

Analysis of Binding Modes between Three Perfluorosulfonates and GPER Based on Computational Simulation and Multiple Spectral Methods

Abstract
:1. Introduction
2. Experimental Materials and Methods
2.1. Materials and Reagents
2.2. In Silico Methods
2.3. Experimental Methods
3. Mechanism Discussion
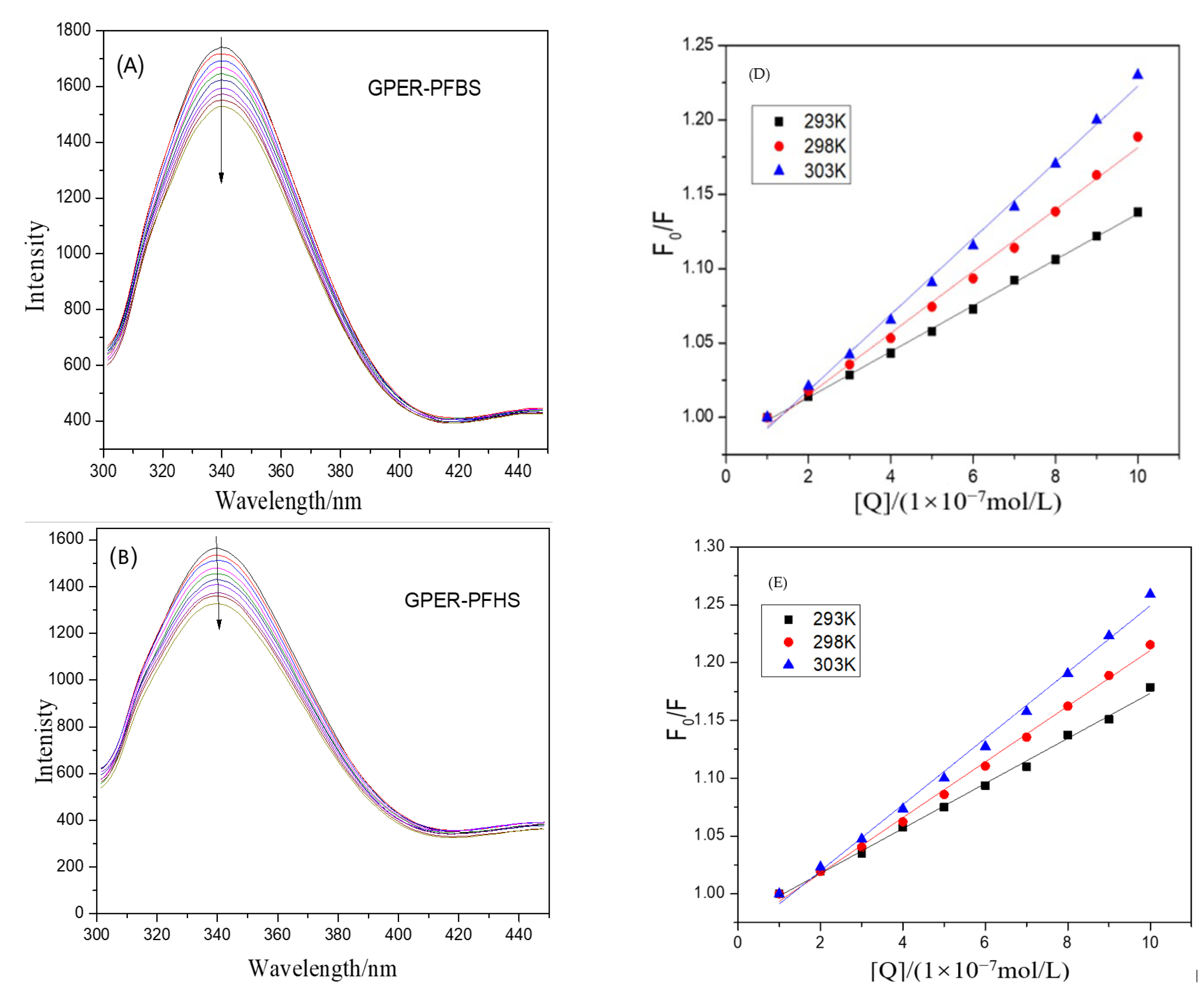
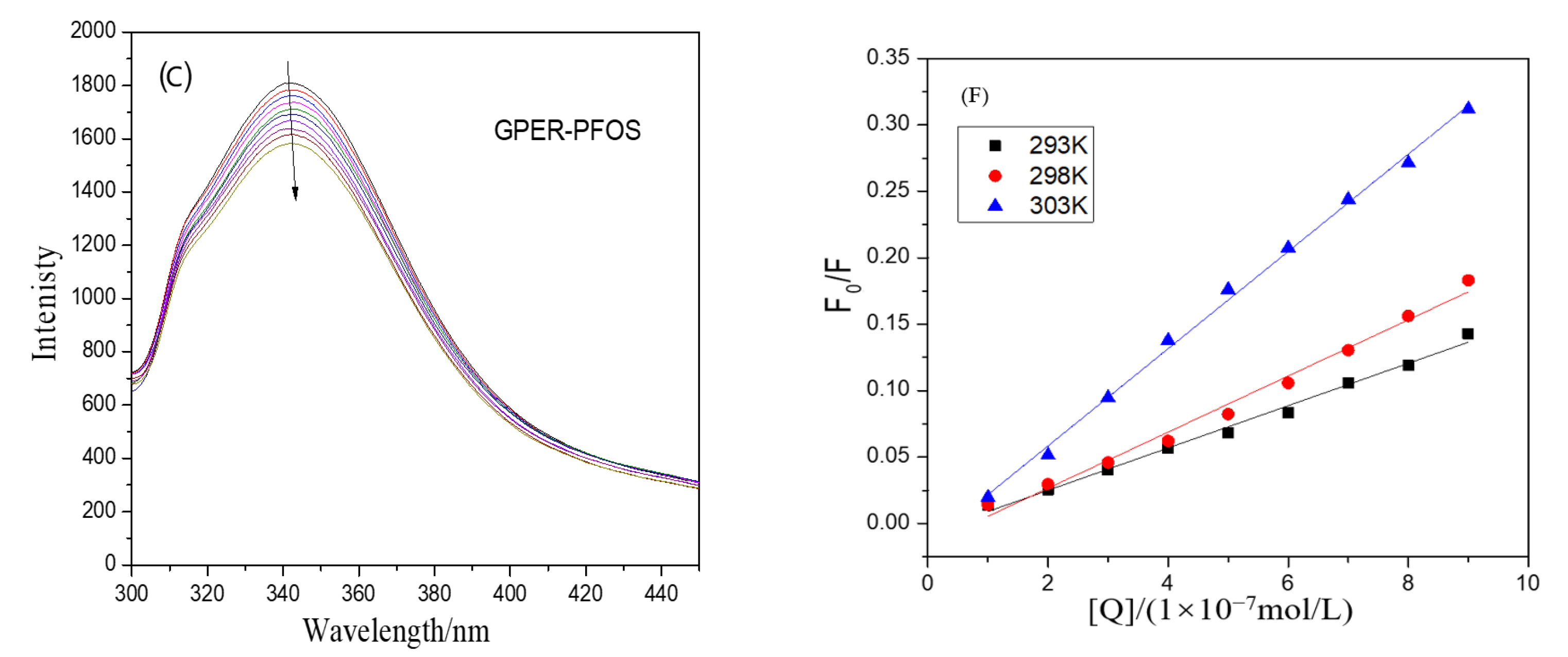
3.1. Unraveling the Dynamics and Binding Affinity in the Interaction between GPER and PFSAs by Steady-State Fluorescence Spectrum
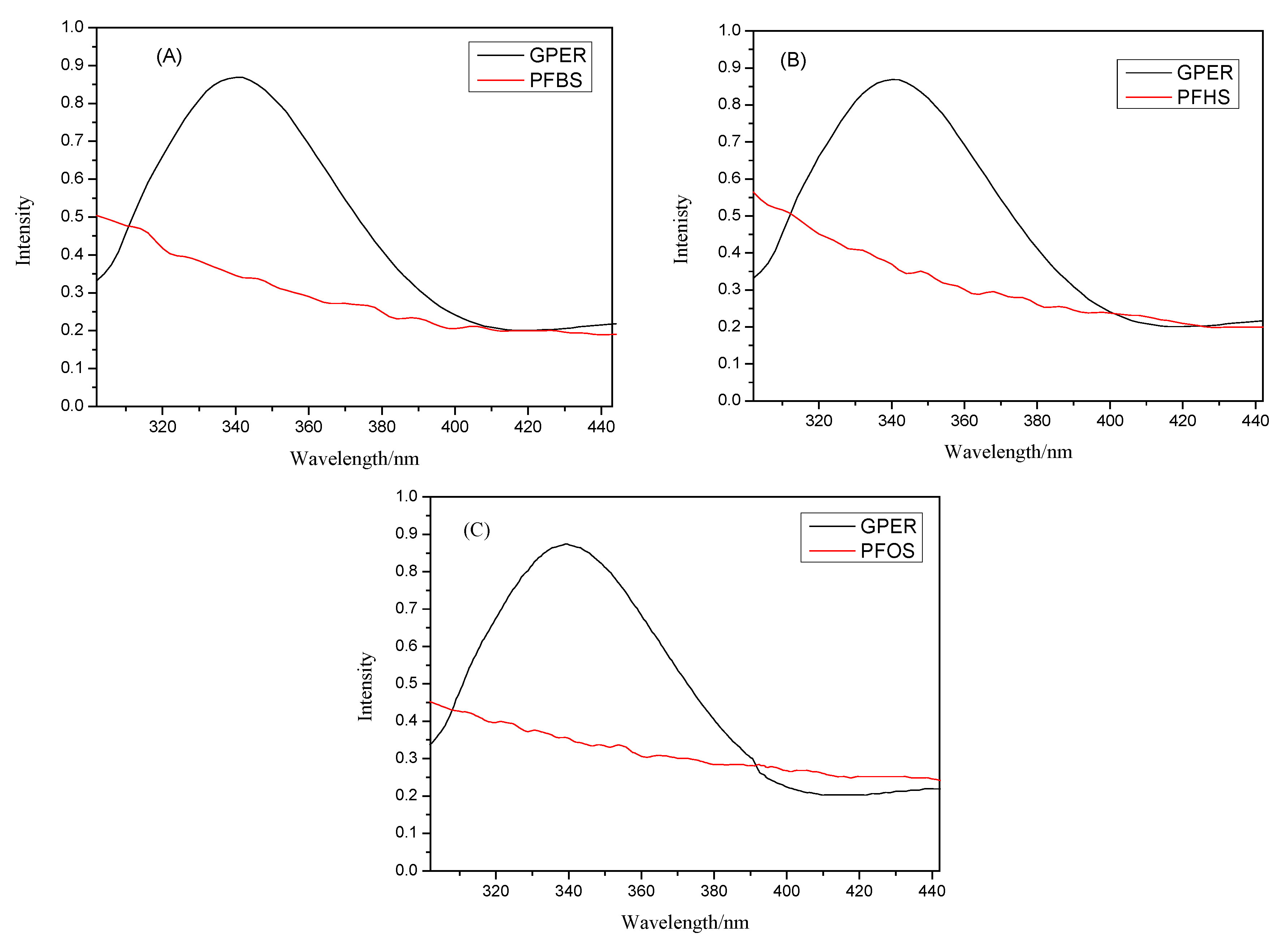
3.2. Mechanisms and Analysis of Fluorescence Quenching by PFSAs: Static Quenching and Non-Radiative Energy Transfer in GPER
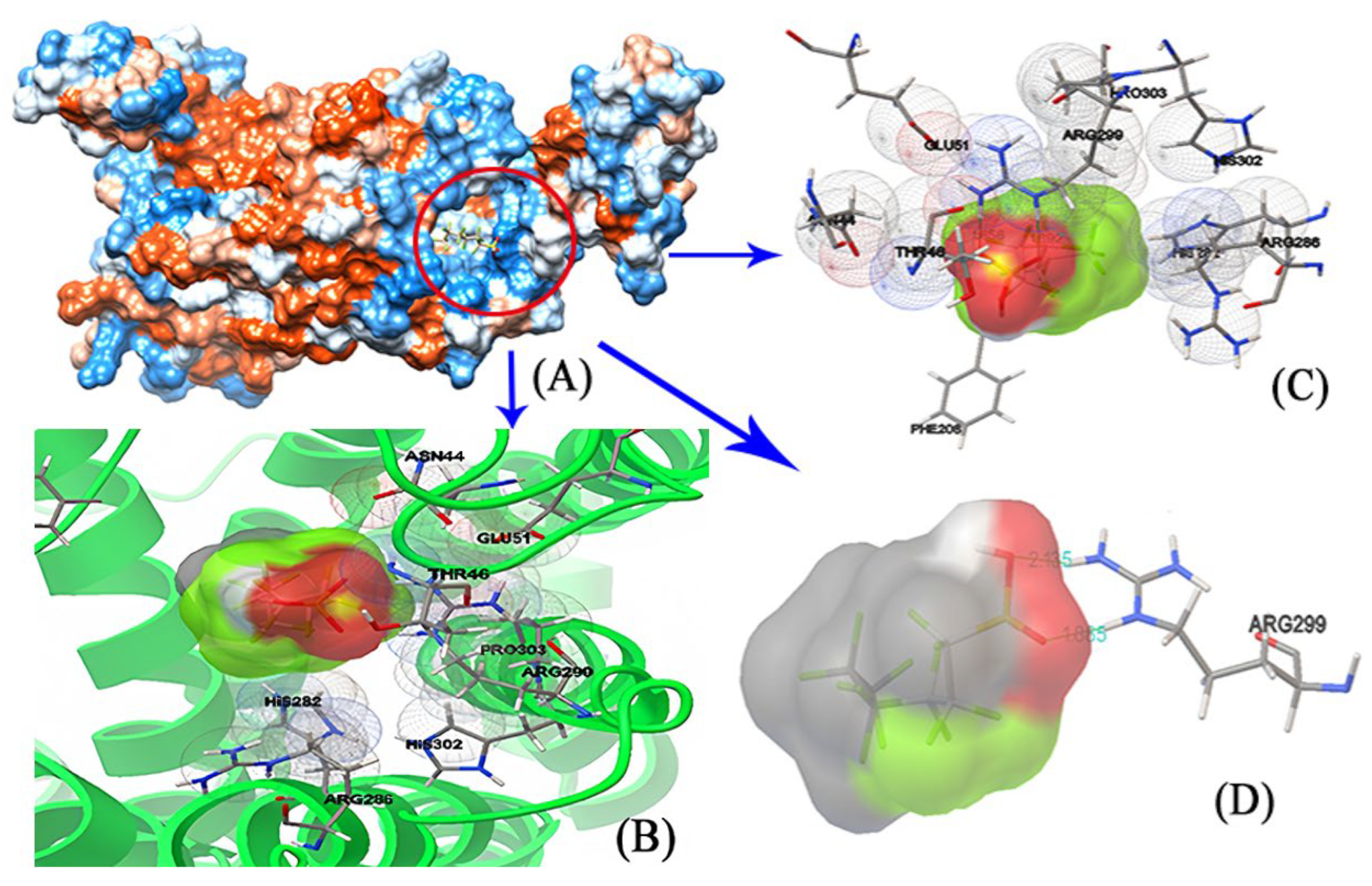
3.3. Molecular Docking: Visualizing the Interactions between PFSA Small Molecules and Large Proteins
4. Secondary Structural Changes
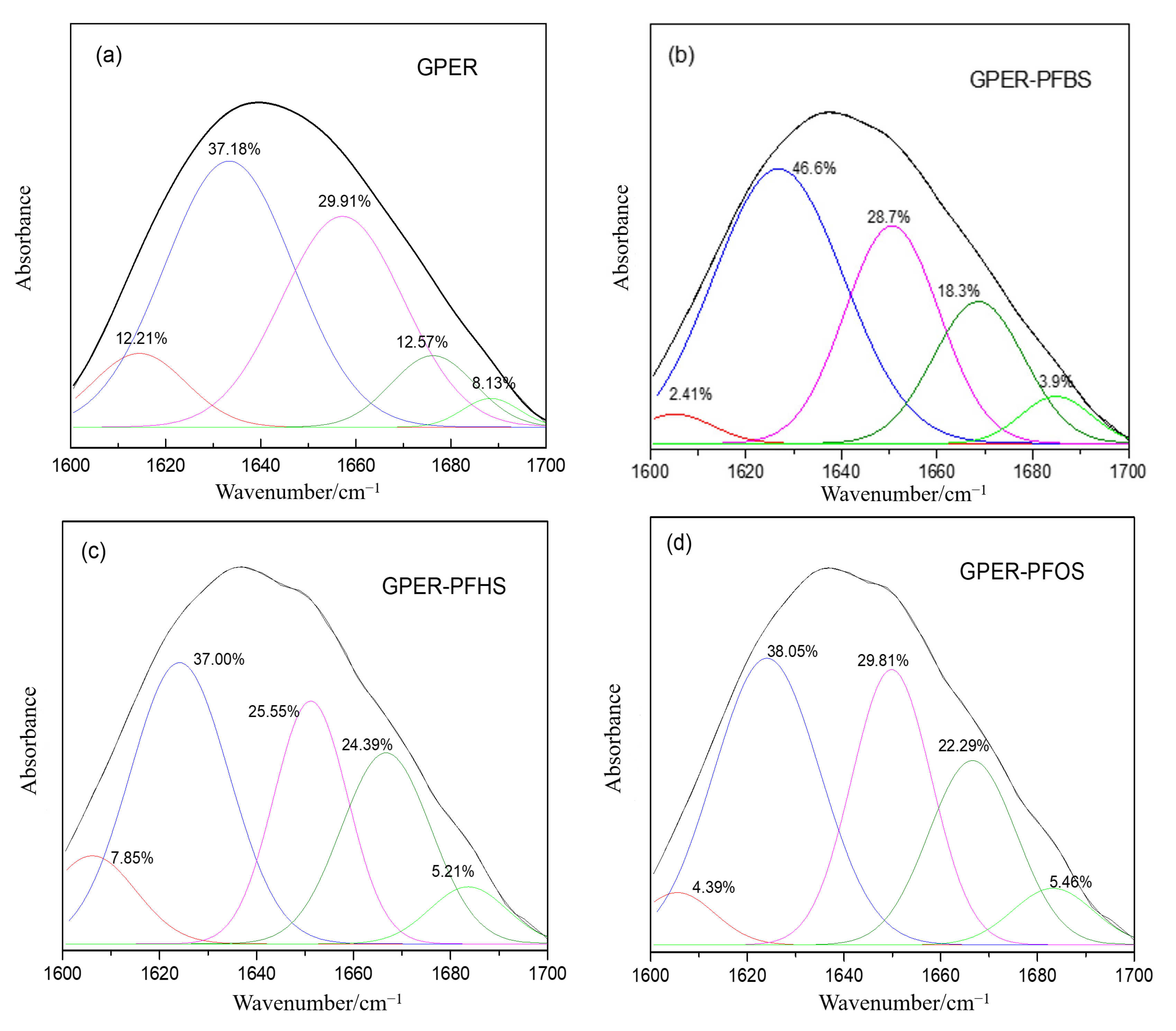
4.1. Impact of PFS Binding on Protein Secondary Structure: Insights from Fourier Transform Infrared (FTIR) Spectroscopy
4.2. Validation of Protein Conformational Changes upon Small Molecular Ligand Binding Using Three-Dimensional Fluorescence Spectrum
5. Molecular Dynamics Simulation and Thermodynamic Analysis
5.1. Structural Stability and Ligand Binding Dynamics of GPER and GPER–PFSA Complexes Investigated through Molecular Dynamic Simulation
5.2. Thermodynamic Analysis of Protein–Small Molecule Ligand Interactions: Significance of Hydrophobic Interactions in the Binding of PFSAs with GPER
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T/K | Ksv | Kq | Ka | n | |
|---|---|---|---|---|---|
| PFBS | 293 K | 1.47 × 105 | 1.47 × 1013 | 1.04 × 106 | 0.9825 |
| 298 K | 1.90 × 105 | 1.90 × 1013 | 1.23 × 106 | 0.9735 | |
| 303 K | 2.31 × 105 | 2.31 × 1013 | 1.40 × 106 | 0.9670 | |
