective cohort study | Brominated flame retardants (BFRs) |
| [50] | |
| Cell Experiments | Rat insulinoma (INS-1) cells | Experimental research | DBP |
| [48] |
| Animal Experiments | Adult male C57BL/6J mice | Experimental research | BDE-153 |
| [51] |
| High-fat-diet male mice (C57BL/6J) | Experimental research | POPs |
| [52] | |
| High-fat-diet male mice (C57BL/6J) | Experimental research | TSP, D2O, PCB126 |
| [41] | |
| High-fat-diet female mice (C57BL/6J) | Experimental research | PCBs |
| [53] | |
| Low-fat-diet or high-fat-diet female mice (C57BL/6J) | Experimental research | PCB-77, PCB-126 |
| [54] | |
| Low-fat-diet or high-fat-diet male mice | Experimental research | PCB-77 |
| [55] | |
| High-fat-diet mice (C57BL/6J) | Experimental research | TCDD |
| [56] | |
| Male C57BL/6J mice and DBA/2J mice | Experimental research | TCDD |
| [57] | |
| Male C57BL/6J mice, AhR mice, islets from C57BL/6J mice | Experimental research | TCDD |
| [58] | |
| High-fat-diet female mice (C57BL/6J) | Experimental research | TCDD |
| [59] | |
| Male CD1 mice | Experimental research | BPA |
| [62] | |
| High-fat-diet mice (C57BL/6J) | Experimental research | BPA |
| [63] | |
| High-fat-diet male mice (C57BL/6J) | Experimental research | BPA |
| [64] |
4. Per- and Polyfluoroalkyl Substances (PFASs)
4.1. Perfluorooctane Sulfonate (PFOS)
4.2. Perfluorooctanoic Acid (PFOA)
| Class | Study Population | Research Models | Chemical | Toxicity | Reference |
|---|---|---|---|---|---|
| Population Studies | General population (474 adolescents and 969 Adults) | Cross-sectional study | PFNA, PFOS, PFOA, etc. |
| [72] |
| General population (1045 adults) | Cross-sectional study | PFOS and PFOA |
| [73] | |
| Pregnant women (171 GDM and 169 controls) | Cross-sectional study | PFOS, PFOA, PFNA, etc. |
| [74] | |
| General population (1016 men and women aged 70 years) | Prospective cohort study | PFOA, PFNA |
| [84] | |
| General population (100 participants) | Prospective cohort study | PFOA, PFOS |
| [85] | |
| Cell Experiments | GPR40-KO C57BL/6, C57BL/6 mice and mouse islet β-cells | Experimental research | PFOS |
| [76] |
| Beta-TC-6 pancreatic cells | Experimental research | PFOS |
| [77] | |
| Mouse pancreatic β-cells | Experimental research | PFOS |
| [78] | |
| Mouse pancreatic β-cell line (MIN6 cells) | Experimental research | PFOA |
| [87] | |
| Animal Experiments | Zebrafish (Danio rerio) embryos | Experimental research | PFOS |
| [75] |
| Pregnant mice and offspring | Experimental research | PFOS |
| [79] | |
| Adult male Balb/c mice | Experimental research | PFOA |
| [12] | |
| Male Balb/c | Experimental research | PFOA |
| [86] |
5. Microplastics
| Class | Study Population | Microplastics | Toxicity | Reference |
|---|---|---|---|---|
| Animal Experiments | Five-week-old high -fat-diet male mice (Mus musculus, ICR) | Polystyrene (5, 50, 100, and 200 μm) |
| [13] |
| High-fat-diet male mice (C57BL/6) | Polystyrene (0.45–0.53 μm) |
| [96] | |
| Mice | Polystyrene nanoplastics |
| [97] | |
| High-fat-diet male mice (C57BL/6) | Polystyrene (80 nm) |
| [98] | |
| ICR mice | Polystyrene (1 μm) |
| [99] |
6. Antibiotics
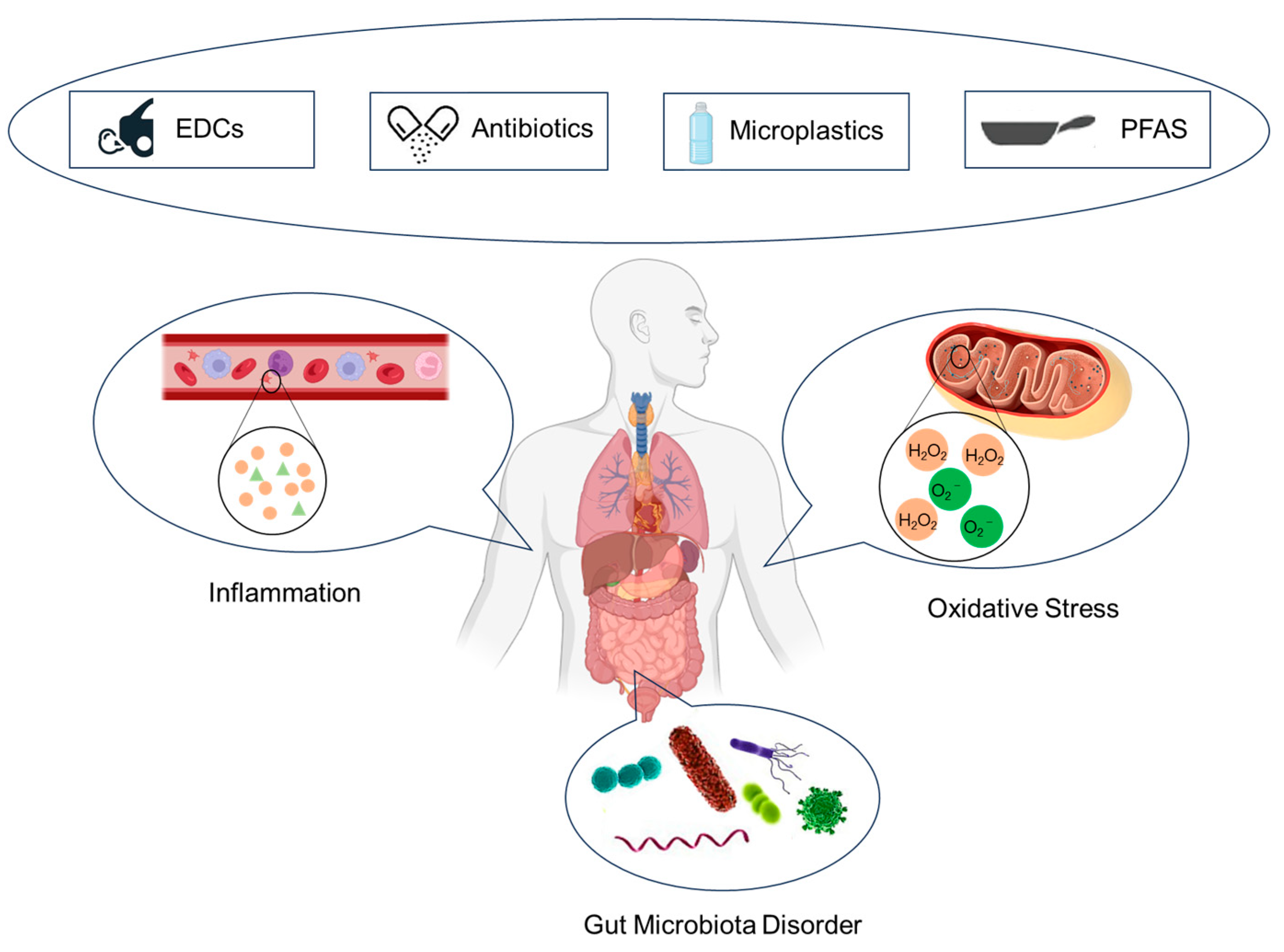
7. Role of Gut Microbiota
| Class | Species | Chemical | Changes in Intestinal Microbiota | Reference | |
|---|---|---|---|---|---|
| Animal Experiments | Female CD-1 mice | PFOS |
|
| [120] |
| ICR mice | Polystyrene microplastics (1 μm) |
|
| [99] | |
| Male ICR mice | Polystyrene microplastics (5, 50, 100, 200 μm) |
|
| [13] | |
| NOD/Shiltj mice | Antibiotics |
|
| [117] |
8. Conclusions
9. Future Research Directions
- i.
- The majority of current epidemiological studies focus on the relationship between the concentration of new pollutants in the serum or urine of the general population and fasting blood sugar, insulin, and glycated hemoglobin concentrations, with some studies involving changes in metabolic pathways related to glucose metabolism and their metabolites. However, studies on the impact of occupational exposure on diabetes, the specific exposure situations of new pollutants in the diet, the correlation with diabetes, and the impact of different geographical locations are relatively scarce. Diet is likely to be a very important exposure, yet rarely assessed in human studies, or when assessed, by questionnaires, often inaccurate. Quantifying the contribution of the human diet by multitargeted metabolomics of food and microbiota-derived metabolites may provide some clues. Therefore, more in-depth and targeted research is still needed to explore the impact of different factors on the development of diabetes.
