
Perinatal Tetrahydrocannabinol Compromises Maternal Care and Increases Litter Attrition in the Long–Evans Rat

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. THC Dosing Solution Preparation
2.2. Breeding and THC Administration
2.3. Developmental Milestones
2.4. Maternal Pup Retrieval
2.5. Blood Sampling and THC Assays
2.6. Statistical Methods
3. Results
3.1. Gestational Body Weight
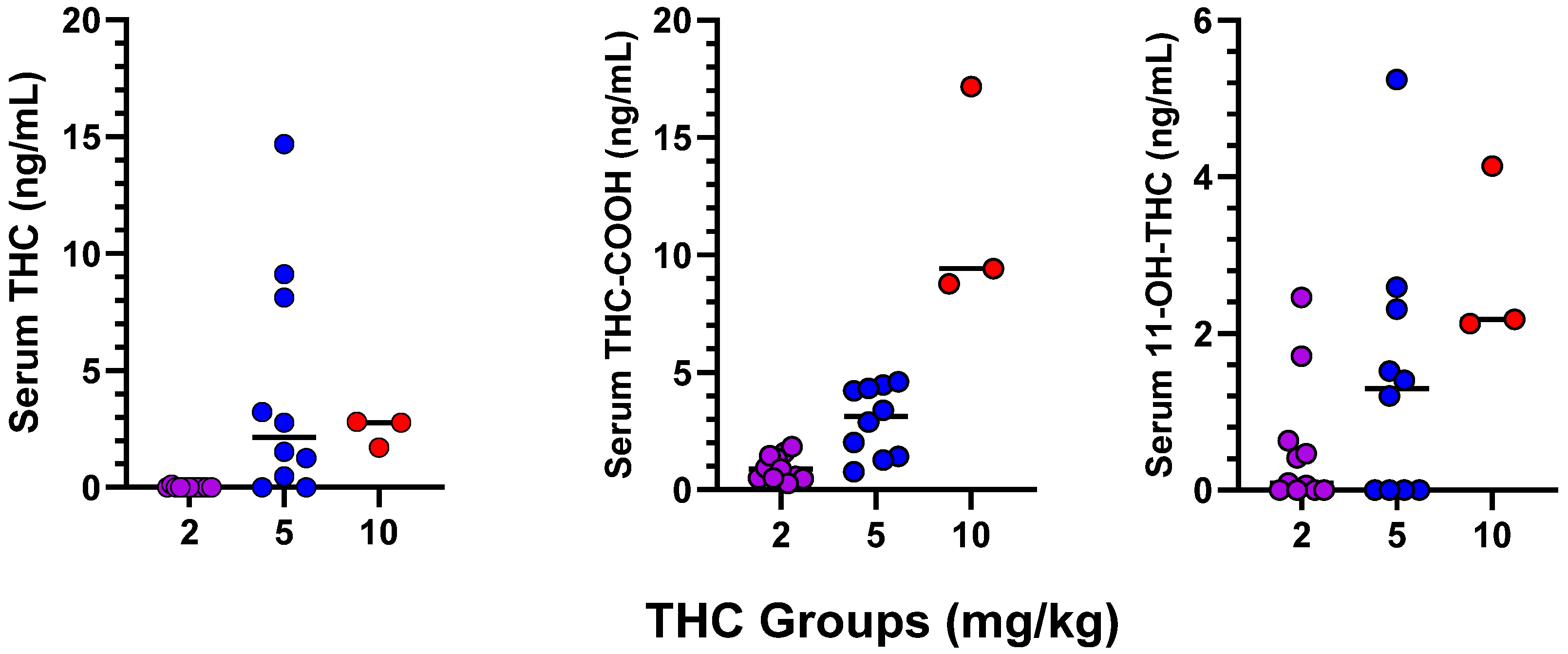
3.2. Serum Levels of THC and Metabolites in Dams
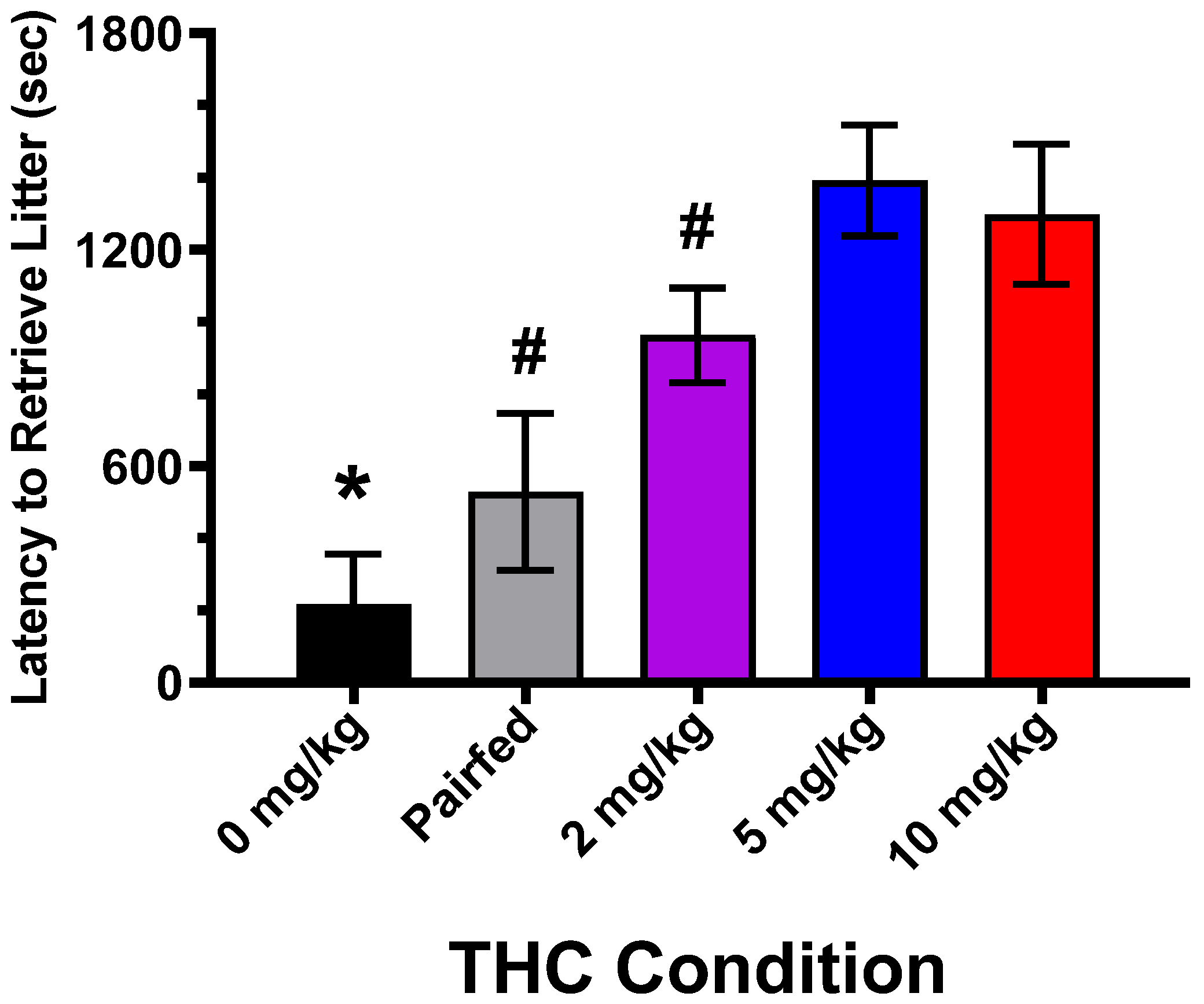
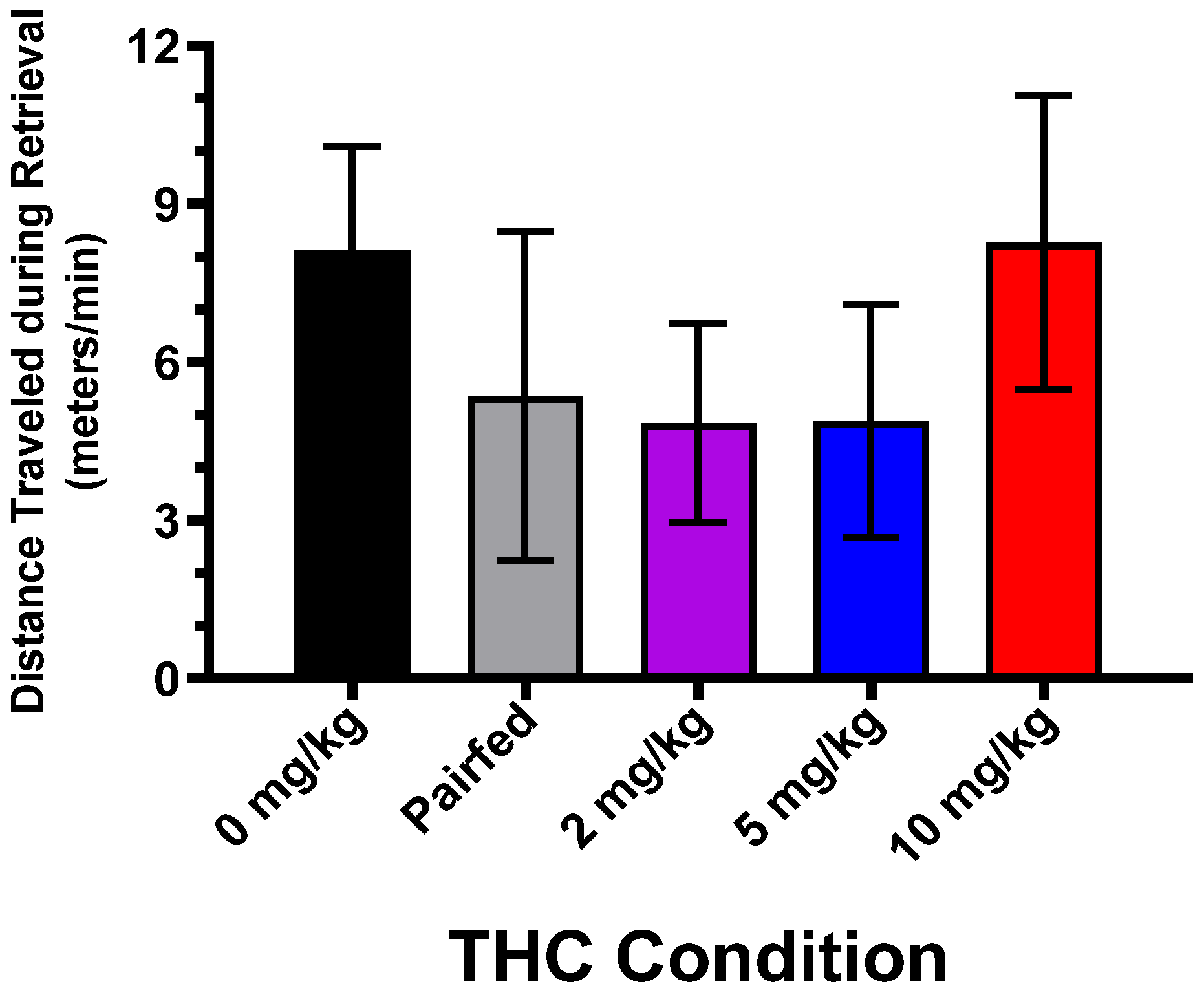
3.3. Maternal Pup Retrieval
3.4. Effects of THC on Pup Developmental Milestones
3.4.1. Body Growth
3.4.2. Eye Opening
3.4.3. Righting Reflex
3.5. Effects of Impaired Maternal Care on Pup Mortality
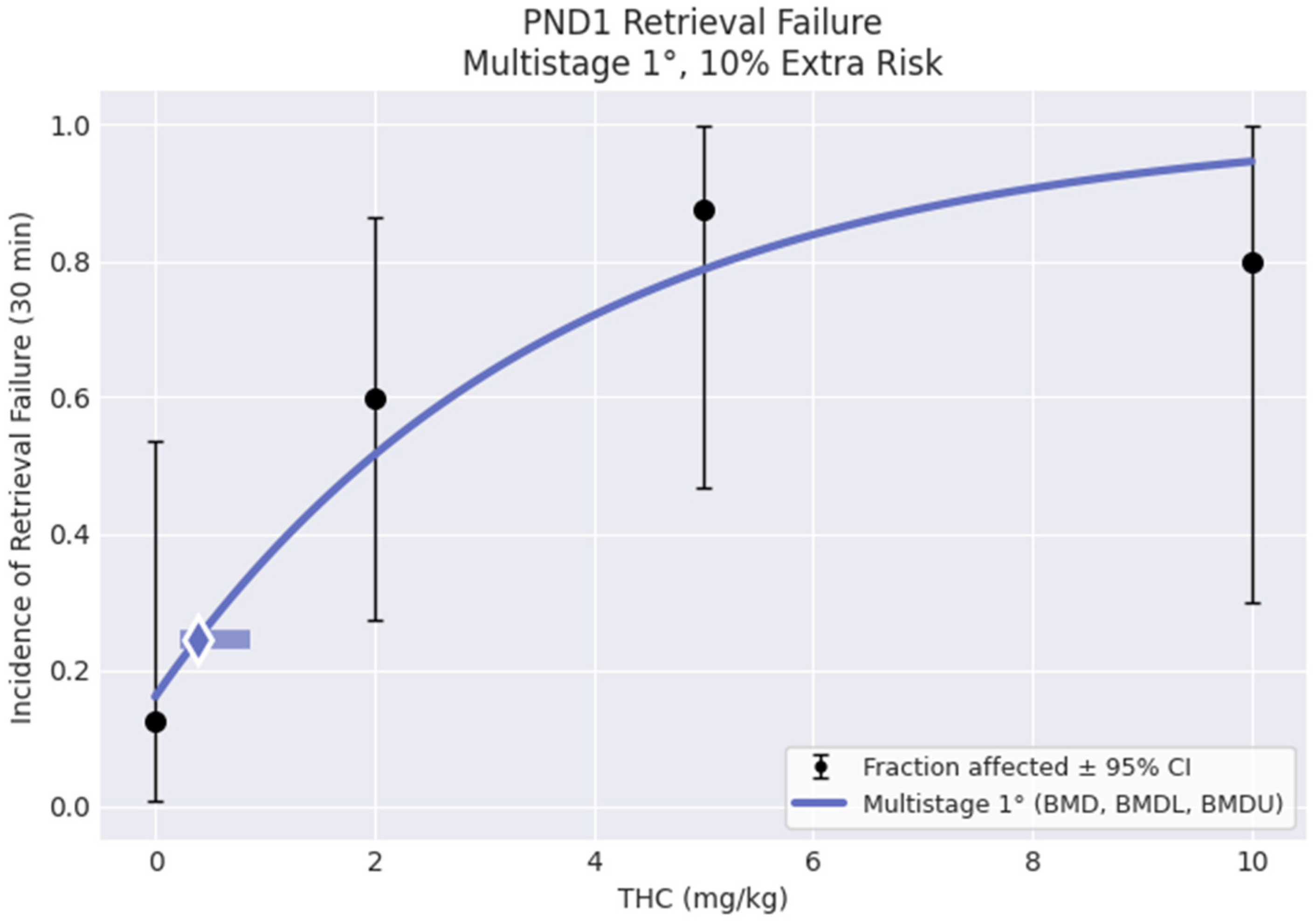
3.6. Benchmark Dose Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cameron, L.D.; Fleszar-Pavlović, S.E.; Yepez, M.; Manzo, R.D.; Brown, P.M. Beliefs about marijuana use during pregnancy and breastfeeding held by residents of a Latino-majority, rural region of California. J. Behav. Med. 2022, 45, 544–557. [Google Scholar] [CrossRef] [PubMed]
- Kvillemo, P.; Strandberg, A.K.; Gripenberg, J. Attitudes to Cannabis Use and Public Prevention Information Among Young Adults: A Qualitative Interview Study With Implications for Prevention Practice. Front. Public Health 2022, 10, 830201. [Google Scholar] [CrossRef] [PubMed]
- Palamar, J.J.; Ompad, D.C.; Petkova, E. Correlates of intentions to use cannabis among US high school seniors in the case of cannabis legalization. Int. J. Drug Policy 2014, 25, 424–435. [Google Scholar] [CrossRef] [PubMed]