| PFHS | 293 K | 1.89 × 105 | 1.89 × 1013 | 1.26 × 106 | 0.9786 |
| 298 K | 2.19 × 105 | 2.19 × 1013 | 1.32 × 106 | 0.9695 | |
| 303 K | 2.58 × 105 | 2.58 × 1013 | 1.42 × 106 | 0.9693 | |
| PFOS | 293 K | 1.41 × 105 | 1.41 × 1013 | 1.25 × 106 | 0.9961 |
| 298 K | 1.69 × 105 | 1.69 × 1013 | 1.67 × 106 | 0.9943 | |
| 303 K | 2.35 × 105 | 2.35 × 1013 | 2.46 × 106 | 0.9986 |
References
- Zhu, J. Synthesis Study of Perfluorinated Bifunctional Compounds with Distal Functional Groups. Ph.D. Thesis, Tianjin University, Tianjin, China, 2013. [Google Scholar]
- Wang, Y.; Zhang, P. Human Exposure Pathways and Pollution Control Technology of Perfluorooctanoic Acid (PFOA) and Perfluorooctane Sulfonate (PFOS) s. Prog. Chem. 2010, 22, 210–219. [Google Scholar]
- Bossi, R.; Riget, F.F.; Dietz, R.; Sonne, C.; Fauser, P.; Dam, M.; Vorkamp, K. Preliminary screening of perfluorooctane sulfonate (PFOS) and other fluorochemicals in fish, birds and marine mammals from Greenland and the Faroe Islands. Environ. Pollut. 2005, 136, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Hu, X. Research Progress on Environmental Pollution Behavior, Toxic Effects, and Mechanisms of PFOS/PFOA. Environ. Chem. 2007, 28, 2153–2162. [Google Scholar]
- Jin, Y.; Liu, X.; Qin, H.; Ma, Y.; Fan, Y.; Zhang, Y.; Saito, N.; Kazuaki, S.; Akio, K. PFOS Pollution in Tap Water and Various Water Bodies in Some Regions of China. Chin. Environ. Sci. 2004, 24, 166–169. [Google Scholar]
- Qing, Z.; Man, Z.; Yu, L.; Lian, H.; Ai, L. Research Trends on Determination and Treatment of Perfluoronic Compounds. Environ. Sci. 2008, 31, 44–52. [Google Scholar]
- Skutlarek, D.; Exner, M.; Färber, H. Perfluorinated surfactants in surface and drinking waters. Environ. Sci. Pollut. Res. Int. 2006, 13, 299–307. [Google Scholar] [PubMed]