- ii.
- The current research primarily focuses on the effects of novel environmental contaminants and their exposure on glucose metabolism in human populations, with fewer studies being directed towards animal models. Thus, there remains a pressing need for comprehensive studies to elucidate the specific mechanisms underlying the impact of these contaminants on diabetic or high-fat-diet populations, as well as the potential health outcomes from long-term low exposure.
- iii.
- Additionally, factors influencing the toxic effects of these novel contaminants, such as dose–response relationships, exposure frequency, gender disparities, and attributes like the type and size of the contaminants, have yet to be thoroughly investigated. Hence, there is an urgent need for more in-depth research into the toxicity of these new contaminants, factors modulating their toxicity levels, and their potential hazards. Such insights would furnish policymakers with a robust scientific foundation, aiding in the resolution of environmental challenges and the safeguarding of human health.
- iv.
- It is widely acknowledged that diabetes is influenced not only by genetic, environmental, and lifestyle factors but also by the structure and composition of the gut microbiota. However, current research on the interrelation between novel contaminants, gut microbiota, and diabetes is relatively scant. Consequently, determining whether exposure to these new contaminants might influence glucose metabolism by altering the gut microbiota’s structure and composition calls for relentless effort and exploration by researchers.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, Y.; Li, Y.; Zhuang, Z.; Song, Z.; Wang, W.; Huang, N.; Dong, X.; Xiao, W.; Jia, J.; Liu, Z.; et al. Associations of polysocial risk score, lifestyle and genetic factors with incident type 2 diabetes: A prospective cohort study. Diabetologia 2022, 65, 2056–2065. [Google Scholar] [CrossRef] [PubMed]
- Papazafiropoulou, A.K.; Papanas, N.; Melidonis, A.; Maltezos, E. Family History of Type 2 Diabetes: Does Having a Diabetic Parent Increase the Risk? Curr. Diabetes Rev. 2017, 13, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Qi, H.; Liu, J.; Zhang, G.; Liu, J.; Liu, N.; Zhu, M.; Zhao, X.; Song, C.; Zhou, Z.; et al. Deubiquitinase OTUD3 regulates metabolism homeostasis in response to nutritional stresses. Cell Metab. 2022, 34, 1023–1041. [Google Scholar] [CrossRef] [PubMed]
- Kolb, H.; Martin, S. Environmental/lifestyle factors in the pathogenesis and prevention of type 2 diabetes. BMC Med. 2017, 15, 131. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.W.; Novak, R.F.; Anderson, H.A.; Birnbaum, L.S.; Blystone, C.; Devito, M.; Jacobs, D.; Köhrle, J.; Lee, D.-H.; Rylander, L.; et al. Evaluation of the association between persistent organic pollutants (POPs) and diabetes in epidemiological studies: A national toxicology program workshop review. Environ. Health Perspect. 2013, 121, 774–783. [Google Scholar] [CrossRef] [PubMed]
- Prata, J.C.; da Costa, J.P.; Lopes, I.; Duarte, A.C.; Rocha-Santos, T. Environmental exposure to microplastics: An overview on possible human health effects. Sci. Total Environ. 2020, 702, 134455. [Google Scholar] [CrossRef]
- Han, D.; Currell, M.J. Persistent organic pollutants in China’s surface water systems. Sci. Total Environ. 2017, 580, 602–625. [Google Scholar] [CrossRef]
- Ouda, M.; Kadadou, D.; Swaidan, B.; Al-Othman, A.; Al-Asheh, S.; Banat, F.; Hasan, S.W. Emerging contaminants in the water bodies of the Middle East and North Africa (MENA): A critical review. Sci. Total Environ. 2021, 754, 142177. [Google Scholar] [CrossRef]
- Niu, H.; Liu, S.; Jiang, Y.; Hu, Y.; Li, Y.; He, L.; Xing, M.; Li, X.; Wu, L.; Chen, Z.; et al. Are Microplastics Toxic? A Review from Eco-Toxicity to Effects on the Gut Microbiota. Metabolites 2023, 13, 739. [Google Scholar] [CrossRef]