- Calvigioni, D.; Hurd, Y.L.; Harkany, T.; Keimpema, E. Neuronal substrates and functional consequences of prenatal cannabis exposure. Eur. Child Adolesc. Psychiatry 2014, 23, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Craft, R.M.; Marusich, J.A.; Wiley, J.L. Sex differences in cannabinoid pharmacology: A reflection of differences in the endocannabinoid system? Life Sci. 2013, 92, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Besse, M.; Parikh, K.; Mark, K. Reported Reasons for Cannabis Use Before and After Pregnancy Recognition. J. Addict. Med. 2023, 17, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Foti, T.R.; Green, A.; Altschuler, A.; Iturralde, E.; Does, M.B.; Jackson-Morris, M.; Adams, S.R.; Goler, N.; Ansley, D.; Conway, A.; et al. Patient Perceptions of Prenatal Cannabis Use and Implications for Clinicians. Obstet. Gynecol. 2023, 142, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Murnan, A.W.; Keim, S.A.; Li, R.; Klebanoff, M.A. Marijuana use and sleep quality during pregnancy. J. Matern.-Fetal Neonatal Med. 2022, 35, 7857–7864. [Google Scholar] [CrossRef] [PubMed]
- Vanderziel, A.; Anthony, J.C.; Barondess, D.; Kerver, J.M.; Alshaarawy, O. Nausea and vomiting of pregnancy and prenatal cannabis use in a Michigan sample. Am. J. Obstet. Gynecol. MFM 2023, 5, 101171. [Google Scholar] [CrossRef] [PubMed]
- Bandoli, G.; Jelliffe-Pawlowski, L.; Schumacher, B.; Baer, R.J.; Felder, J.N.; Fuchs, J.D.; Oltman, S.P.; Steurer, M.A.; Marienfeld, C. Cannabis-related diagnosis in pregnancy and adverse maternal and infant outcomes. Drug Alcohol Depend. 2021, 225, 108757. [Google Scholar] [CrossRef]
