Transcriptomic Analysis of Gonadal Adipose Tissue in Male Mice Exposed Perinatally to 2,2′,4,4′-Tetrabromodiphenyl Ether (BDE-47)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatment
2.2. RNA Extraction and Sequencing
2.3. RT-qPCR
2.4. Bioinformatic Analysis of RNA-seq Data
3. Results
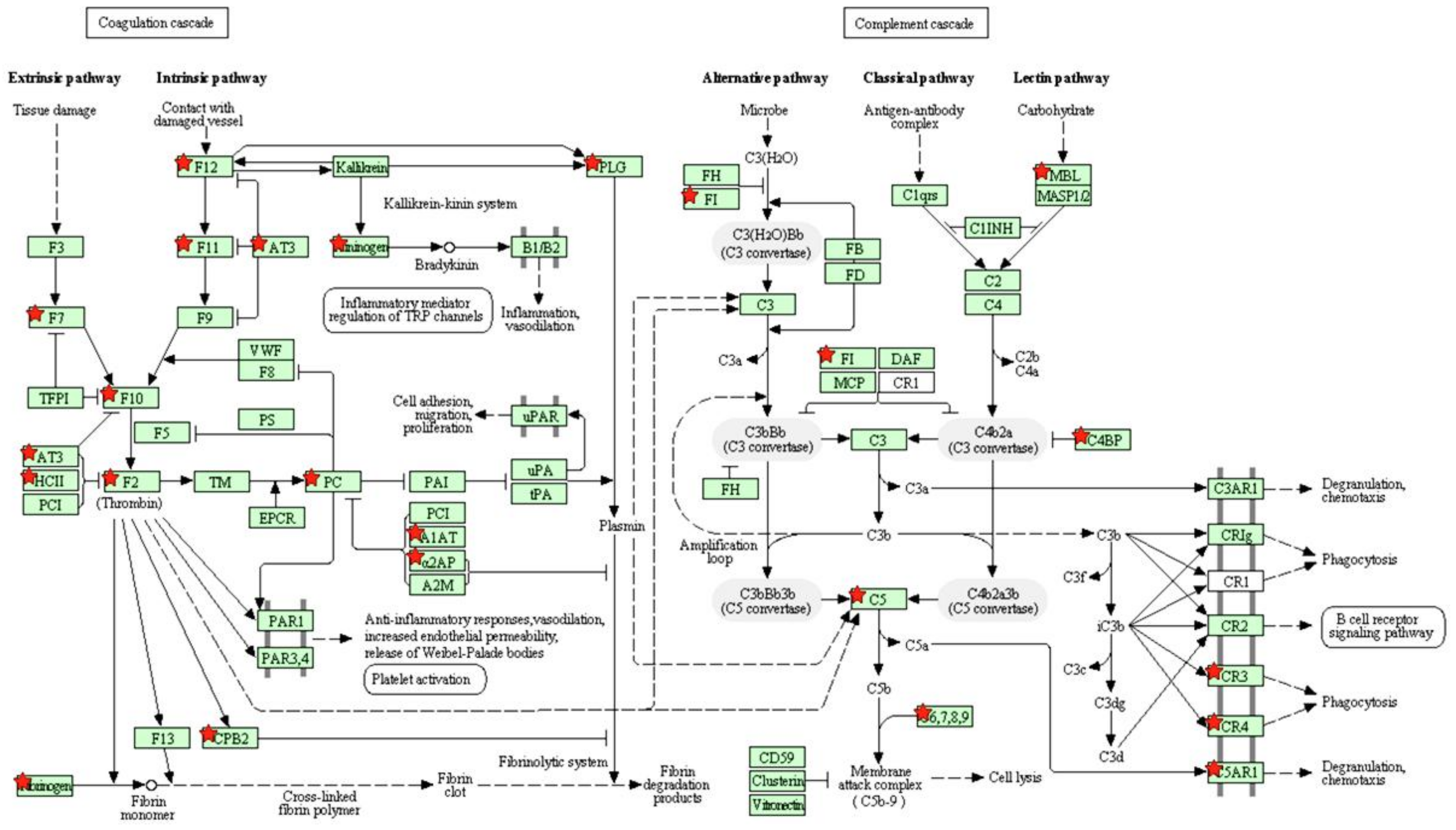
3.1. DAVID Analysis
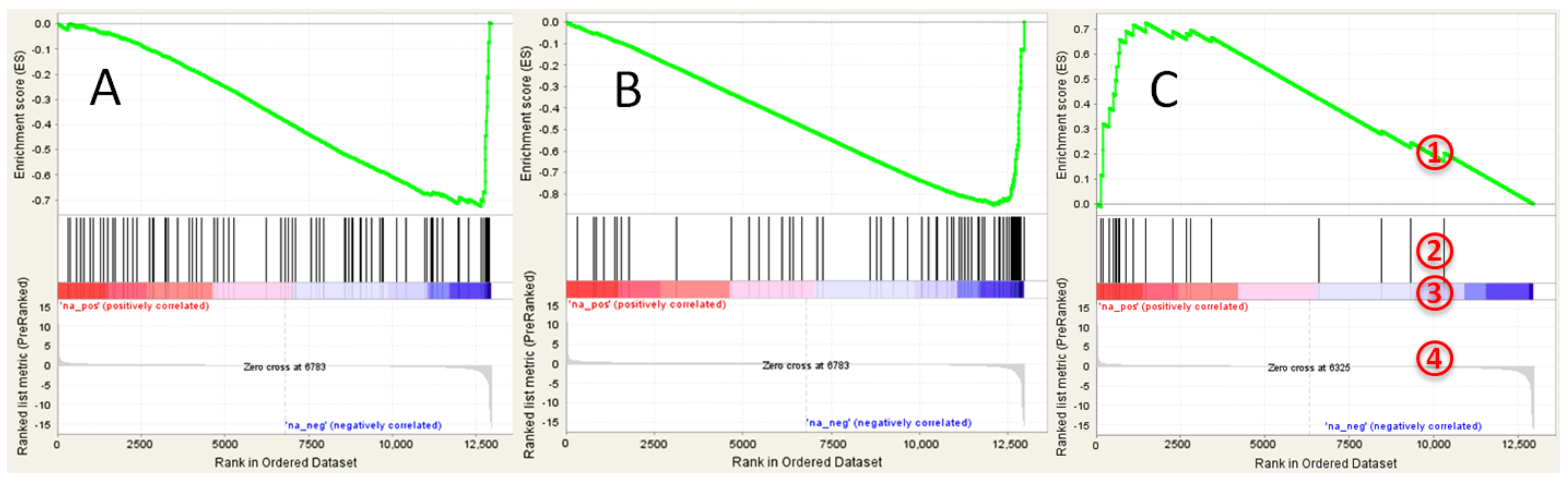
3.2. GSEA Analysis
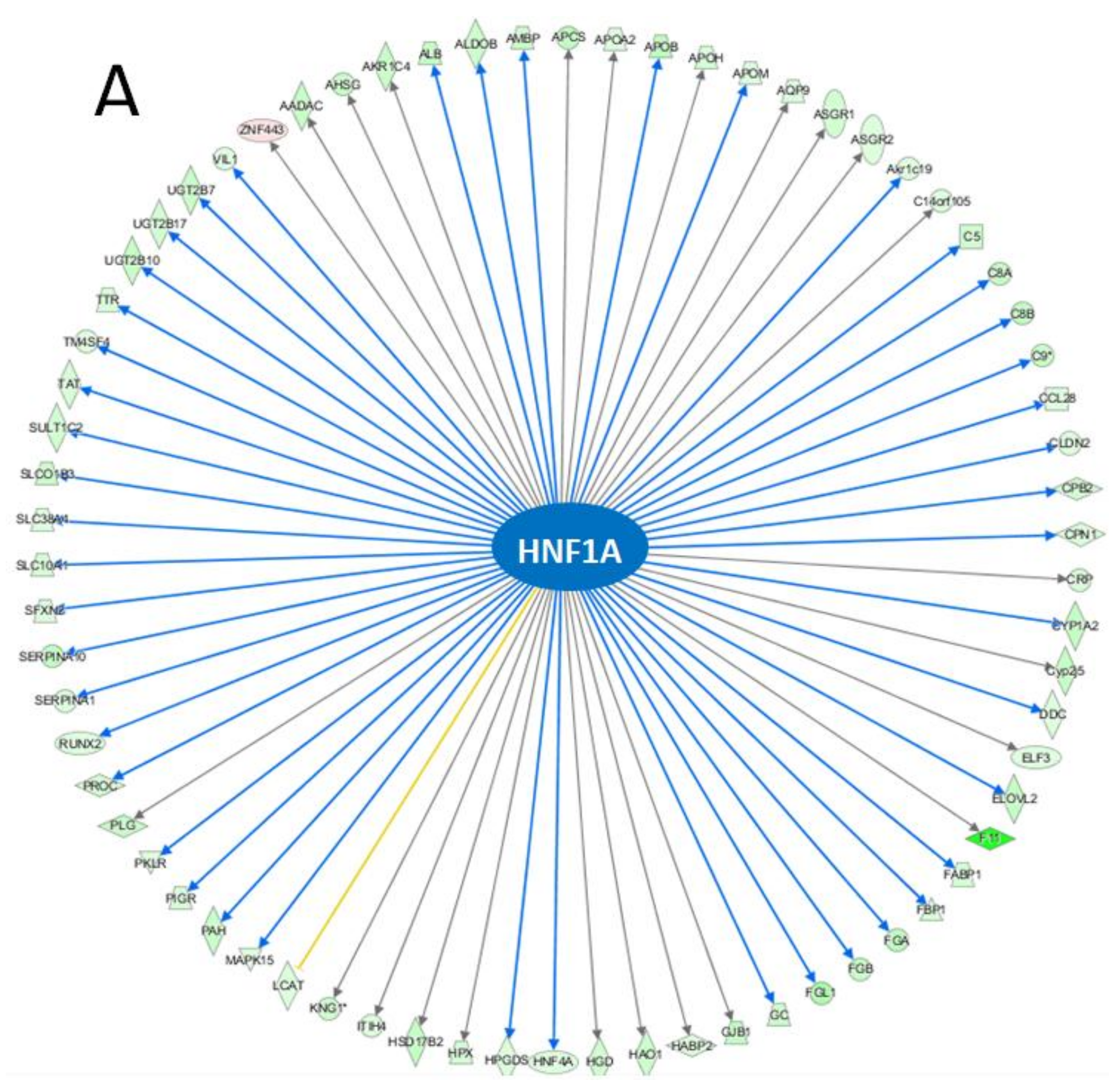