- Li, X.; Yin Yeung, L.W.; Xu, M.; Taniyasu, S.; Lam, P.K.S.; Yamashita, N.; Dai, J. Perfluorooctane sulfonate (PFOS) and other fluorochemicals in fish blood collected near the outfall of wastewater treatment plant (WWTP) in Beijing. Environ. Pollut. 2008, 156, 1298–1303. [Google Scholar] [CrossRef]
- Yeung, L.W.Y.; So, M.K.; Jiang, G.; Taniyasu, S.; Yamashita, N.; Song, M.; Wu, Y.; Li, J.; Giesy, J.P.; Guruge, K.S.; et al. Perfluorooctanesulfonate and related fluorochemicals in human blood samples from China. Environ. Sci. Technol. 2006, 40, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.; Sheng, N.; Zhang, H.; Yan, S.; Zhang, J.; Wang, J. Perfluorooctanoic acid exposure disturbs glucose metabolism in mouse liver. Toxicol. Appl. Pharmacol. 2017, 335, 41–48. [Google Scholar] [CrossRef]
- Huang, D.; Zhang, Y.; Long, J.; Yang, X.; Bao, L.; Yang, Z.; Wu, B.; Si, R.; Zhao, W.; Peng, C.; et al. Polystyrene microplastic exposure induces insulin resistance in mice via dysbacteriosis and pro-inflammation. Sci. Total Environ. 2022, 838, 155937. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Ding, K.; Huang, W.; Xu, F.; Lei, M.; Yue, R. Potential effects of bisphenol A on diabetes mellitus and its chronic complications: A narrative review. Heliyon 2023, 9, e16340. [Google Scholar] [CrossRef] [PubMed]
- Mariana, M.; Cairrao, E. The Relationship between Phthalates and Diabetes: A Review. Metabolites 2023, 13, 746. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.-J.; Wei, Y.-L.; Yao, Y.; Ruan, Q.-Q.; Zeng, E.Y. Global trends of research on emerging contaminants in the environment and humans: A literature assimilation. Environ. Sci. Pollut. Res. Int. 2015, 22, 1635–1643. [Google Scholar] [CrossRef] [PubMed]
- Naidu, R.; Arias Espana, V.A.; Liu, Y.; Jit, J. Emerging contaminants in the environment: Risk-based analysis for better management. Chemosphere 2016, 154, 350–357. [Google Scholar] [CrossRef]
- Khan, S.; Naushad, M.; Govarthanan, M.; Iqbal, J.; Alfadul, S.M. Emerging contaminants of high concern for the environment: Current trends and future research. Environ. Res. 2022, 207, 112609. [Google Scholar] [CrossRef]
- Mohammadi, A.; Dobaradaran, S.; Schmidt, T.C.; Malakootian, M.; Spitz, J. Emerging contaminants migration from pipes used in drinking water distribution systems: A review of the scientific literature. Environ. Sci. Pollut. Res. Int. 2022, 29, 75134–75160. [Google Scholar] [CrossRef]
- Puri, M.; Gandhi, K.; Kumar, M.S. Emerging environmental contaminants: A global perspective on policies and regulations. J. Environ. Manag. 2023, 332, 117344. [Google Scholar] [CrossRef]
- Chen, Y.; Lin, M.; Zhuang, D. Wastewater treatment and emerging contaminants: Bibliometric analysis. Chemosphere 2022, 297, 133932. [Google Scholar] [CrossRef] [PubMed]
- Bodus, B.; O’Malley, K.; Dieter, G.; Gunawardana, C.; McDonald, W. Review of emerging contaminants in green stormwater infrastructure: Antibiotic resistance genes, microplastics, tire wear particles, PFAS, and temperature. Sci. Total Environ. 2023, 906, 167195. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Shukla, P. Microalgal-based bioremediation of emerging contaminants: Mechanisms and challenges. Environ. Pollut. 2023, 337, 122591. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Mohapatra, S.; Zhang, J.; Tran, N.H.; You, L.; He, Y.; Gin, K.Y.-H. Source, fate, transport and modelling of selected emerging contaminants in the aquatic environment: Current status and future perspectives. Water Res. 2022, 217, 118418. [Google Scholar] [CrossRef] [PubMed]
- Dodoo, D.K.; Essumang, D.K.; Jonathan, J.W.A. Accumulation profile and seasonal variations of polychlorinated biphenyls (PCBs) in bivalves Crassostrea tulipa (oysters) and Anadara senilis (mussels) at three different aquatic habitats in two seasons in Ghana. Ecotoxicol. Environ. Saf. 2013, 88, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Bigler, J.; Greene, A. Guidance for Assessing Chemical Contaminant Data for Use in Fish Advisories; US EPA Office of Water, Office of Science and Technology: Washington, DC, USA, 1993; Volume 2.