- Klasner, C.; Brown, J.; Gopalakrishnan, M.; Metwally, D.E.; Besse, M.; Mark, K. Effect of maternal adverse childhood experiences (ACE) and cannabis use on pregnancy outcomes. Arch. Women’s Ment. Health 2022, 25, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Koto, P.; Allen, V.M.; Fahey, J.; Kuhle, S. Maternal cannabis use during pregnancy and maternal and neonatal outcomes: A retrospective cohort study. BJOG Int. J. Obstet. Gynaecol. 2022, 129, 1687–1694. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhu, B.; Liang, D. The associations between prenatal cannabis use disorder and neonatal outcomes. Addiction 2021, 116, 3069–3079. [Google Scholar] [CrossRef] [PubMed]
- Fried, P.A.; Smith, A.M. A literature review of the consequences of prenatal marijuana exposure: An emerging theme of a deficiency in aspects of executive function. Neurotoxicol. Teratol. 2001, 23, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Huizink, A.C. Prenatal cannabis exposure and infant outcomes: Overview of studies. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 52, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Jutras-Aswad, D.; DiNieri, J.A.; Harkany, T.; Hurd, Y.L. Neurobiological consequences of maternal cannabis on human fetal development and its neuropsychiatric outcome. Eur. Arch. Psychiatry Clin. Neurosci. 2009, 259, 395–412. [Google Scholar] [CrossRef] [PubMed]
- Trezza, V.; Cuomo, V.; Vanderschuren, L.J. Cannabis and the developing brain: Insights from behavior. Eur. J. Pharmacol. 2008, 585, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Campolongo, P.; Trezza, V.; Ratano, P.; Palmery, M.; Cuomo, V. Developmental consequences of perinatal cannabis exposure: Behavioral and neuroendocrine effects in adult rodents. Psychopharmacology 2011, 214, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.; Escuredo, L.; Munoz, R.; Rodriguez de Fonseca, F.; Navarro, M. Long-term behavioural and neuroendocrine effects of perinatal activation or blockade of CB cannabinoid receptors. Behav. Pharmacol. 2005, 16, 423–430. [Google Scholar] [CrossRef]