- Hu, W.; Jones, P.D.; Celius, T.; Giesy, J.P. Identification of genes responsive to PFOS using gene expression profiling. Environ. Toxicol. Pharmacol. 2005, 19, 57–70. [Google Scholar] [CrossRef]
- Starkov, A.A.; Wallace, K.B. Structural determinants of fluorochemical-induced mitochondrial dysfunction. Toxicol. Sci. 2002, 66, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Gaixia, W. Gas Chromatography-Mass Spectrometry Detection of Perfluorooctane Sulfonamide in Textiles. Master’s Thesis, Donghua University, Shanghai, China, 2011. [Google Scholar]
- Shi, Y.; Pan, Y.; Wang, J.; Cai, Y. Perfluorinated Chemicals Related Environmental Problems. Prog. Chem. 2009, 21, 369–376. [Google Scholar]
- Xie, B.Y.; Lv, Q.Y.; Ning, C.C.; Yang, B.Y.; Shan, W.W.; Cheng, Y.L.; Gu, C.; Luo, X.Z.; Zhang, Z.B.; Chen, X.J.; et al. TET1-GPER-PI3K/AKT pathway is involved in insulin-driven endometrial cancer cell proliferation. Biochem. Biophys. Res. Commun. 2017, 482, 857–862. [Google Scholar] [CrossRef]
- Méndez-Luna, D.; Martínez-Archundia, M.; Maroun, R.C.; Ceballos-Reyes, G.; Fragoso-Vázquez, M.J.; González-Juárez, D.E.; Correa-Basurto, J. Deciphering the GPER/GPR30-agonist and antagonists interactions using molecular modeling studies, molecular dynamics, and docking simulations. J. Biomol. Struct. Dyn. 2015, 33, 2161–2172. [Google Scholar] [CrossRef] [PubMed]
- Rosano, C.; Ponassi, M.; Santolla, M.F.; Pisano, A.; Felli, L.; Vivacqua, A.; Maggiolini, M.; Lappano, R. Macromolecular Modelling and Docking Simulations for the Discovery of Selective GPER Ligands. AAPS J. 2016, 18, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Lappano, R.; Rosano, C.; FSantolla, M.; Pupo, M.; MDe Francesco, E.; De Marco, P.; Ponassi, M.; Spallarossa, A.; Ranise, A.; Maggiolini, M. Two novel GPER