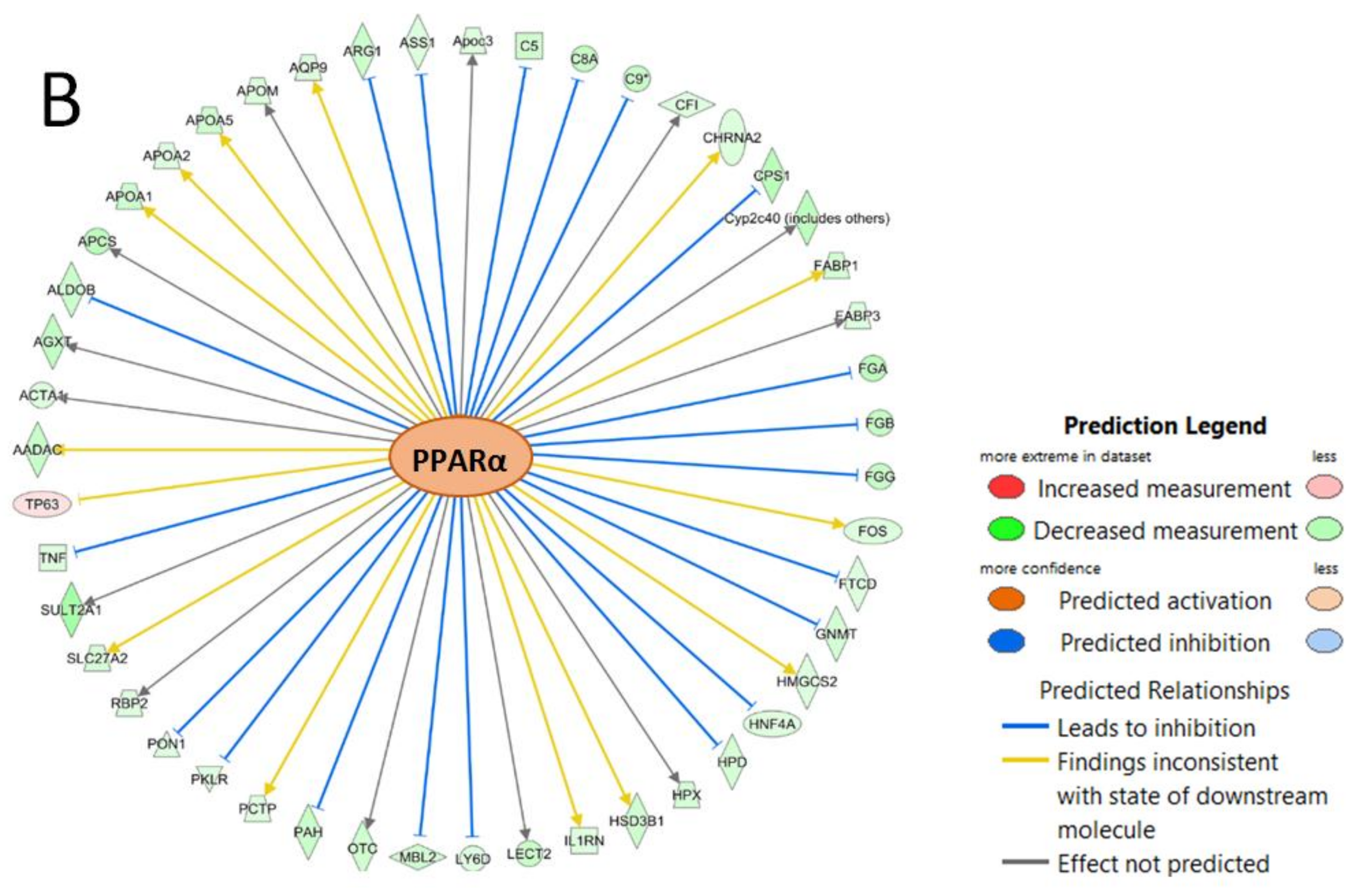
3.3. IPA Analysis
4. Discussion
4.1. Relevance of the Dosing Paradigm
4.2. Downregulation of the Complement and Coagulation Cascades Genes
4.3. Downregulation of Xenobiotic Metabolism Genes
4.4. Upregulation of De Novo Lipogenesis Genes
4.5. Network Affected by BDE-47
4.6. Role of PPARα in the Modulation of BDE-47 Induced Response
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gregg, E.W.; Zhuo, X.; Cheng, Y.J.; Albright, A.L.; Narayan, K.M.; Thompson, T.J. Trends in lifetime risk and years of life lost due to diabetes in the USA, 1985–2011: A modelling study. Lancet Diabetes Endocrinol. 2014, 2, 867–874. [Google Scholar] [CrossRef]
- National Institutes of Health (NIH). Funding Opportunity Announcement: Role of Environmental Chemical Exposures in the Development of Obesity, Type 2 Diabetes and Metabolic Syndrome (R21); National Institutes of Health: Bethesda, MD, USA, 2012.