- Navarro, M.; Rubio, P.; Rodriguez de Fonseca, F. Sex-dimorphic psychomotor activation after perinatal exposure to (-)-delta-9-tetrahydrocannabinol. An ontogenic study in Wistar rats. Psychopharmacology 1994, 116, 414–422. [Google Scholar] [CrossRef]
- Navarro, M.; de Miguel, R.; Rodríguez de Fonseca, F.; Ramos, J.A.; Fernández-Ruiz, J.J. Perinatal cannabinoid exposure modifies the sociosexual approach behavior and the mesolimbic dopaminergic activity of adult male rats. Behav. Brain Res. 1996, 75, 91–98. [Google Scholar] [CrossRef]
- Rubio, P.; Rodriguez de Fonesca, F.; Munoz, R.M.; Ariznavarreta, C.; Martin-Calderon, J.L.; Navarro, M. Long-term behavioral effects of perinatal exposure to delta-9-tetrahydrocannabinol in rats: Possible role of pituitary-adrenal axis. Life Sci. 1995, 56, 2169–2176. [Google Scholar] [CrossRef]
- Vela, G.; Fuentes, J.A.; Bonnin, A.; Fernández-Ruiz, J.; Ruiz-Gayo, M. Perinatal exposure to delta 9-tetrahydrocannabinol (delta 9-THC) leads to changes in opioid-related behavioral patterns in rats. Brain Res. 1995, 680, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Vela, G.; Martín, S.; García-Gil, L.; Crespo, J.A.; Ruiz-Gayo, M.; Fernández-Ruiz, J.J.; García-Lecumberri, C.; Pélaprat, D.; Fuentes, J.A.; Ramos, J.A.; et al. Maternal exposure to delta9-tetrahydrocannabinol facilitates morphine self-administration behavior and changes regional binding to central mu opioid receptors in adult offspring female rats. Brain Res. 1998, 807, 101–109. [Google Scholar] [CrossRef]