- Wang, J.; Shi, Y.; Pan, Y.; Cai, Y. Perfluorooctane sulfonate (PFOS) and other fluorochemicals in viscera and muscle of farmed pigs and chickens in Beijing, China. Chin. Sci. Bull. 2010, 55, 3550–3555. [Google Scholar] [CrossRef]
- Yang, D.; Shi, H.; Li, L.; Li, J.; Jabeen, K.; Kolandhasamy, P. Microplastic Pollution in Table Salts from China. Environ. Sci. Technol. 2015, 49, 13622–13627. [Google Scholar] [CrossRef]
- Mason, S.A.; Welch, V.G.; Neratko, J. Synthetic Polymer Contamination in Bottled Water. Front. Chem. 2018, 6, 407. [Google Scholar] [CrossRef]
- Chen, X.; Wang, S.; Mao, X.; Xiang, X.; Ye, S.; Chen, J.; Zhu, A.; Meng, Y.; Yang, X.; Peng, S.; et al. Adverse health effects of emerging contaminants on inflammatory bowel disease. Front. Public. Health 2023, 11, 1140786. [Google Scholar] [CrossRef]
- Snow, D.D.; Cassada, D.A.; Biswas, S.; Malakar, A.; D’Alessio, M.; Marshall, A.H.L.; Sallach, J.B. Detection, occurrence, and fate of emerging contaminants in agricultural environments (2020). Water Environ. Res. 2020, 92, 1741–1750. [Google Scholar] [CrossRef]
- Alonso-Magdalena, P.; Quesada, I.; Nadal, A. Endocrine disruptors in the etiology of type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2011, 7, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Metcalfe, C.D.; Bayen, S.; Desrosiers, M.; Muñoz, G.; Sauvé, S.; Yargeau, V. Methods for the analysis of endocrine disrupting chemicals in selected environmental matrixes. Environ. Res. 2022, 206, 112616. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Sha, J.; Li, Z.; Wang, W.; Zhang, H. High affinity truncated aptamers for ultra-sensitive colorimetric detection of bisphenol A with label-free aptasensor. Food Chem. 2020, 317, 126459. [Google Scholar] [CrossRef] [PubMed]
- Jebril, S.; Cubillana-Aguilera, L.; Palacios-Santander, J.M.; Dridi, C. A novel electrochemical sensor modified with green gold sononanoparticles and carbon black nanocomposite for bisphenol A detection. Mater. Sci. Eng. B 2021, 264, 114951. [Google Scholar] [CrossRef]
- Lind, P.M.; Lind, L. Endocrine-disrupting chemicals and risk of diabetes: An evidence-based review. Diabetologia 2018, 61, 1495–1502. [Google Scholar] [CrossRef] [PubMed]
- Porta, M.; Gasull, M.; Puigdomènech, E.; Garí, M.; Bosch de Basea, M.; Guillén, M.; López, T.; Bigas, E.; Pumarega, J.; Llebaria, X.; et al. Distribution of blood concentrations of persistent organic pollutants in a representative sample of the population of Catalonia. Environ. Int. 2010, 36, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Kassotis, C.D.; Vandenberg, L.N.; Demeneix, B.A.; Porta, M.; Slama, R.; Trasande, L. Endocrine-disrupting chemicals: Economic, regulatory, and policy implications. Lancet Diabetes Endocrinol. 2020, 8, 719–730. [Google Scholar] [CrossRef]
- Fisher, B.E. Most unwanted. Environ. Health Perspect. 1999, 107, A18–A23. [Google Scholar] [CrossRef]
- Lee, Y.M.; Kim, K.S.; Jacobs, D.R.; Lee, D.H. Persistent organic pollutants in adipose tissue should be considered in obesity research. Obes. Rev. 2017, 18, 129–139. [Google Scholar] [CrossRef]
- Tian, Y.; Rimal, B.; Gui, W.; Koo, I.; Smith, P.B.; Yokoyama, S.; Patterson, A.D. Early Life Polychlorinated Biphenyl 126 Exposure Disrupts Gut Microbiota and Metabolic Homeostasis in Mice Fed with High-Fat Diet in Adulthood. Metabolites 2022, 12, 894. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiao, Y.; Li, Z.; Tao, Y.; Yang, Y. Hazards of phthalates (PAEs) exposure: A review of aquatic animal toxicology studies. Sci. Total Environ. 2021, 771, 145418. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Li, Z.; Wang, H.; Liang, H. An overview of phthalate acid ester pollution in China over the last decade: Environmental occurrence and human exposure. Sci. Total Environ. 2018, 645, 1400–1409. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Sun, H.; Han, L.; Chen, L. Association between phthalate exposure and glycosylated hemoglobin, fasting glucose, and type 2 diabetes mellitus: A case-control study in China. Sci. Total Environ. 2019, 670, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Sun, H.; Yao, Y.; Han, L.; Chen, L. Perturbation of serum metabolome in relation to type 2 diabetes mellitus and urinary levels of phthalate metabolites and bisphenols. Environ. Int. 2021, 155, 106609. [Google Scholar] [CrossRef] [PubMed]
- Shaffer, R.M.; Ferguson, K.K.; Sheppard, L.; James-Todd, T.; Butts, S.; Chandrasekaran, S.; Swan, S.H.; Barrett, E.S.; Nguyen, R.; Bush, N.; et al. Maternal urinary phthalate metabolites in relation to gestational diabetes and glucose intolerance during pregnancy. Environ. Int. 2019, 123, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; He, C.; Liu, X.; An, S.; Wang, X.; Tao, L.; Zhang, H.; Tian, Y.; Wu, N.; Xu, P.; et al. Effects of exposure to phthalate during early pregnancy on gestational diabetes mellitus: A nested case-control study with propensity score matching. Environ. Sci. Pollut. Res. Int. 2023, 30, 33555–33566. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Zheng, J.; Qin, J.; Liu, S.; Liu, X.; Gu, Y.; Yang, S.; Du, J.; Li, S.; Chen, B.; et al. Dibutyl phthalate affects insulin synthesis and secretion by regulating the mitochondrial apoptotic pathway and oxidative stress in rat insulinoma cells. Ecotoxicol. Environ. Saf. 2023, 249, 114396. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, L.; Li, J.; Meng, G.; Chi, M.; Li, T.; Zhao, Y.; Wu, Y. A nested case-control study of the association between exposure to polybrominated diphenyl ethers and the risk of gestational diabetes mellitus. Environ. Int. 2018, 119, 232–238. [Google Scholar] [CrossRef]