- Heindel, J.J.; Vom Saal, F.S.; Blumberg, B.; Bovolin, P.; Calamandrei, G.; Ceresini, G.; Cohn, B.A.; Fabbri, E.; Gioiosa, L.; Kassotis, C.; et al. Parma consensus statement on metabolic disruptors. Environ. Health 2015, 14. [Google Scholar] [CrossRef] [PubMed]
- La Merrill, M.; Emond, C.; Kim, M.J.; Antignac, J.P.; Le Bizec, B.; Clément, K.; Birnbaum, L.S.; Barouki, R. Toxicological function of adipose tissue: Focus on persistent organic pollutants. Environ. Health Perspect. 2013, 121, 162–169. [Google Scholar] [PubMed]
- Trayhurn, P. Endocrine and signalling role of adipose tissue: New perspectives on fat. Acta Physiol. Scand. 2005, 184, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Kelishadi, R.; Poursafa, P.; Jamshidi, F. Role of environmental chemicals in obesity: A systematic review on the current evidence. J. Environ. Public Health 2013, 2013, 896789. [Google Scholar] [CrossRef] [PubMed]
- Regnier, S.M.; Sargis, R.M. Adipocytes under assault: Environmental disruption of adipose physiology. Biochim. Biophys. Acta 2014, 1842, 520–533. [Google Scholar] [CrossRef] [PubMed]
- Suvorov, A.; Takser, L. Facing the challenge of data transfer from animal models to humans: The case of persistent organohalogens. Environ. Health 2008. [Google Scholar] [CrossRef] [PubMed]
- Hurley, S.; Goldberg, D.; Nelson, D.O.; Guo, W.; Wang, Y.; Baek, H.G.; Park, J.S.; Petreas, M.; Bernstein, L.; Anton-Culver, H.; et al. Temporal Evaluation of Polybrominated Diphenyl Ether (PBDE) Serum Levels in Middle-Aged and Older California Women, 2011–2015. Environ. Sci. Technol. 2017, 51, 4697–4704. [Google Scholar] [CrossRef] [PubMed]
- Herbstman, J.B.; Sjödin, A.; Apelberg, B.J.; Witter, F.R.; Patterson, D.G., Jr.; Halden, R.U.; Jones, R.S.; Park, A.; Zhang, Y.; Heidler, J.; et al. Determinants of prenatal exposure to polychlorinated biphenyls (PCBs) and polybrominated diphenyl ethers (PBDEs) in an urban population. Environ. Health Perspect. 2007, 115, 1794–1800. [Google Scholar] [CrossRef] [PubMed]
- Doucet, J.; Tague, B.; Arnold, D.L.; Cooke, G.M.; Hayward, S.; Goodyer, C.G. Persistent organic pollutant residues in human fetal liver and placenta from Greater Montreal, Quebec: A longitudinal study from 1998 through 2006. Environ. Health Perspect. 2009, 117, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Guvenius, D.M.; Aronsson, A.; Ekman-Ordeberg, G.; Bergman, A.; Noren, K. Human prenatal and postnatal exposure to polybrominated diphenyl ethers, polychlorinated biphenyls, polychlorobiphenylols, and pentachlorophenol. Environ. Health Perspect. 2003, 111, 1235–1241. [Google Scholar] [CrossRef] [PubMed]
- Marchitti, S.A.; Lakind, J.S.; Naiman, D.Q.; Berlin, C.M.; Kenneke, J.F. Improving infant exposure and health risk estimates: Using serum data to predict polybrominated diphenyl ether concentrations in breast milk. Environ. Sci. Technol. 2013, 47, 4787–4795. [Google Scholar] [CrossRef] [PubMed]
- Schecter, A.; Pavuk, M.; Papke, O.; Ryan, J.J.; Birnbaum, L.; Rosen, R. Polybrominated diphenyl ethers (PBDEs) in U.S. mothers’ milk. Environ. Health Perspect. 2003, 111, 1723–1729. [Google Scholar] [CrossRef] [PubMed]
- Horton, M.K.; Bousleiman, S.; Jones, R.; Sjodin, A.; Liu, X.; Whyatt, R.; Wapner, R.; Factor-Litvak, P. Predictors of serum concentrations of polybrominated flame retardants among healthy pregnant women in an urban environment: A cross-sectional study. Environ. Health 2013, 12. [Google Scholar] [CrossRef] [PubMed]
- Geyer, H.J.; Schramm, K.W.; Darnerud, P.O.; Aune, M.; Feicht, E.A.; Fried, K.W.; Henkelmann, B.; Lenoir, D.; Schmid, P.; McDonald, T.A. Terminal Elimination Half-lives of the Brominated Flame Retardants TBBPA, HBCD, and Lower Brominated PBDEs in Humans. Organohalogen Compd. 2004, 66, 3820–3825. [Google Scholar]
- Renwick, A.G.; Walton, K. The use of surrogate endpoints to assess potential toxicity in humans. Toxicol. Lett. 2001, 120, 97–110. [Google Scholar] [CrossRef]