agonists induce gene expression changes and growth effects in cancer cells. Curr. Cancer Drug Targets 2012, 12, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Majumder, S.; Das, S.; Moulik, S.R.; Mallick, B.; Pal, P.; Mukherjee, D. G-protein coupled estrogen receptor (GPER) inhibits final oocyte maturation in common carp, Cyprinus carpio. Gen. Comp. Endocrinol. 2015, 211, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, Y. GPCRRD: G protein-coupled receptor spatial restraint database for 3-D structure modeling and function annotation. Bioinformatics 2010, 26, 3004–3005. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef]
- Mirsaleh, M.R.; Meybodi, M.R. Balancing exploration and exploitation in memetic algorithms: A learning automata approach. Comput. Intell. 2017, 34, 282–309. [Google Scholar] [CrossRef]
- Schüttelkopf, A.W.; van Aalten, D.M.F. PRODRG: A tool for high-throughput crystallography of protein-ligand complexes. Acta Crystallogr. 2004, D60, 1355–1363. [Google Scholar] [CrossRef]
- Das, S.; Ghosh, P.; Koley, S.; Roy, A.S. Binding of naringin and naringenin with hen egg white lysozyme: A spectroscopic investigation and molecular docking study. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2018, 192, 211–221. [Google Scholar] [CrossRef]
- Sun, Z.; Cai, C.; Guo, F.; Ye, C.; Luo, Y.; Ye, S.; Luo, J.; Zhu, F.; Jiang, C. Oxygen sensitive polymeric nanocapsules for optical dissolved oxygen sensors. Nanotechnology 2017, 29, 145704. [Google Scholar] [CrossRef]
- Kumar, A.; Dutta, R.K. Photoluminescence Quenching Based Visual and Spectroscopic Method for Mercury Sensing by Surface-Modified Cadmium Sulphide Quantum Dots. J. Nanosci. Nanotechnol. 2018, 18, 2686–2694. [Google Scholar] [CrossRef] [PubMed]