- Ongono, J.S.; Dow, C.; Gambaretti, J.; Severi, G.; Boutron-Ruault, M.-C.; Bonnet, F.; Fagherazzi, G.; Mancini, F.R. Dietary exposure to brominated flame retardants and risk of type 2 diabetes in the French E3N cohort. Environ. Int. 2019, 123, 54–60. [Google Scholar] [CrossRef]
- Liu, Z.-L.; Jiang, S.-R.; Fan, Y.; Wang, J.-S.; Wang, M.-L.; Li, M.-Y. 2,2’,4,4’,5,5’-Hexabromophenyl ether (BDE-153) causes abnormal insulin secretion and disorders of glucose and lipid metabolism in mice. J. Chin. Med. Assoc. 2023, 86, 388–398. [Google Scholar] [CrossRef]
- Ibrahim, M.M.; Fjære, E.; Lock, E.-J.; Naville, D.; Amlund, H.; Meugnier, E.; Le Magueresse Battistoni, B.; Frøyland, L.; Madsen, L.; Jessen, N.; et al. Chronic consumption of farmed salmon containing persistent organic pollutants causes insulin resistance and obesity in mice. PLoS ONE 2011, 6, e25170. [Google Scholar] [CrossRef] [PubMed]
- Gray, S.L.; Shaw, A.C.; Gagne, A.X.; Chan, H.M. Chronic exposure to PCBs (Aroclor 1254) exacerbates obesity-induced insulin resistance and hyperinsulinemia in mice. J. Toxicol. Environ. Health A 2013, 76, 701–715. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.A.; Karounos, M.; English, V.; Fang, J.; Wei, Y.; Stromberg, A.; Sunkara, M.; Morris, A.J.; Swanson, H.I.; Cassis, L.A. Coplanar polychlorinated biphenyls impair glucose homeostasis in lean C57BL/6 mice and mitigate beneficial effects of weight loss on glucose homeostasis in obese mice. Environ. Health Perspect. 2013, 121, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.A.; Shoemaker, R.; English, V.; Larian, N.; Sunkara, M.; Morris, A.J.; Walker, M.; Yiannikouris, F.; Cassis, L.A. Effects of Adipocyte Aryl Hydrocarbon Receptor Deficiency on PCB-Induced Disruption of Glucose Homeostasis in Lean and Obese Mice. Environ. Health Perspect. 2015, 123, 944–950. [Google Scholar] [CrossRef] [PubMed]
- Matteo, G.; Hoyeck, M.P.; Blair, H.L.; Zebarth, J.; Rick, K.R.C.; Williams, A.; Gagné, R.; Buick, J.K.; Yauk, C.L.; Bruin, J.E. Prolonged Low-Dose Dioxin Exposure Impairs Metabolic Adaptability to High-Fat Diet Feeding in Female but Not Male Mice. Endocrinology 2021, 162, bqab050. [Google Scholar] [CrossRef] [PubMed]
- Ishida, T.; Kan-o, S.; Mutoh, J.; Takeda, S.; Ishii, Y.; Hashiguchi, I.; Akamine, A.; Yamada, H. 2,3,7,8-Tetrachlorodibenzo-p-dioxin-induced change in intestinal function and pathology: Evidence for the involvement of arylhydrocarbon receptor-mediated alteration of glucose transportation. Toxicol. Appl. Pharmacol. 2005, 205, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Kurita, H.; Yoshioka, W.; Nishimura, N.; Kubota, N.; Kadowaki, T.; Tohyama, C. Aryl hydrocarbon receptor-mediated effects of 2,3,7,8-tetrachlorodibenzo-p-dioxin on glucose-stimulated insulin secretion in mice. J. Appl. Toxicol. 2009, 29, 689–694. [Google Scholar] [CrossRef]
- Hoyeck, M.P.; Merhi, R.C.; Blair, H.L.; Spencer, C.D.; Payant, M.A.; Martin Alfonso, D.I.; Zhang, M.; Matteo, G.; Chee, M.J.; Bruin, J.E. Female mice exposed to low doses of dioxin during pregnancy and lactation have increased susceptibility to diet-induced obesity and diabetes. Mol. Metab. 2020, 42, 101104. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Hauser, R.; Marcus, M.; Olea, N.; Welshons, W.V. Human exposure to bisphenol A (BPA). Reprod. Toxicol. 2007, 24, 139–177. [Google Scholar] [CrossRef]
- Akash, M.S.H.; Sabir, S.; Rehman, K. Bisphenol A-induced metabolic disorders: From exposure to mechanism of action. Environ. Toxicol. Pharmacol. 2020, 77, 103373. [Google Scholar] [CrossRef]
- Marmugi, A.; Lasserre, F.; Beuzelin, D.; Ducheix, S.; Huc, L.; Polizzi, A.; Chetivaux, M.; Pineau, T.; Martin, P.; Guillou, H.; et al. Adverse effects of long-term exposure to bisphenol A during adulthood leading to hyperglycaemia and hypercholesterolemia in mice. Toxicology 2014, 325, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Deng, P.; Lin, M.; Yang, L.; Li, L.; Guo, L.; Zhang, L.; He, M.; Lu, Y.; Pi, H.; et al. Long-term bisphenol A exposure exacerbates diet-induced prediabetes via TLR4-dependent hypothalamic inflammation. J. Hazard. Mater. 2021, 402, 123926. [Google Scholar] [CrossRef] [PubMed]
- Moon, M.K.; Jeong, I.-K.; Jung Oh, T.; Ahn, H.Y.; Kim, H.H.; Park, Y.J.; Jang, H.C.; Park, K.S. Long-term oral exposure to bisphenol A induces glucose intolerance and insulin resistance. J. Endocrinol. 2015, 226, 35–42. [Google Scholar] [CrossRef]