- Wade, M.G.; Parent, S.; Finnson, K.W.; Foster, W.; Younglai, E.; McMahon, A.; Cyr, D.G.; Hughes, C. Thyroid toxicity due to subchronic exposure to a complex mixture of 16 organochlorines, lead, and cadmium. Toxicol. Sci. 2002, 67, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Marsh, G.; Bergman, A.; Bladh, L.; Gillner, M.; Jakobsson, E. Synthesis of p-hydrobromobiphenyl ethers and binding to the thyroid hormone receptor. Organohalogen Compd. 1998, 37, 305–308. [Google Scholar]
- Vandenberg, L.N.; Welshons, W.V.; Vom Saal, F.S.; Toutain, P.L.; Myers, J.P. Should oral gavage be abandoned in toxicity testing of endocrine disruptors? Environ. Health 2014, 13. [Google Scholar] [CrossRef] [PubMed]
- Suvorov, A.; Vandenberg, L.N. To Cull or not to Cull? Considerations for studies of endocrine disrupting chemicals. Endocrinology 2016. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Hendrickson, D.G.; Sauvageau, M.; Goff, L.; Rinn, J.L.; Pachter, L. Differential analysis of gene regulation at transcript resolution with RNA-seq. Nat. Biotechnol. 2013, 31, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Rakhshandehroo, M.; Knoch, B.; Muller, M.; Kersten, S. Peroxisome proliferator-activated receptor alpha target genes. PPAR Res. 2010, 2010. [Google Scholar] [CrossRef] [PubMed]
- Suvorov, A.; Girard, S.; Lachapelle, S.; Abdelouahab, N.; Sebire, G.; Takser, L. Perinatal exposure to low-dose BDE-47, an emergent environmental contaminant, causes hyperactivity in rat offspring. Neonatology 2009, 95, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Restrepo, B.; Kannan, K.; Rapaport, D.P.; Rodan, B.D. Polybrominated diphenyl ethers and polychlorinated biphenyls in human adipose tissue from New York. Environ. Sci. Technol. 2005, 39, 5177–5182. [Google Scholar] [CrossRef] [PubMed]
- Sanders, J.M.; Chen, L.J.; Lebetkin, E.H.; Burka, L.T. Metabolism and disposition of 2,2′,4,4′-tetrabromodiphenyl ether following administration of single or multiple doses to rats and mice. Xenobiotica 2006, 36, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Staskal, D.F.; Diliberto, J.J.; DeVito, M.J.; Birnbaum, L.S. Toxicokinetics of BDE 47 in female mice: Effect of dose, route of exposure, and time. Toxicol. Sci. 2005, 83, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Antignac, J.P.; Cariou, R.; Maume, D.; Marchand, P.; Monteau, F.; Zalko, D.; Berrebi, A.; Cravedi, J.P.; Andre, F.; Le Bizec, B. Exposure assessment of fetus and newborn to brominated flame retardants in France: Preliminary data. Mol. Nutr. Food Res. 2008, 52, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Schecter, A.; Päpke, O.; Harris, T.R.; Tung, K.C.; Musumba, A.; Olson, J.; Birnbaum, L. Polybrominated diphenyl ether (PBDE) levels in an expanded market basket survey of U.S. food and estimated PBDE dietary intake by age and sex. Environ. Health Perspect. 2006, 114, 1515–1520. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Jiao, Y.; Hu, Y.; Sun, Z.; Zhou, X.; Feng, J.; Li, J.; Wu, Y. Levels of tetrabromobisphenol A, hexabromocyclododecanes and polybrominated diphenyl ethers in human milk from the general population in Beijing, China. Sci. Total Environ. 2013, 452–453, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Wilford, B.H.; Shoeib, M.; Harner, T.; Zhu, J.; Jones, K.C. Polybrominated diphenyl ethers in indoor dust in Ottawa, Canada: Implications for sources and exposure. Environ. Sci. Technol. 2005, 39, 7027–7035. [Google Scholar] [CrossRef] [PubMed]
- Hertle, E.; Stehouwer, C.D.; van Greevenbroek, M.M. The complement system in human cardiometabolic disease. Mol. Immunol. 2014, 61, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Samad, F.; Ruf, W. Inflammation, obesity, and thrombosis. Blood 2013, 122, 3415–3422. [Google Scholar] [CrossRef] [PubMed]
- Samad, F.; Pandey, M.; Loskutoff, D.J. Regulation of tissue factor gene expression in obesity. Blood 2001, 98, 3353–3358. [Google Scholar] [CrossRef] [PubMed]