- Nouhi, A.; Hajjoul, H.; Redon, R.; Gagné, J.; Mounier, S. Time-resolved laser fluorescence spectroscopy of organic ligands by europium: Fluorescence quenching and lifetime properties. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2018, 193, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Sinisi, V.; Forzato, C.; Cefarin, N.; Navarini, L.; Berti, F. Interaction of chlorogenic acids and quinides from coffee with human serum albumin. Food Chem. 2015, 168, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.W.; Ying, Y.; Yang, X.L.; Zheng, Z.Q.; Shi, J.N.; Sun, Y.F.; Huang, P. A spectroscopic study on the interaction between the anticancer drug erlotinib and human serum albumin. J. Incl. Phenom. Macrocycl. Chem. 2014, 78, 405–413. [Google Scholar] [CrossRef]
- Song, Z.; Yuan, W.; Zhu, R.; Wang, S.; Zhang, C.; Yang, B. Study on the interaction between curcumin and CopC by spectroscopic and docking methods. Int. J. Biol. Macromol. 2017, 96, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Lu, X.; Wang, Y.; Brodelius, P.E. Comparison of the interaction between lactoferrin and isomeric drugs. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2017, 173, 593–607. [Google Scholar] [CrossRef] [PubMed]
- Faridbod, F.; Ganjali, M.R.; Larijani, B.; Riahi, S.; Saboury, A.A.; Hosseini, M.; Norouzi, P.; Pillip, C. Interaction study of pioglitazone with albumin by fluorescence spectroscopy and molecular docking. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2011, 78, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Dahl, S.; Kalita, M.M.; Fischer, W.B. Interaction of antivirals with a heptameric bundle model of the p7 protein of hepatitis C virus. Chem. Biol. Drug Des. 2017, 91, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Talebi, S.; Saeedinia, A.; Zeinoddini, M.; Ahmadpour, F.; Sadeghizadeh, M. In silico study of mutations on binding between Interferon alpha 2b and IFNAR1 receptor. Curr. Proteom. 2017, 15, 71–76. [Google Scholar] [CrossRef]