- Lau, C.; Anitole, K.; Hodes, C.; Lai, D.; Pfahles-Hutchens, A.; Seed, J. Perfluoroalkyl acids: A review of monitoring and toxicological findings. Toxicol. Sci. 2007, 99, 366–394. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; DeWitt, J.C.; Higgins, C.P.; Cousins, I.T. A Never-Ending Story of Per- and Polyfluoroalkyl Substances (PFASs)? Environ. Sci. Technol. 2017, 51, 2508–2518. [Google Scholar] [CrossRef] [PubMed]
- Domingo, J.L.; Nadal, M. Human exposure to per- and polyfluoroalkyl substances (PFAS) through drinking water: A review of the recent scientific literature. Environ. Res. 2019, 177, 108648. [Google Scholar] [CrossRef]
- Lee, J.-W.; Lee, H.-K.; Lim, J.-E.; Moon, H.-B. Legacy and emerging per- and polyfluoroalkyl substances (PFASs) in the coastal environment of Korea: Occurrence, spatial distribution, and bioaccumulation potential. Chemosphere 2020, 251, 126633. [Google Scholar] [CrossRef]
- Wan, H.T.; Zhao, Y.G.; Wei, X.; Hui, K.Y.; Giesy, J.P.; Wong, C.K.C. PFOS-induced hepatic steatosis, the mechanistic actions on β-oxidation and lipid transport. Biochim. Biophys. Acta 2012, 1820, 1092–1101. [Google Scholar] [CrossRef]
- Sanan, T.; Magnuson, M. Analysis of per- and polyfluorinated alkyl substances in sub-sampled water matrices with online solid phase extraction/isotope dilution tandem mass spectrometry. J. Chromatogr. A 2020, 1626, 461324. [Google Scholar] [CrossRef]
- Olsen, G.W.; Burris, J.M.; Ehresman, D.J.; Froehlich, J.W.; Seacat, A.M.; Butenhoff, J.L.; Zobel, L.R. Half-life of serum elimination of perfluorooctanesulfonate, perfluorohexanesulfonate, and perfluorooctanoate in retired fluorochemical production workers. Environ. Health Perspect. 2007, 115, 1298–1305. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Chen, P.-C.; Lin, Y.-C.; Lin, L.-Y. Association among serum perfluoroalkyl chemicals, glucose homeostasis, and metabolic syndrome in adolescents and adults. Diabetes Care 2009, 32, 702–707. [Google Scholar] [CrossRef] [PubMed]
- Zeeshan, M.; Zhang, Y.-T.; Yu, S.; Huang, W.-Z.; Zhou, Y.; Vinothkumar, R.; Chu, C.; Li, Q.-Q.; Wu, Q.-Z.; Ye, W.-L.; et al. Exposure to isomers of per- and polyfluoroalkyl substances increases the risk of diabetes and impairs glucose-homeostasis in Chinese adults: Isomers of C8 health project. Chemosphere 2021, 278, 130486. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zhang, L.; Zhou, Q.; Ding, J.; Yin, S.; Shang, X.; Tian, Y. Exposure to per- and polyfluoroalkyl substances as a risk factor for gestational diabetes mellitus through interference with glucose homeostasis. Sci. Total Environ. 2022, 838, 156561. [Google Scholar] [CrossRef] [PubMed]
- Sant, K.E.; Jacobs, H.M.; Borofski, K.A.; Moss, J.B.; Timme-Laragy, A.R. Embryonic exposures to perfluorooctanesulfonic acid (PFOS) disrupt pancreatic organogenesis in the zebrafish, Danio rerio. Environ. Pollut. 2017, 220, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.-P.; Cao, L.-Y.; Li, C.-H.; Guo, L.-H.; Colbourne, J.; Ren, X.-M. Perfluoroalkyl Substances Stimulate Insulin Secretion by Islet β Cells via G Protein-Coupled Receptor 40. Environ. Sci. Technol. 2020, 54, 3428–3436. [Google Scholar] [CrossRef]
- Zhang, L.; Duan, X.; Sun, W.; Sun, H. Perfluorooctane sulfonate acute exposure stimulates insulin secretion via GPR40 pathway. Sci. Total Environ. 2020, 726, 138498. [Google Scholar] [CrossRef]
- Duan, X.; Sun, W.; Sun, H.; Zhang, L. Perfluorooctane sulfonate continual exposure impairs glucose-stimulated insulin secretion via SIRT1-induced upregulation of UCP2 expression. Environ. Pollut. 2021, 278, 116840. [Google Scholar] [CrossRef]
- Wan, H.T.; Zhao, Y.G.; Leung, P.Y.; Wong, C.K.C. Perinatal exposure to perfluorooctane sulfonate affects glucose metabolism in adult offspring. PLoS ONE 2014, 9, e87137. [Google Scholar] [CrossRef]
- Harada, K.; Nakanishi, S.; Sasaki, K.; Furuyama, K.; Nakayama, S.; Saito, N.; Yamakawa, K.; Koizumi, A. Particle size distribution and respiratory deposition estimates of airborne perfluorooctanoate and perfluorooctanesulfonate in Kyoto area, Japan. Bull. Environ. Contam. Toxicol. 2006, 76, 306–310. [Google Scholar] [CrossRef]
- Barton, C.A.; Butler, L.E.; Zarzecki, C.J.; Flaherty, J.; Kaiser, M. Characterizing perfluorooctanoate in ambient air near the fence line of a manufacturing facility: Comparing modeled and monitored values. J. Air Waste Manag. Assoc. 2006, 56, 48–55. [Google Scholar] [CrossRef]
- Davis, K.L.; Aucoin, M.D.; Larsen, B.S.; Kaiser, M.A.; Hartten, A.S. Transport of ammonium perfluorooctanoate in environmental media near a fluoropolymer manufacturing facility. Chemosphere 2007, 67, 2011–2019. [Google Scholar] [CrossRef] [PubMed]