- Suvorov, A.; Takser, L. Global Gene Expression Analysis in the Livers of Rat Offspring Perinatally Exposed to Low Doses of 2,2′,4,4′-tetrabromodiphenyl ether. Environ. Health Perspect. 2010, 118, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Sanders, J.M.; Burka, L.T.; Smith, C.S.; Black, W.; James, R.; Cunningham, M.L. Differential expression of CYP1A, 2B, and 3A genes in the F344 rat following exposure to a polybrominated diphenyl ether mixture or individual components. Toxicol. Sci. 2005, 88, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Wahl, M.; Lahni, B.; Guenther, R.; Kuch, B.; Yang, L.; Straehle, U.; Strack, S.; Weiss, C. A technical mixture of 2,2′,4,4′-tetrabromo diphenyl ether (BDE47) and brominated furans triggers aryl hydrocarbon receptor (AhR) mediated gene expression and toxicity. Chemosphere 2008, 73, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Pacyniak, E.K.; Cheng, X.; Cunningham, M.L.; Crofton, K.; Klaassen, C.D.; Guo, G.L. The flame retardants, polybrominated diphenyl ethers, are pregnane X receptor activators. Toxicol. Sci. 2007, 97, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Dunnick, J.K.; Brix, A.; Cunny, H.; Vallant, M.; Shockley, K.R. Characterization of polybrominated diphenyl ether toxicity in Wistar Han rats and use of liver microarray data for predicting disease susceptibilities. Toxicol. Pathol. 2012, 40, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.; Parker, M.; Mpanga, R.; Cevik, E.C.; Thorburn, C.; Suvorov, A. Developmental Exposure to 2,2′,4,4′–Tetrabromodiphenyl Ether Induces Long-Lasting Changes in Liver Metabolism in Male Mice. J. Endocr. Soc. 2017, 1, 323–344. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Lin, Y.S.; Zheng, X.E.; Senn, T.; Hashizume, T.; Scian, M.; Dickmann, L.J.; Nelson, S.D.; Baillie, T.A.; Hebert, M.F.; et al. An inducible cytochrome P450 3A4-dependent vitamin D catabolic pathway. Mol. Pharmacol. 2012, 81, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Basoli, V.; Santaniello, S.; Cruciani, S.; Ginesu, G.C.; Cossu, M.L.; Delitala, A.P.; Serra, P.A.; Ventura, C.; Maioli, M. Melatonin and Vitamin D Interfere with the Adipogenic Fate of Adipose-Derived Stem Cells. Int. J. Mol. Sci. 2017, 18, 981. [Google Scholar] [CrossRef] [PubMed]
- Mashek, D.G. Hepatic fatty acid trafficking: Multiple forks in the road. Adv. Nutr. 2013, 4, 697–710. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.; Leray, V.; Diez, M.; Serisier, S.; Bloc’h, J.L.; Siliart, B.; Dumon, H. Liver lipid metabolism. J. Anim. Physiol. Anim. Nutr. 2008, 92, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Daniels, J.L.; Pan, I.J.; Jones, R.; Anderson, S.; Patterson, D.G., Jr.; Needham, L.L.; Sjödin, A. Individual characteristics associated with PBDE levels in U.S. human milk samples. Environ. Health Perspect. 2010, 118, 155–160. [Google Scholar] [PubMed]
- Croes, K.; Colles, A.; Koppen, G.; Govarts, E.; Bruckers, L.; Van de Mieroop, E.; Nelen, V.; Covaci, A.; Dirtu, A.C.; Thomsen, C.; et al. Persistent organic pollutants (POPs) in human milk: A biomonitoring study in rural areas of Flanders (Belgium). Chemosphere 2012, 89, 988–994. [Google Scholar] [CrossRef] [PubMed]
- Frederiksen, M.; Thomsen, M.; Vorkamp, K.; Knudsen, L.E. Patterns and concentration levels of polybrominated diphenyl ethers (PBDEs) in placental tissue of women in Denmark. Chemosphere 2009, 76, 1464–1469. [Google Scholar] [CrossRef] [PubMed]