- Suresh, P.; Divya, N.; Nidhi, S.; Rajasekaran, R. Phenytoin-Bovine Serum Albumin interactions—Modeling plasma protein—Drug binding: A multi-spectroscopy and in silico-based correlation. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2018, 193, 523–527. [Google Scholar] [CrossRef]
- Yu, L.; Yuan, Y.; Rensing, C.; Zhou, S. Combined spectroelectrochemical and proteomic characterizations of bidirectional Alcaligenes faecalis-electrode electron transfer. Biosens. Bioelectron. 2018, 106, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Hou, W.; Yang, Y.; Zhao, Y. Simultaneous Determination of Protein and Fat in Milk Powder by FTIRS. Phys. Test. Chem. Anal. Part B Chem. Anal. 2009, 72, 66–69. [Google Scholar]
- Buzoglu, L.; Maltas, E.; Ersoz, M.; Yildiz, S. Synthesis and characterization of PS-b-PGMA diblock copolymer and its interaction with blood proteins and donepezil. React. Funct. Polym. 2014, 82, 25–32. [Google Scholar] [CrossRef]
- Makarska-Bialokoz, M. Interactions of hemin with bovine serum albumin and human hemoglobin: A fluorescence quenching study. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2018, 193, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Goffin, A.; Guérin, S.; Rocher, V.; Varrault, G. Towards a better control of the wastewater treatment process: Excitation-emission matrix fluorescence spectroscopy of dissolved organic matter as a predictive tool of soluble BOD 5, in influents of six Parisian wastewater treatment plants. Environ. Sci. Pollut. Res. 2018, 25, 8765–8776. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Xu, J.; Chen, G. Studying Protein Solution Conformation through Three-Dimensional Fluorescence Spectroscopy. Chin. Sci. J. Chem. 1997, 1, 16–22. [Google Scholar]