- Ehresman, D.J.; Froehlich, J.W.; Olsen, G.W.; Chang, S.-C.; Butenhoff, J.L. Comparison of human whole blood, plasma, and serum matrices for the determination of perfluorooctanesulfonate (PFOS), perfluorooctanoate (PFOA), and other fluorochemicals. Environ. Res. 2007, 103, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Lind, L.; Zethelius, B.; Salihovic, S.; van Bavel, B.; Lind, P.M. Circulating levels of perfluoroalkyl substances and prevalent diabetes in the elderly. Diabetologia 2014, 57, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.M.; Heo, D.-G.; Kim, J.-H.; Yoon, J.S.; Lee, H.W.; Kim, J.-Y.; Moon, J.S.; Won, K.C. Perfluorinated compounds in adults and their association with fasting glucose and incident diabetes: A prospective cohort study. Environ. Health 2022, 21, 101. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Zhang, H.; Zheng, F.; Sheng, N.; Guo, X.; Dai, J. Perfluorooctanoic acid exposure for 28 days affects glucose homeostasis and induces insulin hypersensitivity in mice. Sci. Rep. 2015, 5, 11029. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Wu, D.; Xu, Y.; Zhang, Y.; Sun, Y.; Chang, X.; Zhu, Y.; Tang, W. Perfluorooctanoic acid promotes pancreatic β cell dysfunction and apoptosis through ER stress and the ATF4/CHOP/TRIB3 pathway. Environ. Sci. Pollut. Res. Int. 2022, 29, 84532–84545. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Yu, S.; Zhu, X.; Liao, R.; Zhuo, Z.; He, Y.; Ma, H. In-situ Detection Method for Microplastics in Water by Polarized Light Scattering. Front. Mar. Sci. 2021, 8, 739683. [Google Scholar] [CrossRef]
- Dey, T.K.; Uddin, M.E.; Jamal, M. Detection and removal of microplastics in wastewater: Evolution and impact. Environ. Sci. Pollut. Res. Int. 2021, 28, 16925–16947. [Google Scholar] [CrossRef]
- Yong, C.Q.Y.; Valiyaveettil, S.; Tang, B.L. Toxicity of Microplastics and Nanoplastics in Mammalian Systems. Int. J. Environ. Res. Public. Health 2020, 17, 1509. [Google Scholar] [CrossRef]
- Kumar, R.; Manna, C.; Padha, S.; Verma, A.; Sharma, P.; Dhar, A.; Ghosh, A.; Bhattacharya, P. Micro(nano)plastics pollution and human health: How plastics can induce carcinogenesis to humans? Chemosphere 2022, 298, 134267. [Google Scholar] [CrossRef]
- Xia, X.; Sun, M.; Zhou, M.; Chang, Z.; Li, L. Polyvinyl chloride microplastics induce growth inhibition and oxidative stress in Cyprinus carpio var. larvae. Sci. Total Environ. 2020, 716, 136479. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhang, Y.; Deng, Y.; Jiang, W.; Zhao, Y.; Geng, J.; Ding, L.; Ren, H. Uptake and Accumulation of Polystyrene Microplastics in Zebrafish (Danio rerio) and Toxic Effects in Liver. Environ. Sci. Technol. 2016, 50, 4054–4060. [Google Scholar] [CrossRef] [PubMed]
- Cortés, C.; Domenech, J.; Salazar, M.; Pastor, S.; Marcos, R.; Hernández, A. Nanoplastics as a potential environmental health factor: Effects of polystyrene nanoparticles on human intestinal epithelial Caco-2 cells. Environ. Sci. Nano 2020, 7, 272–285. [Google Scholar] [CrossRef]
- Schirinzi, G.F.; Pérez-Pomeda, I.; Sanchís, J.; Rossini, C.; Farré, M.; Barceló, D. Cytotoxic effects of commonly used nanomaterials and microplastics on cerebral and epithelial human cells. Environ. Res. 2017, 159, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Okamura, T.; Hamaguchi, M.; Hasegawa, Y.; Hashimoto, Y.; Majima, S.; Senmaru, T.; Ushigome, E.; Nakanishi, N.; Asano, M.; Yamazaki, M.; et al. Oral Exposure to Polystyrene Microplastics of Mice on a Normal or High-Fat Diet and Intestinal and Metabolic Outcomes. Environ. Health Perspect. 2023, 131, 27006. [Google Scholar] [CrossRef]
- Fan, X.; Wei, X.; Hu, H.; Zhang, B.; Yang, D.; Du, H.; Zhu, R.; Sun, X.; Oh, Y.; Gu, N. Effects of oral administration of polystyrene nanoplastics on plasma glucose metabolism in mice. Chemosphere 2022, 288, 132607. [Google Scholar] [CrossRef]
- Wang, Y.; Wei, Z.; Xu, K.; Wang, X.; Gao, X.; Han, Q.; Wang, S.; Chen, M. The effect and a mechanistic evaluation of polystyrene nanoplastics on a mouse model of type 2 diabetes. Food Chem. Toxicol. 2023, 173, 113642. [Google Scholar] [CrossRef]
- Shi, C.; Han, X.; Guo, W.; Wu, Q.; Yang, X.; Wang, Y.; Tang, G.; Wang, S.; Wang, Z.; Liu, Y.; et al. Disturbed Gut-Liver axis indicating oral exposure to polystyrene microplastic potentially increases the risk of insulin resistance. Environ. Int. 2022, 164, 107273. [Google Scholar] [CrossRef]
- Klein, E.Y.; Van Boeckel, T.P.; Martinez, E.M.; Pant, S.; Gandra, S.; Levin, S.A.; Goossens, H.; Laxminarayan, R. Global increase and geographic convergence in antibiotic consumption between 2000 and 2015. Proc. Natl. Acad. Sci. USA 2018, 115, E3463–E3470. [Google Scholar] [CrossRef]
- Van Boeckel, T.P.; Gandra, S.; Ashok, A.; Caudron, Q.; Grenfell, B.T.; Levin, S.A.; Laxminarayan, R. Global antibiotic consumption 2000 to 2010: An analysis of national pharmaceutical sales data. Lancet Infect. Dis. 2014, 14, 742–750. [Google Scholar] [CrossRef]
- Tran, N.H.; Chen, H.; Reinhard, M.; Mao, F.; Gin, K.Y.-H. Occurrence and removal of multiple classes of antibiotics and antimicrobial agents in biological wastewater treatment processes. Water Res. 2016, 104, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Lu, N.; Chen, J.; Rao, Z.; Guo, B.; Xu, Y. Recent Advances of Biosensors for Detection of Multiple Antibiotics. Biosensors 2023, 13, 850. [Google Scholar] [CrossRef] [PubMed]