- Coy, K.C.; Haight, S.C.; Anstey, E.; Grant, A.M.; Ruffo, N.; Ko, J.Y. Postpartum Marijuana Use, Perceptions of Safety, and Breastfeeding Initiation and Duration: An Analysis of PRAMS Data From Seven States, 2017. J. Hum. Lact. 2021, 37, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Moss, M.J.; Bushlin, I.; Kazmierczak, S.; Koop, D.; Hendrickson, R.G.; Zuckerman, K.E.; Grigsby, T.M. Cannabis use and measurement of cannabinoids in plasma and breast milk of breastfeeding mothers. Pediatr. Res. 2021, 90, 861–868. [Google Scholar] [CrossRef]
- Chadwick, V.L.; Rohleder, C.; Koethe, D.; Leweke, F.M. Cannabinoids and the endocannabinoid system in anxiety, depression, and dysregulation of emotion in humans. Curr. Opin. Psychiatry 2020, 33, 20–42. [Google Scholar] [CrossRef]
- Mammen, G.; Rueda, S.; Roerecke, M.; Bonato, S.; Lev-Ran, S.; Rehm, J. Association of Cannabis With Long-Term Clinical Symptoms in Anxiety and Mood Disorders: A Systematic Review of Prospective Studies. J. Clin. Psychiatry 2018, 79, 2248. [Google Scholar] [CrossRef]
- Ko, J.Y.; Tong, V.T.; Bombard, J.M.; Hayes, D.K.; Davy, J.; Perham-Hester, K.A. Marijuana use during and after pregnancy and association of prenatal use on birth outcomes: A population-based study. Drug Alcohol Depend. 2018, 187, 72–78. [Google Scholar] [CrossRef]
- Numan, M. Medial preoptic area and maternal behavior in the female rat. J. Comp. Physiol. Psychol. 1974, 87, 746–759. [Google Scholar] [CrossRef] [PubMed]
- Numan, M.; Rosenblatt, J.S.; Komisaruk, B.R. Medial preoptic area and onset of maternal behavior in the rat. J. Comp. Physiol. Psychol. 1977, 91, 146–164. [Google Scholar] [CrossRef]