- Zhu, J.; Zhao, X.; Liu, L.; Song, M.; Wu, S. Quantitative relationships between intermolecular interaction and damping parameters of irganox-1035/NBR hybrids: A combination of experiments, molecular dynamics simulations, and linear regression analyses. J. Appl. Polym. Sci. 2018, 135, 46202. [Google Scholar] [CrossRef]
- Jadhav, A.; Dash, R.; Hirwani, R.; Abdin, M. Sequence and structure insights of kazal type thrombin inhibitor protein: Studied with phylogeny, homology modeling and dynamic MM/GBSA studies. Int. J. Biol. Macromol. 2018, 108, 1045–1052. [Google Scholar] [CrossRef] [PubMed]
- Ross, P.D.; Subramanian, S. Thermodynamics of protein association reactions: Forces contributing to stability. Biochemistry 1981, 20, 3096–3102. [Google Scholar] [CrossRef]
- Li, J.; Li, J.; Jiao, Y.; Dong, C. Spectroscopic analysis and molecular modeling on the interaction of jatrorrhizine with human serum albumin (HSA). Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2014, 118, 48–54. [Google Scholar] [CrossRef]
- Yousefi, R.; Mohammadi, R.; Taheri-Kafrani, A.; Shahsavani, M.B.; Aseman, M.D.; Nabavizadeh, S.M.; Rashidi, M.; Poursasan, N.; Moosavi-Movahedi, A.A. Study of the interaction between two newly synthesized cyclometallated platinum (II) complexes and human serum albumin: Spectroscopic characterization and docking simulatio. J. Lumin. 2015, 159, 139–146. [Google Scholar] [CrossRef]
- Li, X.L.; Hu, Y.J.; Wang, H.; Yu, B.Q.; Yue, H.L. Molecular spectroscopy evidence of berberine binding to DNA: Comparative binding and thermodynamic profile of intercalation. Biomacromolecules 2012, 13, 873–888. [Google Scholar] [CrossRef] [PubMed]