- Gee, J.R.; Moser, V.C. Acute postnatal exposure to brominated diphenylether 47 delays neuromotor ontogeny and alters motor activity in mice. Neurotoxicol. Teratol. 2008, 30, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Bondy, G.S.; Lefebvre, D.E.; Aziz, S.; Cherry, W.; Coady, L.; MacLellan, E.; Armstrong, C.; Barker, M.; Cooke, G.; Gaertner, D.; et al. Toxicologic and immunologic effects of perinatal exposure to the brominated diphenyl ether (BDE) mixture DE-71 in the Sprague-Dawley rat. Environ. Toxicol. 2013, 28, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Suvorov, A.; Battista, M.C.; Takser, L. Perinatal exposure to low-dose 2,2′,4,4′-tetrabromodiphenyl ether affects growth in rat offspring: What is the role of IGF-1? Toxicology 2009, 260, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Fuller, G.M.; Zhang, Z. Transcriptional control mechanism of fibrinogen gene expression. Ann. N. Y. Acad. Sci. 2001, 936, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Friedman, R.D.; Fournier, R.E. The locus control region activates serpin gene expression through recruitment of liver-specific transcription factors and RNA polymerase II. Mol. Cell. Biol. 2007, 27, 5286–5295. [Google Scholar] [CrossRef] [PubMed]
- Golledge, J.; Mangan, S.; Clancy, P. Effects of peroxisome proliferator-activated receptor ligands in modulating tissue factor and tissue factor pathway inhibitor in acutely symptomatic carotid atheromas. Stroke 2007, 38, 1501–1508. [Google Scholar] [CrossRef] [PubMed]
- Kockx, M.; Gervois, P.P.; Poulain, P.; Derudas, B.; Peters, J.M.; Gonzalez, F.J.; Princen, H.M.; Kooistra, T.; Staels, B. Fibrates suppress fibrinogen gene expression in rodents via activation of the peroxisome proliferator-activated receptor-alpha. Blood 1999, 93, 2991–2998. [Google Scholar] [PubMed]
- Cariello, N.F.; Romach, E.H.; Colton, H.M.; Ni, H.; Yoon, L.; Falls, J.G.; Casey, W.; Creech, D.; Anderson, S.P.; Benavides, G.R.; et al. Gene expression profiling of the PPAR-alpha agonist ciprofibrate in the cynomolgus monkey liver. Toxicol. Sci. 2005, 88, 250–264. [Google Scholar] [CrossRef] [PubMed]
- Mogilenko, D.A.; Dizhe, E.B.; Shavva, V.S.; Lapikov, I.A.; Orlov, S.V.; Perevozchikov, A.P. Role of the nuclear receptors HNF4 alpha, PPAR alpha, and LXRs in the TNF alpha-mediated inhibition of human apolipoprotein A-I gene expression in HepG2 cells. Biochemistry 2009, 48, 11950–11960. [Google Scholar] [CrossRef] [PubMed]
- Malik, S. Transcriptional regulation of the apolipoprotein AI gene. Front. Biosci. 2003, 8, d360–d368. [Google Scholar] [CrossRef] [PubMed]
- Berthou, L.; Duverger, N.; Emmanuel, F.; Langouët, S.; Auwerx, J.; Guillouzo, A.; Fruchart, J.C.; Rubin, E.; Denèfle, P.; Staels, B.; et al. Opposite regulation of human versus mouse apolipoprotein A-I by fibrates in human apolipoprotein A-I transgenic mice. J. Clin. Investig. 1996, 97, 2408–2416. [Google Scholar] [CrossRef] [PubMed]
- Prieur, X.; Lesnik, P.; Moreau, M.; Rodríguez, J.C.; Doucet, C.; Chapman, M.J.; Huby, T. Differential regulation of the human versus the mouse apolipoprotein AV gene by PPARalpha. Implications for the study of pharmaceutical modifiers of hypertriglyceridemia in mice. Biochim. Biophys. Acta 2009, 1791, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, M.; Lefebvre, P.; Staels, B. Molecular mechanism of PPARalpha action and its impact on lipid metabolism, inflammation and fibrosis in non-alcoholic fatty liver disease. J. Hepatol. 2015, 62, 720–733. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.M.; Remaley, A.T. High density lipoproteins are modulators of protease activity: Implications in inflammation, complement activation, and atherothrombosis. Atherosclerosis 2017, 259, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Pineda Torra, I.; Jamshidi, Y.; Flavell, D.M.; Fruchart, J.C.; Staels, B. Characterization of the human PPARalpha promoter: Identification of a functional nuclear receptor response element. Mol. Endocrinol. 2002, 16, 1013–1028. [Google Scholar] [PubMed]
- Luthe, G.; Jacobus, J.A.; Robertson, L.W. Receptor interactions by polybrominated diphenyl ethers versus polychlorinated biphenyls: A theoretical Structure-activity assessment. Environ. Toxicol. Pharmacol. 2008, 25, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Tung, E.W.; Boudreau, A.; Wade, M.G.; Atlas, E. Induction of adipocyte differentiation by polybrominated diphenyl ethers (PBDEs) in 3T3-L1 cells. PLoS ONE 2014, 9, e94583. [Google Scholar] [CrossRef] [PubMed]