- Gago-Ferrero, P.; Díaz-Cruz, M.S.; Barceló, D. Occurrence of multiclass UV filters in treated sewage sludge from wastewater treatment plants. Chemosphere 2011, 84, 1158–1165. [Google Scholar] [CrossRef] [PubMed]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of gut microbiota in type 2 diabetes pathophysiology. eBioMedicine 2020, 51, 102590. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, R.R.; Greer, R.L.; Dong, X.; Dsouza, K.N.; Gurung, M.; Wu, J.Y.; Morgun, A.; Shulzhenko, N. Antibiotic-Induced Alterations in Gut Microbiota Are Associated with Changes in Glucose Metabolism in Healthy Mice. Front. Microbiol. 2017, 8, 2306. [Google Scholar] [CrossRef]
- Yuan, J.; Hu, Y.J.; Zheng, J.; Kim, J.H.; Sumerlin, T.; Chen, Y.; He, Y.; Zhang, C.; Tang, J.; Pan, Y.; et al. Long-term use of antibiotics and risk of type 2 diabetes in women: A prospective cohort study. Int. J. Epidemiol. 2020, 49, 1572–1581. [Google Scholar] [CrossRef]
- Fu, L.; Qiu, Y.; Shen, L.; Cui, C.; Wang, S.; Wang, S.; Xie, Y.; Zhao, X.; Gao, X.; Ning, G.; et al. The delayed effects of antibiotics in type 2 diabetes, friend or foe? J. Endocrinol. 2018, 238, 137–149. [Google Scholar] [CrossRef]
- Fenneman, A.C.; Weidner, M.; Chen, L.A.; Nieuwdorp, M.; Blaser, M.J. Antibiotics in the pathogenesis of diabetes and inflammatory diseases of the gastrointestinal tract. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 81–100. [Google Scholar] [CrossRef]
- Buffie, C.G.; Bucci, V.; Stein, R.R.; McKenney, P.T.; Ling, L.; Gobourne, A.; No, D.; Liu, H.; Kinnebrew, M.; Viale, A.; et al. Precision microbiome reconstitution restores bile acid mediated resistance to Clostridium difficile. Nature 2015, 517, 205–208. [Google Scholar] [CrossRef]
- Sharon, G.; Garg, N.; Debelius, J.; Knight, R.; Dorrestein, P.C.; Mazmanian, S.K. Specialized metabolites from the microbiome in health and disease. Cell Metab. 2014, 20, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed]
- Claus, S.P.; Guillou, H.; Ellero-Simatos, S. The gut microbiota: A major player in the toxicity of environmental pollutants? NPJ Biofilms Microbiomes 2016, 2, 16003. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Abo, R.P.; Schlieper, K.A.; Graffam, M.E.; Levine, S.; Wishnok, J.S.; Swenberg, J.A.; Tannenbaum, S.R.; Fox, J.G. Arsenic exposure perturbs the gut microbiome and its metabolic profile in mice: An integrated metagenomics and metabolomics analysis. Environ. Health Perspect. 2014, 122, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Bian, X.; Tu, P.; Chi, L.; Gao, B.; Ru, H.; Lu, K. Saccharin induced liver inflammation in mice by altering the gut microbiota and its metabolic functions. Food Chem. Toxicol. 2017, 107, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Livanos, A.E.; Greiner, T.U.; Vangay, P.; Pathmasiri, W.; Stewart, D.; McRitchie, S.; Li, H.; Chung, J.; Sohn, J.; Kim, S.; et al. Antibiotic-mediated gut microbiome perturbation accelerates development of type 1 diabetes in mice. Nat. Microbiol. 2016, 1, 16140. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, S.; Härkönen, T.; Vatanen, T.; Hyötyläinen, T.; Knip, M.; Orešič, M. Impact of exposure to per- and polyfluoroalkyl substances on fecal microbiota composition in mother-infant dyads. Environ. Int. 2023, 176, 107965. [Google Scholar] [CrossRef]
- Potera, C. POPs and Gut Microbiota: Dietary Exposure Alters Ratio of Bacterial Species. Environ. Health Perspect. 2015, 123, A187. [Google Scholar] [CrossRef]
- Lai, K.P.; Ng, A.H.-M.; Wan, H.T.; Wong, A.Y.-M.; Leung, C.C.-T.; Li, R.; Wong, C.K.-C. Dietary Exposure to the Environmental Chemical, PFOS on the Diversity of Gut Microbiota, Associated With the Development of Metabolic Syndrome. Front. Microbiol. 2018, 9, 2552. [Google Scholar] [CrossRef]
- Crawford, P.A.; Crowley, J.R.; Sambandam, N.; Muegge, B.D.; Costello, E.K.; Hamady, M.; Knight, R.; Gordon, J.I. Regulation of myocardial ketone body metabolism by the gut microbiota during nutrient deprivation. Proc. Natl. Acad. Sci. USA 2009, 106, 11276–11281. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, H.; Xu, M.; Tu, P.; Xu, Y.; Li, X.; Xing, M.; Chen, Z.; Wang, X.; Lou, X.; Wu, L.; et al. Emerging Contaminants: An Emerging Risk Factor for Diabetes Mellitus. Toxics 2024, 12, 47. https://doi.org/10.3390/toxics12010047
Niu H, Xu M, Tu P, Xu Y, Li X, Xing M, Chen Z, Wang X, Lou X, Wu L, et al. Emerging Contaminants: An Emerging Risk Factor for Diabetes Mellitus. Toxics. 2024; 12(1):47. https://doi.org/10.3390/toxics12010047
Chicago/Turabian StyleNiu, Huixia, Manjin Xu, Pengcheng Tu, Yunfeng Xu, Xueqing Li, Mingluan Xing, Zhijian Chen, Xiaofeng Wang, Xiaoming Lou, Lizhi Wu, and et al. 2024. "Emerging Contaminants: An Emerging Risk Factor for Diabetes Mellitus" Toxics 12, no. 1: 47. https://doi.org/10.3390/toxics12010047
APA StyleNiu, H., Xu, M., Tu, P., Xu, Y., Li, X., Xing, M., Chen, Z., Wang, X., Lou, X., Wu, L., & Sun, S. (2024). Emerging Contaminants: An Emerging Risk Factor for Diabetes Mellitus. Toxics, 12(1), 47. https://doi.org/10.3390/toxics12010047