- Numan, M. Motivational systems and the neural circuitry of maternal behavior in the rat. Dev. Psychobiol. 2007, 49, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Numan, M.; Stolzenberg, D.; Dellevigne, A.; Correnti, C.; Numan, M. Temporary inactivation of ventral tegmental area neurons with either muscimol or baclofen reversibly disrupts maternal behavior in rats through different underlying mechanisms. Behav. Neurosci. 2009, 123, 740–751. [Google Scholar] [CrossRef] [PubMed]
- Hoekzema, E.; Tamnes, C.K.; Berns, P.; Barba-Müller, E.; Pozzobon, C.; Picado, M.; Lucco, F.; Martínez-García, M.; Desco, M.; Ballesteros, A.; et al. Becoming a mother entails anatomical changes in the ventral striatum of the human brain that facilitate its responsiveness to offspring cues. Psychoneuroendocrinology 2020, 112, 104507. [Google Scholar] [CrossRef] [PubMed]
- Rilling, J.K. The neural and hormonal bases of human parental care. Neuropsychologia 2013, 51, 731–747. [Google Scholar] [CrossRef] [PubMed]
- Bosch, O. Maternal nurturing is dependent on her innate anxiety: The behavioral roles of brain oxytocin and vasopressin. Horm. Behav. 2011, 59, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Numan, M.; Stolzenberg, D.S. Medial preoptic area interactions with dopamine neural systems in the control of the onset and maintenance of maternal behavior in rats. Front. Neuroendocrinol. 2009, 30, 46–64. [Google Scholar] [CrossRef] [PubMed]
- Butovsky, E.; Juknat, A.; Elbaz JShabat-Simon, M.; Eilam, E.; Zangen, A.; Altstein, M.; Volgel, Z. Chronic exposure to delta9-tetrahydrocannabinol downregulates oxytocin and oxytocin-associated neurophysin in specific brain areas. Mol. Cell. Neurosci. 2006, 31, 795–801. [Google Scholar] [CrossRef] [PubMed]
- NOT-DA-21-049; Notice of Information: Establishment of a Standard THC Unit to Be used in Research. National Institute on Drug Abuse: North Bethesda, MD, USA, 2021.