- Kamstra, J.H.; Hruba, E.; Blumberg, B.; Janesick, A.; Mandrup, S.; Hamers, T.; Legler, J. Transcriptional and epigenetic mechanisms underlying enhanced in vitro adipocyte differentiation by the brominated flame retardant BDE-47. Environ. Sci. Technol. 2014, 48, 4110–4119. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Webster, T.F.; Ferguson, P.L.; Stapleton, H.M. Characterizing the peroxisome proliferator-activated receptor (PPARgamma) ligand binding potential of several major flame retardants, their metabolites, and chemical mixtures in house dust. Environ. Health Perspect. 2015, 123, 166–172. [Google Scholar] [PubMed]
- Grun, F.; Blumberg, B. Endocrine disrupters as obesogens. Mol. Cell. Endocrinol. 2009, 304, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Laplante, M.; Sabatini, D.M. Regulation of mTORC1 and its impact on gene expression at a glance. J. Cell Sci. 2013, 126, 1713–1719. [Google Scholar] [CrossRef] [PubMed]
- Laplante, M.; Sabatini, D.M. mTOR signaling in growth control and disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, S.; Peterson, T.R.; Laplante, M.; Oh, S.; Sabatini, D.M. mTORC1 controls fasting-induced ketogenesis and its modulation by ageing. Nature 2010, 468, 1100–1104. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | qPCR Primers | Fold Change | |
|---|---|---|---|
| RNA-seq | qPCR | ||
| Grem2 | GAGGAGAGGGACAGGGAGAC | 2.8 | 2.6 |
| AGCGAGAGCTTCCAGAACAT | |||
| Cish | CGTTGTCTCTGGGACATGGT | 4.4 | 3.9 |
| ACAGCCAGCAAAGGACAAGA | |||
| Sfrp2 | AGGACAACGACCTCTGCATC | 4.3 | 4.2 |
| TTCTTGGTTTTGCAGGCTTC | |||
| Fgg | AACCCACGAGACAAGCATTC | −15.7 | −16.3 |
| CAAGCTGGGCTACCTTCTGT | |||
| F2 | CCCTGGTTAGCCTGGTACAC | −16.2 | −13.9 |
| CAGCGATAGTGCTTGCTGAG | |||
| Apob | AAAACTTAAGCTTCAGCTGTCCA | −20.3 | −17.5 |
| CTTTCGGAGGTGCTTGAATC | |||
| B2m | CCGGCCTGTATGCTATCCAG | -- | housekeeping |
| TGTTCGGCTTCCCATTCTCC | |||
| Functional Annotation Cluster | Enrichment Score | Genes | p-Value | Benjamini |
|---|---|---|---|---|
| Secreted proteins | 11.17 | 38 | 6.7 × 10−6–3.0 × 10−18 | 7.0 × 10−4–5.9 × 10−16 |
| Peptidase inhibitor activity | 5.15 | 10 | 2.7 × 10−2–1.9 × 10−9 | 6.2 × 10−1–9.2 × 10−8 |
| Blood coagulation | 3.14 | 22 | 2.7 × 10−2–1.9 × 10−9 | 6.2 × 10−1–9.2 × 10−8 |
| Biosynthesis of amino acids | 2.81 | 6 | 4.6 × 10−2–1.2 × 10−4 | 3.0 × 10−1–5.7 × 10−3 |
| Microsomal metabolism | 2.40 | 7 | 1.1 × 10−1–1.9 × 10−5 | 5.3 × 10−1–4.5 × 10−4 |
| Lipid metabolism | 2.27 | 5 | 3.8 × 10−2–7.8 × 10−4 | 3.9 × 10−1–3.0 × 10−2 |
| Canonical Pathway | p-Value | Overlap | Count |
|---|---|---|---|
| FXR/RXR Activation | 2.4 × 10−23 | 25.4% | 32/126 |
| LXR/RXR Activation | 2.99 × 10−17 | 21.5% | 26/121 |
| Acute Phase Response Signaling | 3.02 × 10−17 | 17.8% | 30/169 |
| Coagulation System | 1.71 × 10−15 | 42.9% | 15/35 |
| Extrinsic Prothrombin Activation Pathway | 3.68 × 10−11 | 56.2% | 9/16 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abrha, A.; Suvorov, A. Transcriptomic Analysis of Gonadal Adipose Tissue in Male Mice Exposed Perinatally to 2,2′,4,4′-Tetrabromodiphenyl Ether (BDE-47). Toxics 2018, 6, 21. https://doi.org/10.3390/toxics6020021
Abrha A, Suvorov A. Transcriptomic Analysis of Gonadal Adipose Tissue in Male Mice Exposed Perinatally to 2,2′,4,4′-Tetrabromodiphenyl Ether (BDE-47). Toxics. 2018; 6(2):21. https://doi.org/10.3390/toxics6020021
Chicago/Turabian StyleAbrha, Aser, and Alexander Suvorov. 2018. "Transcriptomic Analysis of Gonadal Adipose Tissue in Male Mice Exposed Perinatally to 2,2′,4,4′-Tetrabromodiphenyl Ether (BDE-47)" Toxics 6, no. 2: 21. https://doi.org/10.3390/toxics6020021
APA StyleAbrha, A., & Suvorov, A. (2018). Transcriptomic Analysis of Gonadal Adipose Tissue in Male Mice Exposed Perinatally to 2,2′,4,4′-Tetrabromodiphenyl Ether (BDE-47). Toxics, 6(2), 21. https://doi.org/10.3390/toxics6020021