- Tu, B.; Wang, Y.; Mi, R.; Ouyang, Y.; Hu, Y.J. Evaluation of the interaction between naringenin and human serum albumin: Insights from fluorescence spectroscopy, electrochemical measurement and molecular docking. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2015, 149, 536–543. [Google Scholar] [CrossRef] [PubMed]













| T/K | ∆H/(kJ·mol) | ∆G/(kJ·mol) | ∆S/(J·mol·K−1) | |
|---|---|---|---|---|
| PFBS | 293 K | 21.95 | −33.75 | 190.11 |
| 298 K | −34.74 | |||
| 303 K | −35.65 | |||
| PFHS | 293 K | 8.83 | −34.22 | 146.89 |
| 298 K | −34.92 | |||
| 303 K | −35.69 | |||
| PFOA | 293 K | 49.99 | −34.20 | 287.18 |
| 298 K | −35.50 | |||
| 303 K | −37.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, W.; Chen, Y.; Wei, Y.; Song, Z.; Li, C.; Zheng, Y.; Yi, Z. Analysis of Binding Modes between Three Perfluorosulfonates and GPER Based on Computational Simulation and Multiple Spectral Methods. Toxics 2024, 12, 315. https://doi.org/10.3390/toxics12050315
Liang W, Chen Y, Wei Y, Song Z, Li C, Zheng Y, Yi Z. Analysis of Binding Modes between Three Perfluorosulfonates and GPER Based on Computational Simulation and Multiple Spectral Methods. Toxics. 2024; 12(5):315. https://doi.org/10.3390/toxics12050315
Chicago/Turabian StyleLiang, Wenhui, Yanting Chen, Yuchen Wei, Zeyu Song, Cancan Li, Yanhong Zheng, and Zhongsheng Yi. 2024. "Analysis of Binding Modes between Three Perfluorosulfonates and GPER Based on Computational Simulation and Multiple Spectral Methods" Toxics 12, no. 5: 315. https://doi.org/10.3390/toxics12050315
APA StyleLiang, W., Chen, Y., Wei, Y., Song, Z., Li, C., Zheng, Y., & Yi, Z. (2024). Analysis of Binding Modes between Three Perfluorosulfonates and GPER Based on Computational Simulation and Multiple Spectral Methods. Toxics, 12(5), 315. https://doi.org/10.3390/toxics12050315