- Castelli, V.; Lavanco, G.; Feo, S.; D’Amico, C.; Micale, V.; Kuchar, M.; Plescia, F.; Brancato, A.; Cannizzaro, C. Prenatal Exposure to Δ9-Tetrahydrocannabinol Affects Hippocampus-Related Cognitive Functions in the Adolescent Rat Offspring: Focus on Specific Markers of Neuroplasticity. Pharmaceutics 2023, 15, 692. [Google Scholar] [CrossRef] [PubMed]
- Drazanova, E.; Ruda-Kucerova, J.; Kratka, L.; Stark, T.; Kuchar, M.; Maryska, M.; Drago, F.; Starcuk, Z., Jr.; Micale, V. Different effects of prenatal MAM vs. perinatal THC exposure on regional cerebral blood perfusion detected by Arterial Spin Labelling MRI in rats. Sci. Rep. 2019, 9, 6062. [Google Scholar] [CrossRef]
- Navarro, M.; Rodríguez de Fonseca, F.; Hernández, M.L.; Ramos, J.A.; Fernández-Ruiz, J.J. Motor behavior and nigrostriatal dopaminergic activity in adult rats perinatally exposed to cannabinoids. Pharmacol. Biochem. Behav. 1994, 47, 47–58. [Google Scholar] [CrossRef]
- US Environmental Protection Agency. Health Effects Test Guidelines OPPTS 870.6300 Developmental Neurotoxicity Study. 1998. Available online: https://nepis.epa.gov/Exe/ZyPURL.cgi?Dockey=P100IRWO.TXT (accessed on 20 April 2024).
- Institute for Laboratory Animal Resources. Guide for the Care and Use of Laboratory Animals, 8th ed.; The National Academy Press: Washington, DC, USA, 2011. [Google Scholar]
- Nephew, B.C.; Bridges, R.S. Effects of chronic social stress during lactation on maternal behavior and growth in rats. Stress 2011, 14, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Abel, E.L. Suppression of pup retrieving behavior in rats following administration of delta-9-Tetrahydrocannabinol. Experientia 1972, 28, 1187–1188. [Google Scholar] [CrossRef] [PubMed]
- Abel, E.L.; Day, N.; Dintcheff, B.A.; Ernst, C.A.S. Inhibition of postnatal maternal performance in rats treated with marijuana extract during pregnancy. Bull. Psychon. Soc. 1979, 14, 353–354. [Google Scholar] [CrossRef]
- Abel, E.L.; Dintcheff, B.A.; Day, N. Effects of marijuana on pregnant rats and their offspring. Psychopharmacology 1980, 71, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Levay, E.A.; Paolini, A.G.; Govic, A.; Hazi, A.; Penman, J.; Kent, S. Anxiety-like behaviour in adult rats perinatally exposed to maternal calorie restriction. Behav. Brain Res. 2008, 191, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Maguire, J.; Mody, I. GABA(A)R plasticity during pregnancy: Relevance to postpartum depression. Neuron 2008, 59, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Numan, M.; Numan, M.J.; Pliakou, N.; Stolzenberg, D.S.; Mullins, O.J.; Murphy, J.M.; Smith, C.D. The effects of D1 or D2 dopamine receptor antagonism in the medial preoptic area, ventral pallidum, or nucleus accumbens on the maternal retrieval response and other aspects of maternal behavior in rats. Behav. Neurosci. 2005, 119, 1588–1604. [Google Scholar] [CrossRef] [PubMed]
- Tseng, A.H.; Harding, J.W.; Craft, R.M. Pharmacokinetic factors in sex differences in delta-9-tetrahydrocannabinol-induced behavioral effects in rats. Behav. Brain Res. 2004, 154, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Ströher, R.; de Oliveira, C.; Costa Lopes, B.; da Silva, L.S.; Regner, G.G.; Richardt Medeiros, H.; de Macedo, I.C.; Caumo, W.; Torres, I.L.S. Maternal deprivation alters nociceptive response in a gender-dependent manner in rats. Int. J. Dev. Neurosci. 2019, 76, 25–33. [Google Scholar] [CrossRef]
- Fragoso, J.; Lira, A.D.O.; Chagas, G.S.; Lucena Cavalcanti, C.C.; Beserra, R.; de Santana-Muniz, G.; Bento-Santos, A.; Martins, G.; Pirola, L.; da Silva Aragão, R.; et al. Maternal voluntary physical activity attenuates delayed neurodevelopment in malnourished rats. Exp. Physiol. 2017, 102, 1486–1499. [Google Scholar] [CrossRef] [PubMed]
- Bignall, K.E. Ontogeny of levels of neural organization: The righting reflex as a model. Exp. Neurol. 1974, 42, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Di Bartolomeo, M.; Stark, T.; Maurel, O.M.; Iannotti, F.A.; Kuchar, M.; Ruda-Kucerova, J.; Piscitelli, F.; Laudani, S.; Pekarik, V.; Salomone, S.; et al. Crosstalk between the transcriptional regulation of dopamine D2 and cannabinoid CB1 receptors in schizophrenia: Analyses in patients and in perinatal Δ9-tetrahydrocannabinol-exposed rats. Pharmacol. Res. 2021, 164, 105357. [Google Scholar] [CrossRef] [PubMed]
- Schou, J.; Prockop, L.D.; Dahlstrom, G.; Rohde, C. Penetration of delta-9-tetrahydrocannabinol and 11-OH-delta-9-tetrahydrocannabinol through the blood-brain barrier. Acta Pharmacol. Toxicol. 1977, 41, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, D.E.; Martin, B.R.; Gamagaris, Z.; Miller, N.; Fico, T. Plasma concentrations of delta-9-tetrahydrocannabinol in dams and fetuses following acute or multiple prenatal dosing in rats. Life Sci. 1989, 44, 697–701. [Google Scholar] [CrossRef] [PubMed]
- Foster, B.C.; Abramovici, H.; Harris, C.S. Cannabis and Cannabinoids: Kinetics and Interactions. Am. J. Med. 2019, 132, 1266–1270. [Google Scholar] [CrossRef] [PubMed]
- Nadulski, T.; Pragst, F.; Weinberg, G.; Roser, P.; Schnelle, M.; Fronk, E.-M.; Stadelmann, A.M. Randomized, double-blind, placebo-controlled study about the effects of cannabidiol (CBD) on the pharmacokinetics of Delta9-tetrahydrocannabinol (THC) after oral application of THC verses standardized cannabis extract. Ther. Drug Monit. 2005, 27, 799–810. [Google Scholar] [CrossRef] [PubMed]
- De Sesso, J.M.; Watson, R.E.; Keen, C.L.; Hazelden, K.P.; Haws, L.C.; Li, A.A. Analysis and integration of developmental neurotoxicity and ancillary data into risk assessment: A case study of dimethoate. J. Toxicol. Environ. Health Part A 2009, 72, 94–109. [Google Scholar] [CrossRef] [PubMed]
- Hindelang, P.; Scharinger, A.; Richling, E.; Walch, S.G.; Lachenmeier, D.W. Using the BMD Approach to Derive Acceptable Daily Intakes of Cannabidiol (CBD) and Tetrahydrocannabinol (THC) Relevant to Electronic Cigarette Liquids. Front. Biosci. 2022, 27, 228. [Google Scholar] [CrossRef] [PubMed]
- Lachenmeier, D.W.; Rehm, J. Comparative Risk Assessment of Alcohol, Tobacco, Cannabis and Other Illicit Drugs Using the Margin of Exposure Approach. Sci. Rep. 2015, 5, 8126. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carlson, E.; Teboul, E.; Canale, C.; Coleman, H.; Angeliu, C.; Garbarini, K.; Markowski, V.P. Perinatal Tetrahydrocannabinol Compromises Maternal Care and Increases Litter Attrition in the Long–Evans Rat. Toxics 2024, 12, 311. https://doi.org/10.3390/toxics12050311
Carlson E, Teboul E, Canale C, Coleman H, Angeliu C, Garbarini K, Markowski VP. Perinatal Tetrahydrocannabinol Compromises Maternal Care and Increases Litter Attrition in the Long–Evans Rat. Toxics. 2024; 12(5):311. https://doi.org/10.3390/toxics12050311
Chicago/Turabian StyleCarlson, Emma, Eric Teboul, Charlene Canale, Harper Coleman, Christina Angeliu, Karissa Garbarini, and Vincent P. Markowski. 2024. "Perinatal Tetrahydrocannabinol Compromises Maternal Care and Increases Litter Attrition in the Long–Evans Rat" Toxics 12, no. 5: 311. https://doi.org/10.3390/toxics12050311
APA StyleCarlson, E., Teboul, E., Canale, C., Coleman, H., Angeliu, C., Garbarini, K., & Markowski, V. P. (2024). Perinatal Tetrahydrocannabinol Compromises Maternal Care and Increases Litter Attrition in the Long–Evans Rat. Toxics, 12(5), 311. https://doi.org/10.3390/toxics12050311